

Summary
Un rapide
Abstract
Criblage de composés pour l'activité in vivo peut être utilisé en tant que première étape d'identifier des candidats qui peuvent être développés en 1,2 agents pharmacologiques. Nous avons développé un roman nanoinjection / électrophysiologie dosage qui permet la détection des effets modulateurs de composés bioactifs sur la fonction d'un circuit neuronal qui sert de médiateur de la réaction de fuite chez Drosophila melanogaster 3,4. Notre test in vivo, qui utilise le système de Drosophila fibre géante (GFS, Figure 1) permet le dépistage des différents types de composés, tels que de petites molécules ou des peptides, et exige que des quantités minimales à obtenir un effet. En outre, le GFS Drosophila offre une grande variété de cibles moléculaires potentielles sur les neurones ou les muscles. Les fibres géantes (GFS) synapse électrique (Gap jonctions) ainsi que chimiquement (cholinergique) sur une synapse périphérique Interneuron (PSI) et le Tergo Muscle Trochanteral neurone (TTMn) 5 6. Enfin, la jonction neuromusculaire (JNM) de l'TTMn et le DLMn avec le saut (TTM) et les muscles de vol (DLM) sont glutamatergique 7-12. Ici, nous montrons comment injecter des quantités nanolitre d'un composé, tout en obtenant électrophysiologiques des enregistrements intracellulaires du système de fibre géante 13 et la façon de surveiller les effets du composé sur la fonction de ce circuit. Nous montrons la spécificité du test avec du citrate de methyllycaconitine (MLA), un antagoniste nAChR, qui perturbe le PSI à DLMn connexion, mais pas le GF TTMn connexion ou la fonction de la jonction neuromusculaire au saut ou les muscles de vol.
Avant de commencer cette vidéo, il est essentiel que vous regardez attentivement et se familiariser avec la vidéo intitulée "JoVE enregistrements électrophysiologiques de la voie des fibres Géant de D. melanogaster "de Augustin et al 7, comme la vidéo présentée ici est conçu comme une extension de cette technique existante. Ici, nous utilisons la méthode électrophysiologique des enregistrements et de se concentrer en détail que sur l'ajout des nanoinjections appariés et de la technique de surveillance.
Protocol
1. Électrophysiologie Rig Set-up
- L'équipement requis pour la plate-forme d'électrophysiologie set-up est décrite en détail par Augustin et al. dans le présent Journal 14. S'il vous plaît se référer à cet article pour une explication détaillée des appareils électrophysiologiques nécessaires.
- Modifier la plate-forme décrit précédemment électrophysiologie set-up 14 par l'ajout d'un micromanipulateur sixième, qui détient le nanoinjector. Pour un accès facile à la tête de l'animal, il doit être placé entre les deux micromanipulateurs électrode de stimulation comme le montre la figure 2.
- Avant de commencer l'expérience, veiller à ce que vous avez une gamme à l'aise sur tous les axes de rotation avec tous les micromanipulateurs, et que toutes les électrodes ainsi que la micropipette d'injection (voir étape 2) peut atteindre l'animal.
2. Nanoinjection Set-up
- Le nanoinjection set-up nécessite une Nanoliter2000 (World Precisisur les instruments, Sarasota, Floride, Etats-Unis) ou de type similaire de l'injecteur qui permet des injections contrôlées en quantités nanolitre.
- Préparer aiguille d'injection en utilisant les micropipettes de verre sont fournies avec l'injecteur en les tirant à une résistance de 80-100 MQ avec une électrode de traction.
- Pour les injections lisses, il est nécessaire de biseauter les micropipettes à une ouverture de 11 à 17 um à un angle de 45 degrés (figure 3).
- Lentement remblayer la micropipette d'injection avec de l'huile synthétique à l'aide d'une seringue Hamilton selon les instructions du manuel Nanoliter2000, veiller à ce que les bulles d'air sont présents.
- Fixez soigneusement la micropipette sur la nanoinjector et se préparent à charger le composé en vidant l'excédent d'huile selon les instructions du manuel Nanoliter2000.
