

Summary
Una rápida
Abstract
Compuestos de cribado para la actividad in vivo puede ser utilizado como un primer paso para identificar a los candidatos que pueden ser desarrollados en 1,2 agentes farmacológicos. Hemos desarrollado un novedoso nanoinjection / electrofisiología ensayo que permite la detección de efectos moduladores bioactivos de los compuestos sobre la función de un circuito neuronal que media la respuesta de escape en la Drosophila melanogaster 3,4. Nuestro ensayo in vivo, que utiliza el sistema Drosophila gigante de fibra (EFP, Figura 1) permite la detección de diferentes tipos de compuestos, tales como pequeñas moléculas o péptidos, y requiere sólo cantidades mínimas para provocar un efecto. Además, el GFS Drosophila ofrece una gran variedad de posibles dianas moleculares en las neuronas o músculos. Las fibras gigantes (GFS) sinapsis eléctrica (uniones Gap), así como químicamente (colinérgico) en una sinapsis periférica Interneuron (PSI) y la neurona Tergo trocantéreo muscular (TTMn) 5 6. Por último, las uniones neuromusculares (UNM) de la TTMn y la DLMn con el salto (TTM) y los músculos de vuelo (DLM) son glutamatérgica 7-12. Aquí, demostramos cómo inyectar cantidades nanolitros de un compuesto, mientras que la obtención de registros intracelulares electrofisiológicos del sistema de fibra gigante 13 y cómo controlar los efectos del compuesto sobre la función de este circuito. Se muestra la especificidad de la prueba con citrato de methyllycaconitine (MLA), un antagonista de nAChR, que interrumpe a la ISP a DLMn conexión, pero no el GF TTMn conexión o la función de la UNM en el salto o los músculos del vuelo.
Antes de comenzar este video es muy importante que se observe con atención y se familiaricen con el video titulado "Jove registros electrofisiológicos de la vía fibra gigante de D. melanogaster "de Augustin et al 7, como el video que aquí se presenta pretende ser una expansión de esta técnica existente. Aquí se utiliza el método de los registros electrofisiológicos y el enfoque en detalle sólo en la suma de los pares de nanoinjections y la técnica de monitoreo.
Protocol
1. Electrofisiología Rig Set-up
- El equipo requerido para el equipo de perforación electrofisiología configuración se describe en detalle por Augustin et al. en este Diario 14. Por favor, consulte este artículo para obtener una explicación detallada de los aparatos electrofisiológicos necesarios.
- Modificar el equipo de electrofisiología se ha descrito anteriormente puesta a punto 14 mediante la adición de un micromanipulador sexto, que ocupa la nanoinjector. Para facilitar el acceso a la cabeza del animal que debe ser colocado entre los dos electrodos estimulantes micromanipuladores como se muestra en la Figura 2.
- Antes de comenzar el experimento, asegúrese de que tiene un rango confortable en todos los ejes de rotación con todos los micromanipulador, y que todos los electrodos, así como la micropipeta de inyección (ver paso 2) puede alcanzar el animal.
2. Nanoinjection Set-up
- El nanoinjection puesta en marcha requiere un Nanoliter2000 (World Precisisobre los instrumentos, Sarasota, FL, EE.UU.) o de tipo similar de inyección que permite inyecciones controladas en cantidades nanolitros.
- Preparar aguja de inyección utilizando las micropipetas de vidrio se suministran con el inyector tirando de ellos a una resistencia de 80-100 mW con un extractor de electrodo.
- Para las inyecciones suaves que se requiere para biselar las micropipetas a una apertura 11-17 m con un ángulo de 45 grados (Figura 3).
- Poco a poco rellenar la micropipeta de inyección de aceite sintético con una jeringa Hamilton con las instrucciones del manual de Nanoliter2000, asegurando que no queden burbujas de aire.
- Con cuidado, garantizar la micropipeta en el nanoinjector y prepararla para cargar el compuesto por vaciar el exceso de aceite según las instrucciones del manual de Nanoliter2000.
- Coloque el inyector en el micromanipulador y cargar el compuesto tal como se indica en el manual de Nanoliter2000. Asegúrese de que la punta de la micropipeta no se rompe durante esteprocedimiento.
