

Summary
Vi beskriver en metod som kombinerar automatiserad cellodling med högt innehåll imaging att visualisera och kvantifiera olika cellulära processer och strukturer, i en hög genomströmning sätt. Sådana metoder kan stöd i den fortsatta funktionella notering av genomet samt identifiera nätverk sjukdomsgen och potentiella mål drog.
Abstract
Den funktionella annotering av genom, byggande av molekylära nätverk och nya läkemedelsmål identifiering, är viktiga utmaningar som måste lösas som ett mycket angeläget 1-4. Flera kompletterande "omik" metoder har gett ledtrådar till de genetiska riskfaktorerna och patogena mekanismer som ligger bakom många neurodegenerativa sjukdomar, men de flesta fynd fortfarande kräver fungerande validering 5. Till exempel är en nyligen genomet hela föreningen studier för Parkinsons sjukdom (PD), identifierat många nya loci som riskfaktorer för sjukdomen, men den underliggande orsakande variant (er) eller patogenetisk mekanism inte känt 6, 7. Eftersom varje tillhörande region kan innehålla flera gener, skulle den funktionella bedömningen av varje av de gener på fenotyper i samband med sjukdomen, med traditionell teknik cellbiologi tar för lång tid.
Det finns också ett behov av att förstå de molekylära nätverk som sammanbindergenetiska mutationer till fenotyper de orsakar. Det förväntas att sjukdomen fenotyper är resultatet av flera interaktioner som har stört. Rekonstruktion av dessa nätverk med traditionella molekylära metoder skulle vara tidskrävande. Dessutom kommer nätverket förutsägelser från oberoende studier av enskilda komponenter, reduktionism metod underskattar förmodligen nätverket komplexitet 8. Denna underskattning kan delvis förklara den låga andelen framgångsrika godkännande av läkemedel på grund av oönskade eller toxiska biverkningar. Att få ett nätverk perspektiv av sjukdomsrelaterade vägar med HT / HC cellulära screening metoder, och identifierar de viktigaste noder inom dessa vägar, skulle kunna leda till identifiering av mål som är mer lämpade för terapeutisk intervention.
High-throughput screening (HTS) är en idealisk metod för att hantera dessa frågor 9-12. men traditionella metoder endimensionella hel-och cell-analyser, som används förenklarSTIC avläsningar för komplexa biologiska processer. De var oförmögna att samtidigt kvantifiera många fenotyper observerats i neurodegenerativa sjukdomar som axonal transport underskott eller förändringar i morfologi fastigheterna 13, 14. Denna metod kunde inte användas för att undersöka dynamiken i cellulära processer eller händelser patogena som inträffar i en delmängd av celler. För att kvantifiera sådana funktioner man måste flytta till flerdimensionella fenotyper kallas hög halt screening (HCS) 4, 15-17. HCS är cellbaserade kvantifiering av flera processer samtidigt, vilket ger en mer detaljerad bild av cellulära svar på olika störningar jämfört med HTS.
HCS har många fördelar jämfört HTS 18, 19, men bedriver en hög genomströmning (HT)-högt innehåll (HC) skärmen i neuronala modeller är problematisk på grund av höga kostnader, miljö variation och mänskliga fel. För att upptäcka cellulära svaren på en "fenomik" skalamed hjälp av HC avbildning man har för att minska variation och fel, samtidigt som man ökar känslighet och reproducerbarhet.
Häri beskriver vi en metod för att korrekt och tillförlitligt sätt utföra shRNA skärmar med hjälp av automatiserade cellodling 20 och HC avbildning i neuronala cellulära modeller. Vi beskriver hur vi har använt denna metod för att identifiera modulatorer för ett visst protein, DJ1, som när muterade orsakar autosomalt recessiv parkinsonism 21.
Kombinera den mångsidighet av HC avbildning med HT metoder är det möjligt att exakt kvantifiera en uppsjö av fenotyper. Detta kan sedan utnyttjas för att öka vår förståelse av genomet, de som är involverade i sjukdomens patogenes samt identifiera potentiella terapeutiska mål.