- Placez l'injecteur sur le micromanipulateur et charger le composé selon les instructions dans le manuel Nanoliter2000. Assurez-vous que la pointe de la micropipette ne se brise pas au cours de cetteProcédure.
- Réglez la quantité désirée de nanolitres être injectés dans la boîte de commande de l'injecteur selon les instructions du manuel Nanoliter2000. S'il vous plaît noter que le montant total injecté ne doit pas dépasser 100 nl. Nous avons constaté que de plus grandes quantités de solutions de contrôle salines peuvent affecter la fonction du circuit de GFS.
- Il est essentiel d'avoir l'injecteur débranché de la boîte de commande lors de l'acquisition d'enregistrement à l'exception de l'injection lui-même, depuis l'alimentation électrique de l'nanoinjector interfère avec les enregistrements, qui est visible comme un bruit de fond (Figure 4). Cependant, ne pas déconnecter l'alimentation de la boîte de commande comme il le re-régler.
3. Drosophila melanogaster Préparation
- Anesthésier 2 à 6 mouches d'un jour avec CO 2 ou sur la glace comme décrit précédemment 14,15.
- Une fois immobile, utilisez une paire de pince à épiler pour transférer l'animal à un réseau Wi petite plaquee de la cire molle dentaire par le ramasser de ses jambes. S'il vous plaît noter que la mouche des fruits mâle pèse environ 1,0 mg et une mouche à fruit femelle pèse environ 1,2 mg, ce qui composé à poids corporel ratio est différent chez les mouches mâles et femelles. Par conséquent, il est recommandé d'utiliser un seul sexe pour les expériences.
- Comme décrit précédemment 14,15, soigneusement monter du côté battant dorsale et veiller à ce que le thorax et la tête sont immobilisées avec de la cire dentaire mou placé autour du corps. Étaler soigneusement les ailes de telle sorte qu'elles se trouvaient perpendiculaire au thorax (Figure 2, C). La mouche doit être montée avec le moins de dommages possible.
4. Nanoinjection Jumelé / électrophysiologie
- Placez la volée monté sur la plate-forme d'électrophysiologie avec sa tête vers les électrodes de stimulation.
- Empaler l'animal avec le stimulant, sol correspondant et des électrodes d'enregistrement tel que décrit précédemment 14,15 (Figure 2, C). Sauf indication contraire souhaité, électrode d'enregistrement seul endroit dans le DLM et l'autre dans le muscle TTM. Le muscle DLM est situé dans le thorax entre les antérieurs dorso-centrales poils et la ligne médiane de la mouche. Le muscle TTM est situé près des attaches d'aile, entre les postérieurs et antérieurs Supra-Alar poils de la mouche 7.
- Aligner la micropipette d'injection contenant le composé avec le centre de la ocelles trois situés sur la partie médiane postérieure de la tête, mais ne pas injecter encore (Figure 2, C).
- Avant l'injection du composé obtenir un enregistrement de base de la GF et GF TTM à DLM voies de la fibre géante (GFS, figure 1) par la stimulation du cerveau. Pour ce faire, activer les fibres géantes (GFS) avec 10 trains de 10 stimuli (40-50 mV) chaque donnée pour une durée de 0,03 ms à 100 Hz avec un délai de 1 seconde entre les trains 14,15 (figure 4). Le type sauvage vole shOuld être en mesure de suivre un-to-one à ce rythme de stimulation à la fois pour DLM et les voies de TTM. Jeter la volée si le GF et GF DLM aux voies TTM ne suivent pas la stimulation de 100 Hz à un ratio de 1:1.
- Mettre à la stimulation continue de la GF avec des impulsions uniques à 1 Hz (Figure 4).
- Rapidement brancher l'injecteur dans la boîte de contrôle. Même si le bruit de fond va interférer avec les enregistrements de stimulation 1 Hz, ne pas abandonner.