- Establecer la cantidad deseada de nanolitros a ser inyectados en la caja de control del inyector según las instrucciones del manual de Nanoliter2000. Tenga en cuenta que la inyección de la cantidad total no debe superar los 100 nl. Hemos encontrado que las grandes cantidades de soluciones de control de solución salina puede afectar a la función del circuito de EFP.
- Es crítico para que el inyector desconectado de la caja de control durante la adquisición de grabación con la excepción de la inyección en sí, ya que la fuente de alimentación de la nanoinjector interfiere con las grabaciones, lo que es visible como el ruido de fondo (Figura 4). Sin embargo, no desconecte la fuente de alimentación a la caja de control, ya que volver a configurar.
3. Drosophila melanogaster Preparación
- Anestesie de 2 a 6 días de edad moscas con CO 2 o en el hielo a lo descrito previamente 14,15.
- Una vez inmóvil, utilizar un par de pinzas para transferir al animal a una red Wi plato pequeñoª cera blanda dental por recogerlo en sus piernas. Tenga en cuenta que una mosca de la fruta macho pesa alrededor de 1,0 mg y una mosca de la fruta hembra pesa alrededor de 1,2 mg, por lo tanto compuesto de-a-peso relación es diferente en las moscas macho en comparación con las mujeres. Por lo tanto, se recomienda utilizar sólo un género para los experimentos.
- Como se ha descrito previamente 14,15, cuidadosamente montar la parte dorsal mosca y asegurarse de que el tórax y la cabeza se inmovilizan con cera blanda dental colocado alrededor del cuerpo. Con cuidado, extender las alas de manera que ponen perpendicular al tórax (fig. 2, C). La mosca debe ser montado con el menor daño posible.
4. Nanoinjection emparejado / electrofisiología
- Coloque la marcha montados en la plataforma de electrofisiología con la cabeza hacia los electrodos de estimulación.
- Empalar el animal con el estimulante correspondiente suelo, y electrodos de registro como se ha descrito previamente 14,15 (Figura 2, C). A menos que se lo desea, coloque un electrodo en la grabación de DLM y el otro en el músculo TTM. El músculo DLM está situado en el tórax entre las anteriores dorso-centrales pelos y la línea media de la mosca. El músculo TTM se encuentra cerca de los accesorios de las alas, entre los anteriores y posteriores Supra-Alar pelos de la mosca de la 7.
- Alinear la micropipeta inyectable que contiene el compuesto con el centro de la ocelos tres situado en la porción media posterior de la cabeza, pero no se inyecte todavía (Figura 2, C).
- Antes de la inyección compuesto de obtener un registro al inicio del GF de TTM y GF de DLM vías del sistema de fibra gigante (EFP, Figura 1) a través de la estimulación cerebral. Para ello, activar las fibras gigantes (GFS), con 10 trenes de 10 estímulos (40-50 mV) cada una y durante un tiempo de 0,03 ms a 100 Hz con un retraso de 1 segundo entre las 14,15 trenes (Figura 4). De tipo salvaje vuela shOuld ser capaz de seguir uno a uno en este tipo de estimulación tanto para DLM y las vías de TTM. Deseche la marcha si el GF GF DLM y de las vías de TTM no siguen la estimulación de 100 Hz con una relación 1:1.
- Cambie a una estimulación continua del GF con pulsos individuales en 1 Hz (Figura 4).
- Rápidamente conecte el inyector en la caja de control. A pesar de que el ruido de fondo puede interferir con las grabaciones de 1 Hz de estimulación, no lo deje.
- Introducir cuidadosamente la micropipeta inyección en la cápsula de la cabeza de la mosca justo debajo de la cutícula, e inyectar la cantidad deseada de compuesto en la hemolinfa de la mosca mientras se mantiene la estimulación de 1 Hz (Figura 4). Debido al sistema circulatorio abierto de la mosca, todo el sistema nervioso será expuesto en el compuesto en cuestión de segundos. Aunque un sitio de inyección en particular no es crítico para entregar compuestos en la hemolinfa, se encuentra la región de la ocelos, que se localizand medial en la parte más dorsal de la cápsula de la cabeza, que es un sitio práctico que permite una inyección que lleva a la distribución rápida y uniforme del compuesto.
- Inmediatamente eliminar micropipeta inyección desde el sitio de inyección y desconectar el inyector de la caja de control mientras continúa la estimulación de 1 Hz GF hasta 1 min después de la inyección (Figura 4).