Protocol
1. Automatiserad anhängare cellodling (Figur 1) 20
- Förbered det automatiska systemet cellodling för inmatning av plattor cellkultur. Ladda förbrukningsvaror (t ex pipettspetsar, cellodling tallrikar, analys plattor) i systemet via det grafiska användargränssnittet (GUI). Se till att det finns tillräckligt med cellodling medier, buffrad fosfat saltlösning (PBS) och trypsin i robotsystem.
- Manuellt frö två omnitray plattor med 2 x 10 6 celler per platta, av SH-SY5Y neuroblastom cellinje. Behåll celler i Opti-MEM med 10% fetalt bovint serum (FBS). Sätt plattorna i cellen kulturen roboten med hjälp av det grafiska användargränssnittet. Cellerna kommer att inkuberas vid 37 ° C och 5% CO 2.
- Välj vilken cellodling protokoll måste påbörjas 20. Man kan välja mellan en anhängare cellodling 22, odling och expansion av embryonala stamceller (ES-celler) på celler musen feeder 23 eller odling av fjädring CELLS 22.
- Markera den tillhörande protokoll cellkultur (figur 1) och se tillhörande cellinje specifik parameter filer justeras så att sammanflödet tröskeln (det område i omnitray platta som innehåller celler) är satt till 70%. Ställ totala trypsinization tid till två minuter.
- Instruera roboten för att förbereda nya omnitray tallrikar, med en sådd cell antal 2 x 10 6 celler per platta.
- Ingång omnitray plattor i cellodling systemet med hjälp av automatiserade tillhörande protokoll cellkultur (Figur 1). Detta protokoll omfattar följande steg: Plattorna inkuberas och avbildat tills de fördefinierade sammanflödet tröskel. Om cellerna inte når sammanflödet tröskeln inom 5 avläsningar, är plattorna tas bort från systemet. Vid ankomsten till den användardefinierade sammanflödet nivå räknas celler tvättas, trypsinized och räknas. Ett fördefinierat antal celler läggs till nya omnitray plattor och om det finns tillräckligt många celler, en viss steler av analysen plattor transporteras till däck och ett definierat antal celler doseras i varje brunn. Analysen plattor direkt kan avbildas med den integrerade mikroskop eller utdata från systemet för vidare behandling.
- Instruera systemet för att förbereda 4 analysen plattor per omnitray tallrik, med totalt 5000 celler per brunn.
2. shRNA virus produktion och plätering i analysen plattor (Tidsåtgång: 6 dagar)
- Odla bakterie glycerol lager som innehåller shRNA vektorer (Open Biosystems, TRC1) övernattning i 2 ml Luria-Bertani medelstora media som innehåller 100 mikrogram / ml ampicilin (Sigma-Aldrich).
- Utdrag plasmider enligt tillverkarens protokollet (Promega guiden MagneSil TFX).
- Producera virus använder RNAi Consortium hög genomströmning Lentiviral Produktion (96 brunnar) protokoll 24. Att arbeta med lentivirus är relativt säker eftersom viruset partiklar används för transduktion är replikationskompetenta bristfällig och split-gen förpackningar strategier används för sin produktion. Men när man arbetar med lentivirus, ytterligare biosäkerhet rutiner är nödvändiga för att minimera risken för sig själv och andra 25. Alla experiment måste utföras i ett MLII eller BSL2 säkerhetsnivå laboratorium. Alla plaster (pipetter, plastskålar, media) som har varit i kontakt med lentivirus partiklar skall inkuberas med blekmedel i 24 timmar före kassering.
- Beräkna den mångfald av infektion (MOI) av lentivirus genom att bestämma den andel av GFP positiva celler med pLKO.1 GFP plasmiden (Sigma-Aldrich).
- Tavla lentivirus i analysen tallrikar, med en MOI på 3.
3. Lentiviral transduktion och neuronal differentiering av SH-SY5Y celler (Tidsåtgång: 6 dagar)
- Celler läggs till analysen plåtarna (se steg 1,7). Ladda analys plattor innehåller shRNA lentivirus i det automatiska cellodlingssystem.
- Efter 24 timmar, media påanalysen plattor kommer att ändras till Opti-MEM innehållande 0,5% FBS och 0,1 mikroM retinoinsyra att börja differentiering processen. Differentiering av SH-SY5Y celler gör visualisering av neuritic strukturer och synkroniserar celldelning.