- Insérer soigneusement la micropipette injection dans la capsule de tête de la mouche juste au-dessous de la cuticule, et injecter la quantité désirée de composé dans l'hémolymphe de la volée tout en maintenant la stimulation de 1 Hz (figure 4). Grâce au système circulatoire ouvert de la mouche, le système nerveux tout entier seront exposés au composé en quelques secondes. Bien qu'un site d'injection particulier n'est pas critique pour délivrer des composés dans l'hémolymphe, on trouve la région des ocelles, qui sont de localiserd médial du côté le plus dorsale de la capsule céphalique, d'être un site pratique qui permet une injection facile qui mène à une distribution rapide et même du composé.
- Retirer immédiatement micropipette d'injection à partir du site d'injection et de débrancher l'injecteur du boîtier de commande tout en continuant le 1 Hz GF stimulation jusqu'à 1 min après l'injection (figure 4).
- Dans le but de révéler des effets plus subtils de composés sur la SFP, l'accent sur les fibres géantes (GFS) avec 10 trains de 10 stimuli à 100 Hz avec un délai de 1 seconde entre les trains. Continuer à tester la fonction des voies de GF avec ce paradigme toutes les 5 minutes jusqu'à 15 minutes (Figure 4). Toutefois, des intervalles plus courts ou plus longs périodes de surveillance sont également possibles.
- Afin de tester si le composé a un effet au niveau des jonctions neuromusculaires (NMJs) de la SFP, et peut-être affiner les effets du composé, procéder pour activer le moteur neurones directionTLY par la stimulation thoracique. Pour ce faire, retirez les électrodes de stimulation dans les yeux et de les remplacer sur les côtés antérieurs du thorax afin de stimuler les neurones moteurs avec 10 trains de 10 stimuli à 100 Hz.
Remarque: Les traces d'électrophysiologie montrés dans la vidéo ne correspondent pas à des effets de l'injection de colorant pur.
5. Les résultats représentatifs
Effet d'un antagoniste sur l'ISP aux DLM synapse du système fibre géant
Methyllycaconitine citrate (MLA) est un antagoniste nAChR qui est spécifique pour les sous-types a7 nAChR. Le PSI à DLMn synapse dans la voie de GF-DLM est dépendante de la sous-type Dα7 nAChR pour un bon fonctionnement, tandis que l'enlèvement génétique de sous-type Dα7 nAChR est sans effet sur la voie GF-MTT 5,6. Afin de démontrer la spécificité et la sensibilité de notre test nous avons injecté député à différentes concentrations (0, 0,02, 0,04, 0.08, 0,12 ng / mg, 46 nl injecté) dans la tête de l'animal (n = 10 pour un traitement Composé; n = 15 pour le traitement solution saline). Seules les mouches mâles (de type sauvage génotype sauvage 10E) ont été utilisés, et l'effet du composé a été contrôlée pour un total de 15 minutes après l'injection.
La figure 5 représente la différence entre les enregistrements de référence obtenues avant l'injection et ceux obtenus après l'injection en réponse à l'entraide judiciaire et la solution de contrôle solution saline. Nous avons trouvé que l'injection de l'entraide judiciaire pour conséquence l'incapacité de la voie de GF-DLM de suivre un-à-un à 100 Hz par des stimulations de l'ETG dans le cerveau alors que la voie GF-TTM est demeurée inchangée. (Figure 5, trace Haut et milieu, t-test effectué entre les contrôles salins [0 ng / mg] et les différentes concentrations de MLA à chaque point de temps, sauf si les données sont non-paramétrique [normalité et l'égalité des variances testé], sinon nous utilisons un test de Mann-Whitney. * p <0,001). Toutefois, une r one-to-oneÉPONSE de la DLM a été observée lorsque les neurones moteurs ont été stimulées directement (figure 5, trace en bas), ce qui démontre que la fonction de la JNM DLM et TTM n'est pas affectée par le député. MLA est apparu pour atteindre son maximum d'effet 1 minute après l'injection pour 0,04, 0,08 et 0,12 ng / mg de MLA injecté, comme aucun autre changement significatif n'a été noté au cours des 15 minutes suivantes de période d'essai. En outre, le composé a atteint un effet maximum à 0,08 ng / mg depuis plus fortes réponses n'ont pas été observés avec la dose plus élevée de 0,12 ng / mg.