- Con el fin de revelar los efectos más sutiles de los compuestos en el modelo GFS, destacar las fibras gigantes (GFS), con 10 trenes de 10 estímulos a 100 Hz con un retraso de 1 segundo entre los trenes. Continuar para probar la función de las vías de GF con este paradigma cada 5 minutos hasta 15 minutos (Figura 4). Sin embargo, intervalos más cortos o más largos períodos de seguimiento son también posibles.
- Con el fin de probar si el compuesto tiene un efecto en las uniones neuromusculares (NMJs) de la EFP, y posiblemente reducir los efectos del compuesto, proceder a activar el motor neuronas direccióntemente por la estimulación torácica. Para ello, quite los electrodos de estimulación de los ojos y vuelva a colocar a los lados anterior del tórax con el fin de estimular las neuronas motoras con 10 trenes de 10 estímulos a 100 Hz.
Nota: Las huellas de electrofisiología se muestran en el video no corresponden a los efectos de la inyección de tinte puro.
5. Los resultados representativos
Efecto de un antagonista en el PSI a DLM sinapsis del sistema de fibra gigante
Methyllycaconitine citrato (MLA) es un antagonista nAChR que es específico para a7 subtipos Nachr. El PSI DLMn sinapsis en la vía de GF-DLM depende del subtipo de nAChR Dα7 para su correcto funcionamiento, mientras que la eliminación genética del subtipo nAChR Dα7 no tiene ningún efecto sobre la vía de GF-TTM 5,6. A fin de demostrar la especificidad y sensibilidad de nuestro ensayo se inyectó MLA a diferentes concentraciones (0, 0,02, 0,04, 0.08, 0,12 ng / mg, 46 nl inyectado) en la cabeza del animal (n = 10 por tratamiento con el compuesto, n = 15 para el tratamiento de solución salina). Sólo las moscas macho (de tipo silvestre genotipo 10E salvaje) fueron utilizados, y el efecto del compuesto se controló durante un total de 15 minutos después de la inyección.
La Figura 5 representa la diferencia entre las grabaciones de referencia obtenidos antes de la inyección y los obtenidos después de la inyección en respuesta a MLA y solución salina de control. Hemos encontrado que la inyección de MLA como resultado la incapacidad de la vía de GF-DLM para seguir uno a uno a 100 Hz por estimulaciones de la FC en el cerebro, mientras que la vía de GF-MTT no se vio afectado. (Figura 5, trazo superior y media, t-test realizado entre los controles de solución salina [0 ng / mg] y las diferentes concentraciones de MLA en cada punto de tiempo a menos que los datos no paramétricos [la normalidad y la igualdad de las diferencias prueba], de lo contrario se utiliza una prueba de rangos de Mann-Whitney Sum. * p <0,001). Sin embargo, una r uno-a-unoespuesta de la DLM se observó cuando las neuronas motoras fueron estimuladas directamente (Figura 5, traza de fondo), lo que demuestra que la función de la UNM DLM y TTM no se ve afectada por la MLA. MLA apareció para alcanzar su efecto máximo 1 minuto después de la inyección de 0,04, 0,08 y 0,12 ng / mg de MLA inyectado, como no hay cambios significativos adicionales se observaron durante los siguientes 15 minutos de período de prueba. Además, el compuesto alcanza un efecto máximo a 0,08 ng / mg, ya fuertes respuestas no se observaron con la dosis mayor de 0,12 ng / mg.

Figura 1. El gigante de fibra de diagrama del sistema del sistema de fibra gigante (EFP). Las fibras gigantes (GFS, que se muestra en rojo) sinapsis eléctrica (uniones Gap), así como químicamente (colinérgico) en un Interneuron sinapsis periférica (PSI, que se muestran en verde) y la neurona Tergo muscular trocantéreo (TTMn, que se muestra en amarillo) 5. El PSI DLMn (neurona dorsal del músculo longitudinal, que se muestra en azul) de conexión depende de Dα7 nAChR subtipo 6. Por último, la unión neuromuscular (UNM) de la TTMn y DLMn el salto a la (TTM, que se muestra en color morado) y los músculos del vuelo (DLM, que se muestra en morado) es glutamatérgica.