- Fortsätt inkubering av analysen plattorna i differentieringen media för 5 dagar. Detta säkerställer maximal ÖVERVÄLDIGANDE av mål genuttryck.
- På dag 5, tillsätt 50 mikroM H 2 O 2 till hälften av analysen plattor i 24 timmar för att stimulera flyttning av DJ1 till mitokondrierna.
- På dag 6, lägg Mitotracker CmxROS (Invitrogen) till cellerna på en slutlig koncentration av 200 Nm per brunn och inkubera vid 37 ° C i 30 minuter.
- Man kan låta systemet bildplattorna direkt med hjälp av HC kameran eller plattor kan exporteras från systemet för vidare behandling.
4. Automatiserad immunfärgning av analysen plattor (Tidsåtgång: 2 dagar)
Bildkvaliteten är paramount för att genomföra en känslig och pålitlig HCS. Skador på cellulär cellslager på grund av felaktiga pipettering kan leda till dålig bildkvalitet och oåterkallegia resultat. För att minimera skadorna cellager var immunfärgning genomförs med hjälp av en fjärrstyrd station. Proceduren liknar en som har beskrivits tidigare 26, men har anpassats för att öka genomströmningen och minska förbrukningsartiklar användning.
- Fix celler med 100 l 4% paraformaldehyd förvärmas till 37 ° C. Inkubera 20 minuter vid rumstemperatur.
- Tvätta cellerna med 200 l PBS i 5 minuter, 3 gånger.
- Inkubera assay plattor med 200 l PBS innehållande 0,1% Triton (PBST) i 10 minuter.
- Tvätta cellerna med 200 l PBS i 5 minuter, 3 gånger.
- Inkubera assay plattor med 200 l av block buffert (PBST med 5% FBS) i 1 timme i rumstemperatur.
- Tvätta cellerna med 200 l PBS i 5 minuter, 3 gånger.
- Inkubera med follpå grund av primära antikroppar över natten vid 4 ° C:
- Goat DJ1 N20 (Santa Cruz, 5 mikrogram / ml)
- Kanin β-III tubulin (Sigma-Aldrich, 1 mikrogram / ml)
- Följande dag, tvätta cellerna med 200 l PBS i 5 minuter, 3 gånger.
- Inkubera assay plattor med följande sekundära antikroppar 1 timme i rumstemperatur:
- AlexaFluor 488 åsna anti-get (Invitrogen, 2 mikrogram / ml)
- AlexaFluor 647 get-anti-kanin (Invitrogen, 2 mikrogram / ml
- Tvätta cellerna med 200 l PBS i 5 minuter, 3 gånger.
- Inkubera cellerna med Hoechst (Invitrogen, 1 mikrogram / ml) i 10 minuter.
- Tvätta cellerna med 200 l PBS i 5 minuter, 3 gånger.
- Förvara plattorna vid 4 ° C tills de kan avbildas.
5. Hög halt bild förvärv och bildanalys (Tidsåtgång: 5 dagar)
- Bild totalt 30 fält per brunn med 20x objektiv. Visualisera DJ1 med FITC filter set, mitokondrierna med TRITC filtret set, β-III tubulin med Cy5 filtret inställt och kärnorna med hjälp av UV-filter set (Figur 3).
- Analysera bilder med kompartmentanalys Bioapplication (Cellomics, ThermoFisher) för att fastställa den genomsnittliga intensiteten i Mitotracker signalen i mitokondrierna. (Figur 4B, F).
- För att bestämma den genomsnittliga överlappningen koefficienten mellan DJ1 och mitokondrier, analysera bilderna med hjälp av Cellomics Colocalisation bioapplication (Cellomics, ThermoFisher). Definiera områden av intresse (ROI) enligt följande: ROI A - kärnan (Figur 4A, E), ROI B - mitokondrier (Figur 4 B, F). Uteslut ROI A från ROI B för att säkerställa en analys av enbart cytoplasman. Definiera mitokondrierna som målområde jag och DJ1 som målområde II (Figur 4C, G).
- Analysera bilder med den neuronala Profiling bioapplication (Cellomics, Thermofisher) för att spåra den genomsnittliga längden av neurites från β-III-tubulin färgning (Figur4D, H).