Figure 1. Le diagramme de fibre géante du système de la fibre géante (GFS). Les fibres géantes (GFS, montré en rouge) synapse électrique (Gap jonctions) ainsi que chimiquement (cholinergique) sur un périphérique Interneuron synapse (PSI, en vert) et le neurone Tergo Muscle Trochanteral (TTMn, représenté en jaune) 5. Le PSI DLMn (dorsale neurone musculaire longitudinale, représentée en bleu) de connexion dépend de Dα7 sous-type nAChR 6. Enfin, la jonction neuromusculaire (JNM) de l'TTMn et le DLMn sur le saut (TTM, en violet) et les muscles de vol (DLM, en violet) est glutamatergique.
Remarque: Le GF à la connexion ISP est à la fois électrique et chimique. Cependant, chez les mutants shakB (qui ne disposent pas des jonctions communicantes), aucune réponse ne peut être enregistré à partir du DLM sur la stimulation de l'ETG dans le cerveau, ce qui démontre que le composant chimique en l'absence de connexions électriques ne suffit pas d'évoquer un potentiel d'action dans le PSI 5,16-18. Parce que le GF à la connexion ISP est dépendante de jonctions lacunaires, ce chiffre montre que la jonction GAP à la synapse pour des raisons de simplicité.

Figure 2.
Micromanipulateurs set-haut.
- Une modification de set-up d'un protocole déjà publié 14 est utilisée pour ajuster le micromanipulateur d'injection pour les enregistrements appariés avec GFS nanoinjections simultanées. La préparation à la mouche montée est orienté à la tête de la mouche vers l'expérimentateur. Le micromanipulateur d'injection (N ° 1) est placé en face de l'expérimentateur entre les deux manipulateurs pour les électrodes de stimulation (2 tungstène et # N ° 3). Les deux micromanipulateurs pour les électrodes d'enregistrement de verre (n ° 4 et 5) sont placés sur le côté gauche et droite, respectivement. Le micromanipulateur pour l'électrode de masse de tungstène (# 6) est placé le plus loin dans le dos soit sur le côté gauche (quels) ou sur le côté droit.
- Une vue rapprochée du sommet de l'arrangement des différentes électrodes et micropipette d'injection.
- Un correctement monté D. melanogaster empalé avec des électrodes et micropipette d'injection prête pour l'injection. Notez que l'animal corpsest monté avec son thorax horizontalement et ses ailes déployées. La cire est bien enroulé autour de son corps empêchant l'animal de se déplacer. En outre, l'électrode de masse (# 6, dans l'abdomen), les électrodes d'enregistrement de verre (# 4 et # 5, dans le thorax, a souligné par des contours noirs), et les électrodes de stimulation (# 2 et # 3, une dans chaque oeil ) sont empalés en place, comme décrit précédemment 14. La micropipette d'injection (# 1) est correctement aligné avec le centre de la ocelles trois (encerclée). Insertion de la micropipette d'injection doit être placée dans ce domaine.

Figure 3. Micropipette d'injection biseauté. Un schéma d'une micropipette correctement biseauté est montré ici. L'ouverture électrode doit être biseauté à un angle de 45 degrés et une ouverture entre 11 à 17 um. Une micropipette l'injection proprement dite en biseau est crucial pour une injection en douceur avec un minimum dAmage à la mouche.