Nota: El Fondo Mundial para la conexión ISP es eléctrica y química. Sin embargo, en los mutantes que carecen de shakB (uniones), no hay respuesta puede ser grabado desde el DLM a la estimulación de la FC en el cerebro, lo que demuestra que el componente químico en la ausencia de conexiones eléctricas no es suficiente para provocar un potencial de acción en el PSI 5,16-18. Debido a que el GF a la conexión de la ISP es dependiente de las uniones, esta cifra sólo muestra la brecha de la salida en la sinapsis por razones de simplicidad.

Figura 2.
Micromanipuladores set-up.
- Una modificación puesta en marcha de un protocolo previamente publicado 14 se utiliza para ajustar el micromanipulador de inyección de pares de grabaciones de las EFP con nanoinjections simultáneas. La preparación marcha montada se orienta a la cabeza de la mosca hacia el experimentador. El micromanipulador inyección (# 1) se coloca delante de la experimentador entre los dos manipuladores para los electrodos de tungsteno estimulantes (# 2 y # 3). Los dos micromanipuladores para los electrodos de vidrio de grabación (# 4 y # 5) se colocan en el lado izquierdo y derecho, respectivamente. El micromanipulador para el electrodo de tierra de tungsteno (# 6) se coloca más lejos en la parte posterior ya sea en el lado izquierdo (mostrado aquí), o en el lado derecho.
- Una vista cercana de la parte superior de las disposiciones de los diversos electrodos y micropipeta de inyección.
- Un bien montado D. melanogaster empalado con electrodos y micropipeta inyección lista para la inyección. Observe que el cuerpo del animalse monta con su tórax en sentido horizontal y sus alas extendidas. La cera está firmemente envuelto alrededor de su cuerpo impidiendo que el animal se mueva. Además, el electrodo de tierra (# 6, en el abdomen), los electrodos de registro de vidrio (# 4 y # 5, en el tórax, resaltada en contornos oscuros) y los electrodos de estimulación (# 2 y # 3, una en cada ojo ) se empalado en su lugar, como se ha descrito previamente 14. La micropipeta de inyección (# 1) está correctamente alineado con el centro de la ocelos tres (círculo). La inserción de la micropipeta inyección debe ser colocado en esta área.

Figura 3. Micropipeta de inyección de biselado. Un diagrama de una micropipeta adecuadamente biselados se muestra aquí. La apertura del electrodo debe ser biselado a un ángulo de 45 grados y tiene una abertura entre 11 a 17 micras. Una micropipeta adecuada inyección de biselado es crucial para una inyección suave con un mínimo de dAmage a la mosca.

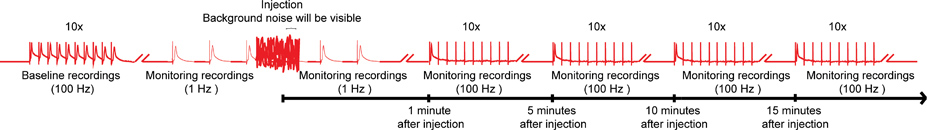
Figura 4. Esquema general del protocolo de nanoinjection / electrofisiología. Un esquema representativo del régimen general de la nanoinjection / protocolo de electrofisiología. Comience por la obtención de un registro al inicio mediante la estimulación de las fibras gigantes (GFS) a 100 Hz con 10 trenes de 10 estímulos cada uno (sólo uno de los trenes que se muestra aquí). Antes de la inyección, comienzan los estímulos 1 Hz un segundo de diferencia. Durante el tiempo de inyección (inyectores, mientras que está conectado a la caja de control), se observará el ruido de fondo considerable, sin embargo, no interrumpir las grabaciones. Después de la inyección (inyectores y está desconectado de la caja de control), continuar con la estimulación de 1 Hz durante 1 minuto más. Por último, procede destacar la FC con 10 trenes de 10 estímulos a 100 Hz, y poniendo a prueba la función de las vías de GF con este paradigma cada 5 minutos hasta 15 minutos. Nota: Las grabaciones wantes de manipular para crear el esquema general y no representan un resultado específico obtenido. No es a escala, no todos los restos se muestran. Haga clic aquí para ampliar la imagen .

Figura 5. Los efectos de la MLA en la EFP.