- Bildplattor med Opera LX automatiserade konfokala läsare (Perkin-Elmer). Bild totalt 30 fält per brunn med 60x objektiv med nedsänkning i vatten. Visualisera mitokondrier med 561 nM laser och kärnor med UV-excitation.
- Analysera bilder med Spot-Edge-Ridge (SER) textur funktioner algoritm. Den SER-Ridge filtret sänder intensiteten i pixlar bildar ås-liknande mönster. Ju mer fragmenterad mitokondrierna, desto högre SER-Ridge poäng (Figur 8).
6. Data normalisering och analys
- Importera data från bildanalys programvara i BioConductor CellHTS2 paket för R-programvaran miljön (R-version 2.11.1, BioConductor version 2.6).
- Logaritmen bas (2) omvandla informationen innan per platta median baserat normalisering 27, 28. Applicera inte variansen justeringen per platta.
- Att identifiera modifierare av en fenotyp, använd en tvåvägs ANOVA mellan different behandlingsgrupperna dvs Scrambled infekterade obehandlade celler kontra kodade toxin behandlade cellerna kontra målgenen obehandlade celler kontra målgenen behandlade cellerna (figur 5-7).
7. Representativa resultat
Mutationer inom DJ1 ge upphov till tidig debut-recessiv parkinsonism 21, men det är oklart hur förlusten av DJ1 ger upphov till sjukdomen fenotyp. Det är känt att cellerna brister i DJ1 är mer mottagliga för oxidativ stress-inducerad celldöd och som svar på oxidativ stress, DJ1 translocates från cytoplasman till mitokondrierna 29, 30. Genom att bygga HC-analyser för att övervaka dessa fenotyper, kan vi identifiera gener som reglerar eller påverkar fenotyper i samband med DJ1. Denna metod kan hjälpa tyda de vägar där DJ1 funktioner och som kan vara inblandade i sjukdomen patogenes.
Exempel på en epistatisk interaktion med DJ1 (Figur 5): Knockdown av DJ1 i utsatta cellertill toxisk resulterar i en ökad förlust av cellernas livskraft (Bar-B: image-B) jämfört med celler infekterade med äggröra lentivirus (BAR-A: image-A). ÖVERVÄLDIGANDE av målgenen A har en liknande effekt som observerats i celler med en DJ1 ÖVERVÄLDIGANDE (BAR-C: image-C). ÖVERVÄLDIGANDE både DJ1 och målgenen En resulterar i en betydligt större förlust av cellernas livskraft än förlust av antingen gen ensam (BAR-D: image-D). Detta tyder på en epistatisk interaktion mellan DJ1 och målgenen A.
Exempel på en gen som reglerar DJ1 translokation (Figur 6): När celler utsätts för ett toxin, DJ1 translocates från cytoplasman till mitokondrierna, som kvantifieras av en högre överlappning koefficient mellan DJ1 och mitokondrier (BAR-A: image kontra BAR-C: bild C). I celler där målgenen B har tystats, mindre DJ1 translocates till mitokondrierna när cellerna utsätts för giftet. Detta tyder på att målgenen B är inblandad i transporten av DJ1 till mitokondrierna. (Bar-B: bild B och Bar-D: bild D)
Exempel på en gen som deltar i neuronal utväxt (Figur 7): Knockdown av målgenen C i vild typ SH-SY5Y celler resulterar i en betydande ökning av neurite längd (Bar-B: image-B) jämfört med celler infekterade med lentivirus uttrycka äggröra shRNA (BAR-A: bild A). Denna effekt är vilse i cellerna inkuberas med toxin (BAR-C och D).
Exempel på en gen som deltar i mitokondriella morfologi (figur 8): Infektion av vilda typen SH-SY5Y celler med shRNA riktade genen D resulterar i en minskning av den mitokondriella SER-Ridge segmentering värde (Figur 8, Bild C och D) jämfört med celler infekterade med äggröra lentivirus (Figur 8, Bild A och B).