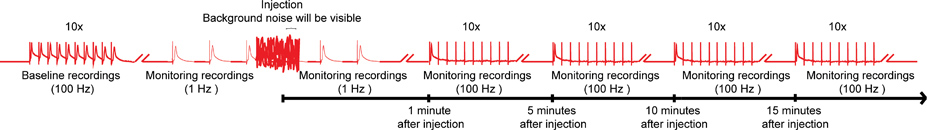
Figure 4. Schéma d'ensemble du protocole nanoinjection / électrophysiologie. Un diagramme représentant de l'ensemble du régime de la nanoinjection / protocole d'électrophysiologie. Commencez par l'obtention d'un enregistrement de base en stimulant les fibres géantes (GFS) à 100 Hz avec 10 trains de 10 stimuli chacune (un seul train illustré ici). Avant l'injection, de commencer les stimulations 1 Hz une seconde. Pendant le temps d'injection (injecteurs tout est branché sur le boîtier de commande), vous pourrez observer le bruit de fond important, cependant, ne pas interrompre les enregistrements. Après l'injection (et de l'injecteur est débranché de la boîte de commande), continuer la stimulation de 1 Hz pendant environ 1 minute de plus. Enfin, passez à souligner le GFS avec 10 trains de 10 stimuli à 100 Hz et de continuer à tester la fonction des voies de GF avec ce paradigme toutes les 5 minutes jusqu'à 15 minutes. Remarque: Les enregistrements wavant manipulé pour créer l'économie générale et ne représentent pas un résultat spécifique obtenu. Pas à l'échelle, et non pas toutes les traces sont visibles. Cliquez ici pour agrandir l'image .

Figure 5. Les effets de la MLA dans le SFG.
- Une représentation graphique des effets de la α7 nAChR antagoniste methyllycaconitine citrate (MLA) sur la voie de GF-DLM de la mouche des fruits à différentes concentrations (0, 0,02, 0,04, 0,08, 0,12 ng / mg n = 10 pour le traitement composé.; n = 15 pour le traitement salin). Seulement une minute après l'injection, un effet significatif et immédiat a été observée avec 0,04 ng / mg de MLA. Un effet significatif a également été vu avec 0.8ng/mg et 0.12ng/mg de l'entraide judiciaire à 100Hz stimulation de la SFP. Aucune différence significative n'a été notée entre les contrôles salines et de 0,02 ng / mg de MLA. En outre, aucun i changereffet n a été observée après une post-injection minute pendant le temps testé (15 minutes). Un t-test a été effectué entre les contrôles salins (0 ng / mg) et les différentes concentrations de MLA à chaque point dans le temps. Les niveaux sont signalés en moyenne + / - SEM, * p <0,001.
- Traces échantillon des réponses DLM à 100 Hz de stimulation. Trace du haut montre les réponses du muscle avant l'injection député lors de la stimulation GF dans le cerveau. Notez que le muscle est capable de répondre à chaque stimulus one-to-one à 100 Hz. Moyen de trace affiche les réponses de la DLM après injection de MLA (0,12 ng / mg). Notez que le muscle n'est pas en mesure de répondre à chaque stimulus one-to-one à 100 Hz. (Astérisques). Trace du bas montre les réponses de la DLM de la même préparation (0,12 ng / mg) lors de la stimulation directe des neurones moteurs dans le thorax. Parce que DLM répond one-to-one à 100 Hz avec une stimulation thoracique, l'échec de réponses avec des stimulations du cerveau peut être attribuée à l'cholinergique PSI-DLMn connexion.
- Une représentation graphique des effets à des concentrations différentes d'entraide judiciaire (0, 0,02, 0,04, 0,08, 0,12 ng / mg) sur la voie de GF-TTM. Aucun effet significatif n'a été observée entre une solution saline (0 ng / mg) et les injections de composés à aucun moment. Un t-test a été effectué entre les contrôles salins (0 ng / mg) et les différentes concentrations de MLA à chaque instant, * p <0,001.
- Traces échantillon des réponses TTM à 100 Hz de stimulation. Trace du haut montre les réponses du muscle avant l'injection député lors de l'activation GF avec une stimulation dans le cerveau. Notez que le muscle est capable de répondre à tous les stimuli à 100 Hz. Moyen trace montre les réponses de la DLM après injection de MLA (0,12 ng / mg). Les réponses du muscle à la stimulation TTM du GF dans le cerveau reste en tête-à-un. Trace du bas montre les réponses de la TTM de la même préparation à 100 Hz de stimulation des neurones moteurs dans le thorax (0,12 ng / mg).