- Una representación gráfica de los efectos de la α7 antagonista de citrato de nAChR Methyllycaconitine (MLA) en el camino de GF-DLM de la mosca de la fruta a diferentes concentraciones (0, 0,02, 0,04, 0,08, 0,12 ng / mg de n = 10 por tratamiento con el compuesto.; n = 15 para el tratamiento de solución salina). Sólo un minuto después de la inyección, un efecto inmediato y significativo fue visto con 0,04 ng / mg de MLA. Un efecto significativo también fue visto con 0.8ng/mg y 0.12ng/mg del Eje de acción a 100 Hz con la estimulación de la FG. No se observaron diferencias significativas entre los controles de solución salina y 0,02 ng / mg de MLA. Además, no puedo cambiarn efecto fue visto después de un minuto después de la inyección durante el tiempo de prueba (15 minutos). Un t-test se realizó entre los controles de solución salina (0 ng / mg) y las diferentes concentraciones del Eje de acción en cada momento. Los niveles se presentan como media + / - SEM, * p <0,001.
- Rastros de muestra de las respuestas a 100 DLM Hz de estimulación. Trazo superior muestra las respuestas de los músculos antes de la inyección diputado a la estimulación de la FG en el cerebro. Observe que el músculo es capaz de responder a cada estímulo de uno a uno a 100 Hz. Medio traza muestra las respuestas de la DLM después de la inyección de MLA (0,12 ng / mg). Observe que el músculo no es capaz de responder a cada estímulo de uno a uno a 100 Hz. (Asteriscos). Traza de fondo muestra las respuestas del DLM de la misma preparación (0,12 ng / mg) a la estimulación directa de las neuronas motoras en el tórax. Debido DLM responde uno a uno a 100 Hz con estimulación torácica, la falta de respuestas con estimulaciones cerebrales se puede atribuir a la colinérgica PSI-DLMn conexión.
- Una representación gráfica de los efectos de diferentes concentraciones de MLA (0, 0,02, 0,04, 0,08, 0,12 ng / mg) en la vía GF-TTM. No se observaron efectos significativos entre la solución salina (0 ng / mg) y las inyecciones de compuestos en cualquier punto del tiempo. Un t-test se realizó entre los controles de solución salina (0 ng / mg) y las diferentes concentraciones del Eje de acción en cada momento, * p <0,001.
- Rastros de muestra de las respuestas a 100 TTM Hz de estimulación. Trazo superior muestra las respuestas del músculo antes de la inyección MLA tras la activación GF con la estimulación en el cerebro. Tenga en cuenta que el músculo es capaz de responder a todos los estímulos a 100 Hz. Medio traza muestra las respuestas de la DLM después de la inyección de MLA (0,12 ng / mg). Las respuestas del músculo MTT a la estimulación del GF en el cerebro siendo uno-a-uno. Traza de fondo muestra las respuestas de la MTT de la misma preparación a la estimulación de 100 Hz de las neuronas motoras en el tórax (0,12 ng / mg).
Subscription Required. Please recommend JoVE to your librarian.
Discussion
El bioensayo nanoinjection / electrofisiología que aquí se presenta permite una detección rápida de los compuestos en el sistema nervioso de la mosca de la fruta. Esta es una novedosa técnica in vivo que requiere pequeñas cantidades de un compuesto para provocar un efecto sobre una variedad de objetivos moleculares en un circuito bien caracterizado neuronal. Este método puede ser utilizado para probar la bioactividad de compuestos diferentes, de las toxinas desconocidas a agentes farmacológicos disponibles comercialmente.
Aquí hemos demostrado la función de nuestro ensayo con MLA, que tuvo un efecto sobre el sistema de fibra gigante (GFS) de la mosca de la fruta (Figura 5). Hemos encontrado que es selectivamente interrumpido el GF vía DLM, pero no el GF vía TTM. La activación de las neuronas motoras directamente a través de la estimulación torácica demostró que el defecto en la vía de GF DLM no se debió a una disfunción en la unión neuromuscular (UNM), pero era consistente con la efectividad antagónicat del Eje de acción en los subtipos de nAChR Dα7 presentes en la sinapsis PSI-DLMn (Figura 1). Aunque la conexión a GF TTMn ha demostrado ser colinérgico, se desconoce si Dα7 subunidades nAChR están presentes en esta sinapsis. Además, la ausencia genética de la colina acetiltransferasa (Car) gen o el subtipo nAChR Dα7 (Dα7) de genes no se perturbe la función de la conexión de GF-TTMn debido a la presencia simultánea de una unión eléctrica 5,6,17,19, 20, lo que hace que la vía improbable de ser afectados por MLA.