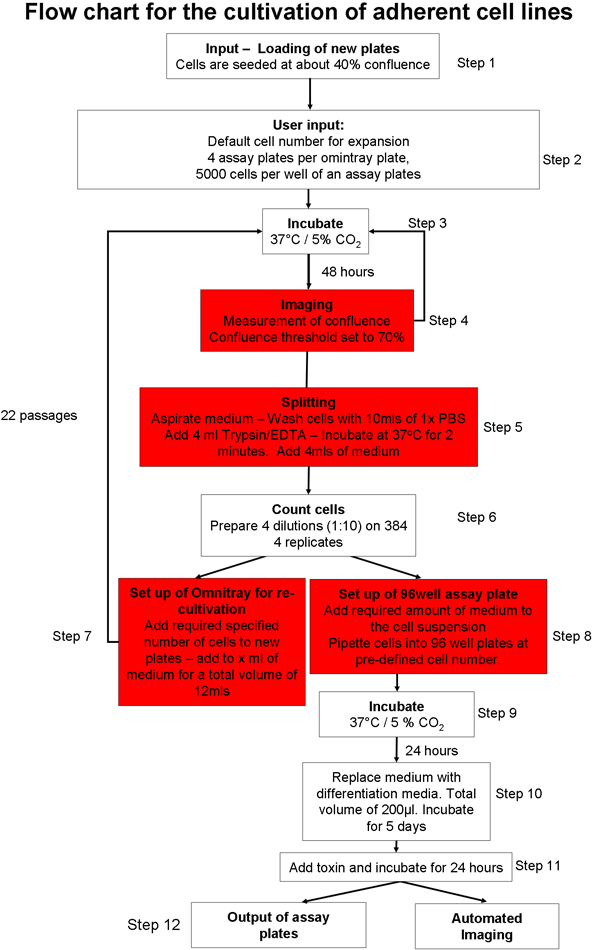
Figur 1 Sammanfattning av automatiserade cellodling protokollet.:

Figur 2 Schematisk översikt av urvalsförfarandet, bildanalys och statistiska metoder som används vid urvalsförfarandet:. I) Celler odlas tills de är sammanhängande och därefter klädd i analysen plattorna innehåller shRNA lentivirus. Celler är differentierade i 5 dagar och toxin läggs sedan till plattorna under 24 timmar. Assay plattorna är utdata från systemet och immunostained. Den tid det tar för var och en av de processer som anges inom parentes. ii) Data förvärvas med hjälp av en HC imager (cellviabiliteten, protein translokation och neuronal utväxt) och ett automatiserat konfokala imager (mitokondriell morfologi). Data exporteras till CellHTS2 paketet inom R, (2) bas logga omvandlas och normaliseras. Två-vägs ANOVA används för att identifiera signifikanta interaktioner mellan olika variabler.
Figur 3. Komposit bilder av celler som förvärvats av HC avbildning. A) Obehandlad celler, B) celler som behandlats med H 2 O 2. DJ1 är märkt i grönt, mitokondrierna i rött och kärnan i blått. Neurite färgning är inte markerad.

Figur 4. Kvantifiering av flera cellulära funktioner från en HCS. AD) Obehandlad SH-SY5Y celler. EH) H 2 O 2 behandlas SH-SY5Y celler. A, E) Nuclei segmentering och definition av ROI A, B, F) Identifiering och kvantifiering av mitokondrier, ROI B, C, G) Fastställande av DJ1, Target kanal II, D, G) Identifikation och beräkning av genomsnittliga neurite längd. Celler nära kanten av bilden är undantagna från analysen. Inset är bilder före analys.

Figur 6. Identifiering av en gen som reglerar DJ1 translokation (bokstäver på staplarna motsvarar bokstäverna på bilderna). Bilder A till D är SH-SY5Y celler märkta för DJ1 (grön), mitokondrier (röd) och kärnor (blå) som användes för kvantifiering av DJ1 translokation till mitokondrierna.

Figur 7. Identifiering av en gen som deltar i neuronal utväxt (bokstäver på staplarna motsvarar bokstäverna på bilderna). Bilder A till D är SH-SY5Y celler märkta för β-III-tubulin (grön) och kärnor (blå) som användes för kvantifiering av neurite längd.

Figur 8. Identifiering av en gen som deltar i reglering av mitokondriell morfologi. Bild A och C är sammansatta bilder av SH-SY5Y celler infekterade med äggröra shRNA eller en shRNA inriktning gen D respektive. Mitokondrierna är färgade i rött, medan kärnor är färgade i blått. Bild B och D är visualiseringar av SER-Ridge kvantifiering.