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Le bio-essai nanoinjection / électrophysiologie présentée ici permet de dépistage rapide de composés dans le système nerveux de la mouche des fruits. C'est un roman dans la technique in vivo qui nécessite de petites quantités d'un composé de susciter un effet sur une variété de cibles moléculaires dans un circuit neuronal bien caractérisé. Cette méthode peut être utilisée pour tester la bioactivité des composés différents, à partir de toxines inconnues disponibles dans le commerce des agents pharmacologiques.
Ici, nous avons démontré la fonction de notre analyse en utilisant député, qui a eu un effet sur le système de fibre géante (SFP) de la mouche des fruits (Figure 5). Nous avons constaté que de façon sélective perturbé le GF voie DLM, mais pas le GF voie TTM. Activer les neurones moteurs directement via la stimulation thoracique a démontré que le défaut dans la voie de GF DLM n'est pas due à un dysfonctionnement de la jonction neuromusculaire (JNM), mais il était conforme à l'effi antagonistest de l'entraide judiciaire à des sous-types de nAChR Dα7 présenter à la synapse PSI-DLMn (Figure 1). Bien que le GF à la connexion TTMn s'est avéré cholinergique, il ne sait pas si Dα7 sous-unités nAChR sont présents à cette synapse. En outre, l'absence génétique de la choline acétyltransférase (Cha) gène ou la sous-type Dα7 nAChR (Dα7) gène ne perturbe pas la fonction de la connexion GF-TTMn raison de la présence simultanée d'une jonction électrique 5,6,17,19, 20, ce qui rend peu probable la voie d'être affectés par le député.
Après l'injection du composé, la solution devrait immédiatement plonger l'ensemble du système nerveux de l'animal grâce à son système circulatoire ouvert 21. Si elle est correctement injecté, le produit atteint généralement le thorax et l'abdomen en quelques secondes, mais une dispersion homogène peut prendre jusqu'à une minute. Toutefois, si le composé n'est pas injecté correctement dans l'hémolymphe (c.-à-injection de la micropipette trop deep va dans le tissu cérébral), puis plus lentement à travers la dispersion de l'animal est observé. Alors que colorant peut être utilisée pour pratiquer une technique d'injection appropriée, comme indiqué dans la vidéo, il est recommandé de ne pas co-injecter le colorant alimentaire bleu avec un composé à tester comme il peut modifier les propriétés du composé et donc sa bioactivité. En outre, puisque la plupart des solutions utilisées en tant que solvant sont de couleur claire (sérum physiologique, le DMSO, etc), il est difficile de voir si oui ou non le composé a été éjecté de l'aiguille d'injection. Par conséquent, lors de la dissolution d'un composé spécifique, il est important de veiller à ce que cela va complètement dans la solution; particules non dissoutes contraire vont rapidement obstruer l'embout d'injection, la prévention de toute éjection de fluide. En outre, bien que la dispersion composé peut être immédiate tout au long de l'hémolymphe, la réalisation des objectifs dans le système nerveux central, ainsi que pour atteindre son dosage maximal, peut prendre plus longtemps basée sur les propriétés chimiques du composé, comme la taille et Polarity, et sa capacité à pénétrer la barrière de la mouche du sang du cerveau. 22 Ainsi, il est important de surveiller les effets potentiels des composés inconnus quelques minutes après l'injection parce que différents composés peuvent avoir des variations dans les temps des effets d'apparition, ce qui peut augmenter au fil du temps, dans certains cas . Effets forts et immédiats du composé qui bloquent complètement la fonction des neurones peut être déjà vu avec les réponses déclenchées à 1 Hz, tandis que la stimulation de la SFP à des fréquences plus élevées (100 Hz) est utilisé pour détecter des effets plus subtils dus à des doses plus faibles ou puissance d'un composé. Si aucun effet n'est observé après l'injection du composé, il peut être due soit à la posologie de médicaments à petites ou le fait que le composé spécifique de la cible moléculaire n'est pas présent dans la SFP.