Después de la inyección compuesto, la solución inmediata debe sumergir todo el sistema nervioso del animal debido a su sistema circulatorio abierto 21. Si bien se inyecta, el compuesto por lo general alcanza el tórax y el abdomen en cuestión de segundos, pero una dispersión homogénea puede tardar hasta un minuto. Sin embargo, si el compuesto no se inyecta correctamente en la hemolinfa (es decir, la inyección de la micropipeta también deep entrar en el tejido cerebral), entonces más lenta la dispersión a través del animal que se observa. Mientras colorante puede ser utilizado para practicar una técnica de inyección adecuada como se muestra en el video, no se recomienda para co-inyectar el colorante azul con un compuesto a ensayar ya que pueden alterar las propiedades del compuesto y por lo tanto su bioactividad. Adicionalmente, puesto que la mayoría de las soluciones utilizadas como disolvente son de color claro (solución salina, DMSO, etc), es difícil de ver si el compuesto se expulsa de la aguja de inyección. Por lo tanto, cuando se disuelve un compuesto específico que es importante para asegurar que se va completamente en la solución; partículas no disueltas rápidamente de lo contrario se obstruirá la punta de inyección, evitando que cualquier eyección de líquido. Además, aunque la dispersión compuesto puede ser inmediata a lo largo de la hemolinfa, alcanzar los objetivos en el sistema nervioso central, así como alcanzar su dosis máxima, puede llevar más tiempo basado en las propiedades químicas del compuesto, como el tamaño y Polarity, y su capacidad para impregnar barrera de la mosca de sangre del cerebro. 22 Así, es importante para controlar los efectos potenciales de compuestos desconocidos varios minutos después de la inyección debido a diferentes compuestos pueden tener una variación en los tiempos de los efectos de aparición, que pueden aumentar con el tiempo, en algunos casos . Efectos fuertes e inmediatos del compuesto que bloquea completamente el funcionamiento de las neuronas ya se puede ver con las respuestas provocadas a 1 Hz, mientras que la estimulación de la EFP en frecuencias más altas (100 Hz) se utiliza para detectar los efectos más sutiles debido a la menor dosis o potencia de un compuesto. Si no se observan efectos después de la inyección compuesto que puede ser debido a la dosis del fármaco pequeño o el hecho de que el compuesto de blanco molecular específico no está presente en la EFP.
Además, cuando se utiliza el bioensayo se presenta aquí como una herramienta de detección para nuevos compuestos (tales como conotoxinas), es importante hacer notar que el ensayo se limita a las dianas moleculares encontradas en elEFP de la mosca. Aunque el propio ensayo no permite la localización de los objetivos reales moleculares del compuesto inyectado, sí permite el estrechamiento de los posibles objetivos dentro de la EFP. Los exámenes adicionales, tales como patch clamp en neuronas o en los músculos o los estudios genéticos la interacción con Drosophila melanogaster mutantes, se pueden hacer para determinar el objetivo específico de estos compuestos. Finalmente, el protocolo de grabación presentada fue diseñado para detectar los efectos antagonistas sobre la función de la EFP. Sin embargo, el protocolo de grabación puede ser fácilmente ajustado para supervisar los efectos agonistas por vigilancia pasiva para las respuestas inducidas por el compuesto en lugar de probar si la EFP no es capaz de responder de forma fiable cuando el circuito es estimulado por el experimentador.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
No hay conflictos de interés declarado.
Acknowledgments
Nos gustaría agradecer a los miembros del laboratorio y el laboratorio de Mari Godenschwege, en particular, Yonezawa Aline, por los comentarios y ayuda con este protocolo. Este trabajo fue financiado por el Instituto Nacional de Trastornos Neurológicos y Accidentes Cerebrovasculares de subvención R21NS06637 de FM y TAG, AB fue financiado por el número de la National Science premio de la Fundación 082925, MUR: Biología Integrativa para futuros investigadores.