Discussion
Med minskade kostnader för HT / HC cellulära screening system, i kombination med tillgången på kraftfulla Genomvid verktyg för att modifiera genfunktion, är HT / HC skärmar blir allt vanligare i den akademiska världen. Den metoden har redan framgångsrikt tillämpats på olika forskningsområden som identifiering av läkemedel mål i cancer 9, 31-33 och embryonal utveckling 34-36 och har även potential för tillämpning i dechiffrera involverade i de neuropsykiatriska störningar 37,38. Men genomförandet av ett sådant system kräver en betydande investering i tid och ansträngning med processoptimering ofta tar minst 6 månader. Alla åtgärder, såsom trypsinization tider, pipettering hastigheter och densitet seeda måste justeras för att säkerställa att cellerna är friska och växer ständigt. Förebyggande av bakteriell kontamination är en av de svåraste utmaningarna för automatiserad cellodling med veckostädning protokoll i Combination med konstant sköljning av all vätska som bär linjer med 70% etanol är nödvändigt för föroreningar fria kulturer. Det blir också nödvändigt att förbättra robotik, så att ytterligare instrument, t.ex. konfokala mikroskop för högre upplösning och -80 ° C frysar för sammansatta lagring kan integreras.
Det finns också begränsningar som måste åtgärdas för att förbättra den känsliga, hastighet och nyttan av denna metod för att studera gen nätverk och identifiera gener involverade i patogena molekylära vägar.
Att genomföra en HT / HC skärmen och se till att tillförlitliga uppgifter samlas in, flera aspekter måste optimeras. För det första är tillförlitligheten i mätningen av största vikt och är beroende av robusthet och känslighet analysen. Till exempel analyser som beskrivs ovan är lämpliga för mindre skärmar, men är svåra att genomföra på ett genomet stor skala, på grund av de steg antalet bearbetning som krävs innan bilden förvärvet.Således skulle man ha för att bygga stabila cellinjer som uttrycker reportern genen, vilket skulle möjliggöra direkt avbildning och leda till minskad variation på grund av minskat antal processteg. För närvarande utforma ett test som korrekt beskriver och tillförlitligt kvantifierar en fenotyp av intresse är en stor flaskhals i HC screening process.
Många skärmar bedrivs i däggdjursceller med olika RNAi bibliotek, som alla lider ej åsyftade effekter, begränsad geners uttryck effektivitet och ofullständig genomet täckning. Således biblioteken behöver göras som är mer specifika, potent och har bättre täckning. Ansträngningar görs för att skapa sådana Förhoppningen är sådana strävanden kommer att förbättra reproducerbarheten för HT / HC SCRE sv träffar.
En begränsning av många stora baserat skala cell skärmar är att de bedrivs i neuroblastom celler eftersom de kan vara genmanipulerade och odlade ett stort antal relativt enkelt. Dock är betydelsen av "träffar" som identifierats i ex vivo-modeller cellodling för in vivo-funktionen tveksamt, särskilt som hjärnan består av högt specialiserade celltyper som bildar en tät och komplicerat nätverk av synapsförbindelser att fungera som en starkt integrerad enhet. Som en följd är det vanligt att träffar identifieras med hjälp av screening som beskrivs ovan, är validerade i sekundära skärmar med hjälp av ytterligare tekniker och i mer fysiologiskt relevanta modeller 39. För att förbättra översättningen av träffar som identifierats under HCS, mer representativ och sofistikerade modeller, såsom primär celler och differentierade stamceller eller co-kultur-system behöver utvecklas och anpassas för HT / HC metoder.
ntent "> Med en kombination av automatiserade cellodling och HC avbildning ett snabbt kan få nya insikter i hur nervceller fungerar och bestämma vilka vägar är viktiga för sjukdomens utveckling. kombinera HCS / HTS data med information som härrör från andra" omik synsätt, kommer det då vara möjligt att konstruera ett system biologi överblick av sjukdomar i hjärnan, vilket underlättar terapeutisk utveckling.Disclosures
Vi har inget att lämna ut
Acknowledgments
Vi tackar Hamilton programmerare och specialister för fortsatt stöd och Eva Blaas för tekniskt stöd. Detta arbete stöddes av två NWO Investeringsbidrag (911-07-031 och 40-00506-98-10011), The Prinses Beatrix Fonds Wetenschapsprijs 2009 och neurovetenskap Campus Amsterdam, SJ stöds av Ti-Pharma: T5-207.