En outre, lors de l'utilisation du bio-essai présenté ici comme un outil de dépistage pour les nouveaux composés (tels que conotoxines), il est important de noter que le test est limité aux cibles moléculaires trouvés dans leGFS de la mouche. Bien que le dosage lui-même ne permet pas la localisation des cibles moléculaires réelles du composé injecté, il ne permet pour le rétrécissement des cibles potentielles au sein de la SFP. Des tests supplémentaires, tels que patch-clamp sur des neurones ou des muscles ou des études d'interaction avec des mutants génétiques de Drosophila melanogaster, peuvent être effectués pour déterminer la cible spécifique de ces composés. Enfin, le protocole d'enregistrement présenté a été conçu pour détecter des effets antagonistes sur la fonction de la SFP. Toutefois, le protocole d'enregistrement peut être facilement ajustée pour surveiller les effets agonistes en écoutant passivement des réponses induites par le composé plutôt que de tester si le SFP n'est pas en mesure de répondre de manière fiable lorsque le circuit est stimulée par l'expérimentateur.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Pas de conflits d'intérêt déclarés.
Acknowledgments
Nous tenons à remercier les membres du laboratoire Mari et le laboratoire Godenschwege, en particulier Aline Yonezawa, pour les commentaires et l'aide à ce protocole. Ce travail a été financé par l'Institut national des troubles neurologiques et des maladies de subvention à la bande FM R21NS06637 et TAG; AB a été financé par le nombre National Science prix de la Fondation 082925, URM: biologie intégrative pour les futurs chercheurs.
Materials
| Name | Company | Catalog Number | Comments |
| Recording glass electrodes: borosilicate glass capillaries | World Precision Instruments, Inc. | 1B100F-4 | 1.0mm OD, 0.58mm ID |
| Stimulator | Grass Technologies | Model S48 | |
| Amplifier | Getting Instruments, Inc. | Model 5A | |
| Data acquisition Software: Digidata | Molecular Devices | Model 1440A | |
| Data collection software: pCLAMP | Molecular Devices | Version 10 | |
| Stereomicroscope with fiber optic microscope ring illuminator | AmScope | SM-4T Model HL250-AR | |
| Dissecting scope for mounting | AmScope | SM-2TZ | |
| Kite Manual Micromanipulator & Tilting Base | World Precision Instruments, Inc. | Model # M3301 Kite: Model # KITE-M3-L | |
| Drosophila melanogaster Wild 10E genotype (wild type strain) | Bloomington Stock center | Stock # 3892 | |
| Vertical pipette puller | David Kopf Instruments | Model 700c | |
| Injection glass micropipettes: Borosilicate glass capillaries | World Precision Instruments, Inc. | Catalogue # 4878 | 1.14mm OD, 0.5mm ID |
| Silicon oil | Fisher Scientific | Catalogue # S159-500 | |
| Beveler | Sutter Instrument Co. | K.T. Brown Type Model # BV-10 | |
| Nanoliter2000 | World Precision Instruments, Inc. | Catalogue # B203XVY | |
| Blue food coloring | McCormick & Co. | N/A | Ingredients: Water, Propylene Glycol, FD&C Blue 1, and 0.1% Propylparaben (preservative). |
| Methyllycaconitine citrate (MLA) | Tocris Bioscience | Catalogue # 1029 | |
| Plastic wax sticks | Hygenic Corporation (Akron Ohio USA) |
References
- Koehn, F. E., Carter, G. T. The evolving role of natural products in drug discovery. Nat. Rev. Drug Discov. 4, 206-220 (2005).
- Miljanich, G. P. Ziconotide: neuronal calcium channel blocker for treating severe chronic pain. Curr. Med. Chem. 11, 3029-3040 (2004).
- Layer, R. T., Wagstaff, J. D., White, H. S. Conantokins: peptide antagonists of NMDA receptors. Curr. Med. Chem. 11, 3073-3084 (2004).