Materials
| Name | Company | Catalog Number | Comments |
| AI.CELLHOST | Hamilton Co | http://www.hamiltonrobotics.com/en-uk/applications/cellomics/ | |
| OPTI-MEM | Invitrogen | 31985-054 | |
| RETINOIC ACID | Sigma-Aldrich | R2625 | |
| OMNITRAY PLATES | Nalge Nunc international | 465219 | |
| 96 WELL CULTURE PLATES | Greiner Bio-One | 655086 | |
| DJ1 N20 ANTIBODY | Santa Cruz Biotechnology, Inc. | SC27004 | |
| BETA-III TUBULIN ANTIBODY | Sigma-Aldrich | T3952 | |
| MITOTRACKER CMXROS | Invitrogen | M-7512 | |
| H–CHST-33342 | Invitrogen | H1399 | |
| HYDROGEN PEROXIDE | Sigma-Aldrich | 216763-100ML | |
| TRYPSIN | Invitrogen | 25050014 | |
| DULBECCO’S PHOSPHATE BUFFERED SALINE | Invitrogen | 14190086 | |
| PROMEGA WIZARD MAGNESIL TFX | Promega Corp. | A2380 | |
| SHRNA CLONES | Open Biosystems | http://www.openbiosystems.com/RNAi/shRNALibraries/ TRCLibraryDetails/ | |
| CELLOMICS BIOAPPLICATIONS | Thermo Fisher Scientific, Inc. | http://www.thermo.com/hcs |
References
- Geschwind, D. H., Konopka, G. Neuroscience in the era of functional genomics and systems biology. Nature. 461, 908-915 (2009).
- Ge, H., Walhout, A. J., Vidal, M. Integrating 'omic' information: a bridge between genomics and systems biology. Trends. Genet. 19, 551-560 (2003).
- Zhu, H., Snyder, M. 'Omic' approaches for unraveling signaling networks. Curr. Opin. Cell. Biol. 14, 173-179 (2002).
- Jain, S., Heutink, P. From single genes to gene networks: high-throughput-high-content screening for neurological disease. Neuron. 68, 207-217 (2010).
- Manolio, T. A. Finding the missing heritability of complex diseases. Nature. 461, 747-753 (2009).
- Nalls, M. A. Imputation of sequence variants for identification of genetic risks for Parkinson's disease: a meta-analysis of genome-wide association studies. Lancet. 377, 641-649 (2011).
- Simon-Sanchez, J. Genome-wide association study confirms extant PD risk loci among the Dutch. Eur. J. Hum. Genet. 19 (6), 655-661 (2011).
- Van Regenmortel, M. H. Reductionism and complexity in molecular biology. Scientists now have the tools to unravel biological and overcome the limitations of reductionism. EMBO. Rep. 5, 1016-1020 (2004).
- Aherne, G. W., McDonald, E., Workman, P. Finding the needle in the haystack: why high-throughput screening is good for your health. Breast. Cancer. Res. 4, 148-154 (2002).
- An, W. F., Tolliday, N. J. Introduction: cell-based assays for high-throughput screening. Methods. Mol. Biol. 486, 1-12 (2009).
- Mayr, L. M., Bojanic, D. Novel trends in high-throughput screening. Curr. Opin. Pharmacol. 9, 580-588 (2009).
- Hertzberg, R. P., Pope, A. J. High-throughput screening: new technology for the 21st century. Curr. Opin. Chem. Biol. 4, 445-451 (2000).
- Conrad, C., Gerlich, D. W. Automated microscopy for high-content RNAi screening. J. Cell. Biol. 188, 453-461 (2010).
- Thomas, N. High-content screening: a decade of evolution. J. Biomol. Screen. 15, 1-9 (2010).