- Lewis, R. J. Conotoxins as selective inhibitors of neuronal ion channels, receptors and transporters. IUBMB Life. 56, 89-93 (2004).
- Allen, M. J., Godenschwege, T. A., Tanouye, M. A., Phelan, P. Making an escape: development and function of the Drosophila giant fibre system. Semin. Cell Dev. Biol. 17, 31-41 (2006).
- Fayyazuddin, A., Zaheer, M. A., Hiesinger, P. R., Bellen, H. J. The nicotinic acetylcholine receptor Dalpha7 is required for an escape behavior in Drosophila. PLoS biology. 4, e63 (2006).
- Jan, L. Y., Jan, Y. N. L-glutamate as an excitatory transmitter at the Drosophila larval neuromuscular junction. The Journal of physiology. 262, 215-236 (1976).
- Usherwood, P. N., Machili, P., Leaf, G. L-Glutamate at insect excitatory nerve-muscle synapses. Nature. 219, 1169-1172 (1968).
- Marrus, S. B., Portman, S. L., Allen, M. J., Moffat, K. G., DiAntonio, A. Differential localization of glutamate receptor subunits at the Drosophila neuromuscular junction. The Journal of neuroscience : the official journal of the Society for Neuroscience. 24, 1406-1415 (2004).
- Petersen, S. A., Fetter, R. D., Noordermeer, J. N., Goodman, C. S., DiAntonio, A. Genetic analysis of glutamate receptors in Drosophila reveals a retrograde signal regulating presynaptic transmitter release. Neuron. 19, 1237-1248 (1997).
- Qin, G. Four different subunits are essential for expressing the synaptic glutamate receptor at neuromuscular junctions of Drosophila. The Journal of neuroscience : the official journal of the Society for Neuroscience. 25, 3209-3218 (2005).
- Schuster, C. M. Molecular cloning of an invertebrate glutamate receptor subunit expressed in Drosophila muscle. Science. 254, 112-114 (1991).
- Tanouye, M. A., Wyman, R. J. Motor outputs of giant nerve fiber in Drosophila. Journal of. 44, 405-421 (1980).
- Augustin, H., Allen, M. J., Partridge, L. Electrophysiological Recordings from the Giant Fiber Pathway of D. melanogaster. J. Vis. Exp. (47), e2412 (2011).
- Allen, M. J., Godenschwege, T. Drosophila Neurobiology. Zhang, B., Freeman, M. R., Waddell, S. , Cold Spring Harbor Laboratory Press. 215-224 (2010).
- Blagburn, J. M., Alexopoulos, H., Davies, J. A., Bacon, J. P. Null mutation in shaking-B eliminates electrical, but not chemical, synapses in the Drosophila giant fiber system: a structural study. J. Comp. Neurol. 404, 449-458 (1999).
- Thomas, J. B., Wyman, R. J. Mutations altering synaptic connectivity between identified neurons in Drosophila. J. Neurosci. 4, 530-538 (1984).
- Baird, D. H., Schalet, A. P., Wyman, R. J. The Passover locus in Drosophila melanogaster: complex complementation and different effects on the giant fiber neural pathway. Genetics. 126, 1045-1059 (1990).
- Gorczyca, M., Hall, J. C. Identification of a cholinergic synapse in the giant fiber pathway of Drosophila using conditional mutations of acetylcholine synthesis. J. Neurogenet. 1, 289-313 (1984).
- Allen, M. J., Murphey, R. K. The chemical component of the mixed GF-TTMn synapse in Drosophila melanogaster uses acetylcholine as its neurotransmitter. The European journal of neuroscience. 26, 439-445 (2007).
- Mejia, M. A novel approach for in vivo screening of toxins using the Drosophila Giant Fiber circuit. Toxicon. 56, 1398-1407 (2010).
- Stork, T. Organization and function of the blood-brain barrier in Drosophila. J. Neurosci. 28, 587-597 (2008).

{kind=link}