- Arrasate, M., Finkbeiner, S. Automated microscope system for determining factors that predict neuronal fate. Proc. Natl. Acad. Sci. U. S. A. 102, 3840-3845 (2005).
- Dragunow, M. High-content analysis in neuroscience. Nat. Rev. Neurosci. 9, 779-788 (2008).
- Varma, H., Lo, D. C., Stockwell, B. R. High throughput screening for neurodegeneration and complex disease phenotypes. Comb. Chem. High. Throughput. Screen. 11, 238-248 (2008).
- Durr, O. Robust hit identification by quality assurance and multivariate data analysis of a high-content, cell-based assay. J. Biomol. Screen. 12, 1042-1049 (2007).
- Miller, J. Quantitative relationships between huntingtin levels, polyglutamine length, inclusion body formation, and neuronal death provide novel insight into huntington's disease molecular pathogenesis. J. Neurosci. 30, 10541-10550 (2010).
- Jain, S., Sondervan, D., Rizzu, P., Bochdanovits, Z., Caminada, D., Heutink, P. The Complete Automation of Cell Culture. Journal of Biomolecular Screening. 16 (8), (2011).
- Bonifati, V. Mutations in the DJ-1 gene associated with autosomal recessive early-onset parkinsonism. Science. 299, 256-259 (2003).
- Ricardo, R., Phelan, K. Trypsinizing and Subculturing Mammalian Cells. J. Vis. Exp. (16), e755-e755 (2008).
- Kent, L. Culture and Maintenance of Human Embryonic Stem Cells. J. Vis. Exp. (34), e1427-e1427 (2009).
- Moffat, J. A lentiviral RNAi library for human and mouse genes applied to an arrayed viral high-content screen. Cell. 124, 1283-1298 (2006).
- Volksgezondheid, M. V. Integrale versie van de Regeling genetisch gemodificeerde organismen en het Besluit genetische gemodificeerde. Organismen. , (2004).
- Anderl, J. L., Redpath, S., Ball, A. J. A Neuronal and Astrocyte Co-Culture Assay for High Content Analysis of Neurotoxicity. J. Vis. Exp. (27), e1173-e1173 (2009).
- Wiles, A. M., Ravi, D., Bhavani, S., Bishop, A. J. An analysis of normalization methods for Drosophila RNAi genomic screens and development of a robust validation scheme. J. Biomol. Screen. 13, 777-784 (2008).
- Canet-Aviles, R. M. The Parkinson's disease protein DJ-1 is neuroprotective due to cysteine-sulfinic acid-driven mitochondrial localization. Proc. Natl. Acad. Sci. U. S. A. 101, 9103-9108 (2004).
- Blackinton, J. Formation of a stabilized cysteine sulfinic acid is critical for the mitochondrial function of the parkinsonism protein DJ-1. J. Biol. Chem. 284, 6476-6485 (2009).
- Gupta, P. B. Identification of selective inhibitors of cancer stem cells by high-throughput screening. Cell. 138, 645-659 (2009).
- Luo, J. A genome-wide RNAi screen identifies multiple synthetic lethal interactions with the Ras oncogene. Cell. 137, 835-848 (2009).
- Mouchet, E. H., Simpson, P. B. High-content assays in oncology drug discovery: opportunities and challenges. IDrugs. 11, 422-427 (2008).
- Vogt, A. Automated image-based phenotypic analysis in zebrafish embryos. Dev. Dyn. 238, 656-663 (2009).
- Pardo-Martin, C. High-throughput in vivo vertebrate screening. Nat. Methods. 7, 634-636 (2010).
- Vogt, A., Codore, H., Day, B. W., Hukriede, N. A., Tsang, M. Development of automated imaging and analysis for zebrafish chemical screens. J. Vis. Exp. (40), e1900-e1900 (2010).
- Ross, P. J., Ellis, J. Modeling complex neuropsychiatric disease with induced pluripotent stem cells. F1000. Biol. Rep. 2, 84-84 (2010).
- Ebert, A. D., Svendsen, C. N. Human stem cells and drug screening: opportunities and challenges. Nat. Rev. Drug. Discov. 9, 367-372 (2010).
- An, W. F., Tolliday, N. Cell-based assays for high-throughput screening. Mol. Biotechnol. 45, 180-186 (2010).