Materials
| Name | Company | Catalog Number | Comments |
| Recording glass electrodes: borosilicate glass capillaries | World Precision Instruments, Inc. | 1B100F-4 | 1.0mm OD, 0.58mm ID |
| Stimulator | Grass Technologies | Model S48 | |
| Amplifier | Getting Instruments, Inc. | Model 5A | |
| Data acquisition Software: Digidata | Molecular Devices | Model 1440A | |
| Data collection software: pCLAMP | Molecular Devices | Version 10 | |
| Stereomicroscope with fiber optic microscope ring illuminator | AmScope | SM-4T Model HL250-AR | |
| Dissecting scope for mounting | AmScope | SM-2TZ | |
| Kite Manual Micromanipulator & Tilting Base | World Precision Instruments, Inc. | Model # M3301 Kite: Model # KITE-M3-L | |
| Drosophila melanogaster Wild 10E genotype (wild type strain) | Bloomington Stock center | Stock # 3892 | |
| Vertical pipette puller | David Kopf Instruments | Model 700c | |
| Injection glass micropipettes: Borosilicate glass capillaries | World Precision Instruments, Inc. | Catalogue # 4878 | 1.14mm OD, 0.5mm ID |
| Silicon oil | Fisher Scientific | Catalogue # S159-500 | |
| Beveler | Sutter Instrument Co. | K.T. Brown Type Model # BV-10 | |
| Nanoliter2000 | World Precision Instruments, Inc. | Catalogue # B203XVY | |
| Blue food coloring | McCormick & Co. | N/A | Ingredients: Water, Propylene Glycol, FD&C Blue 1, and 0.1% Propylparaben (preservative). |
| Methyllycaconitine citrate (MLA) | Tocris Bioscience | Catalogue # 1029 | |
| Plastic wax sticks | Hygenic Corporation (Akron Ohio USA) |
References
- Koehn, F. E., Carter, G. T. The evolving role of natural products in drug discovery. Nat. Rev. Drug Discov. 4, 206-220 (2005).
- Miljanich, G. P. Ziconotide: neuronal calcium channel blocker for treating severe chronic pain. Curr. Med. Chem. 11, 3029-3040 (2004).
- Layer, R. T., Wagstaff, J. D., White, H. S. Conantokins: peptide antagonists of NMDA receptors. Curr. Med. Chem. 11, 3073-3084 (2004).
- Lewis, R. J. Conotoxins as selective inhibitors of neuronal ion channels, receptors and transporters. IUBMB Life. 56, 89-93 (2004).
- Allen, M. J., Godenschwege, T. A., Tanouye, M. A., Phelan, P. Making an escape: development and function of the Drosophila giant fibre system. Semin. Cell Dev. Biol. 17, 31-41 (2006).
- Fayyazuddin, A., Zaheer, M. A., Hiesinger, P. R., Bellen, H. J. The nicotinic acetylcholine receptor Dalpha7 is required for an escape behavior in Drosophila. PLoS biology. 4, e63 (2006).
- Jan, L. Y., Jan, Y. N. L-glutamate as an excitatory transmitter at the Drosophila larval neuromuscular junction. The Journal of physiology. 262, 215-236 (1976).
- Usherwood, P. N., Machili, P., Leaf, G. L-Glutamate at insect excitatory nerve-muscle synapses. Nature. 219, 1169-1172 (1968).
- Marrus, S. B., Portman, S. L., Allen, M. J., Moffat, K. G., DiAntonio, A. Differential localization of glutamate receptor subunits at the Drosophila neuromuscular junction. The Journal of neuroscience : the official journal of the Society for Neuroscience. 24, 1406-1415 (2004).
- Petersen, S. A., Fetter, R. D., Noordermeer, J. N., Goodman, C. S., DiAntonio, A. Genetic analysis of glutamate receptors in Drosophila reveals a retrograde signal regulating presynaptic transmitter release. Neuron. 19, 1237-1248 (1997).
- Qin, G. Four different subunits are essential for expressing the synaptic glutamate receptor at neuromuscular junctions of Drosophila. The Journal of neuroscience : the official journal of the Society for Neuroscience. 25, 3209-3218 (2005).
- Schuster, C. M. Molecular cloning of an invertebrate glutamate receptor subunit expressed in Drosophila muscle. Science. 254, 112-114 (1991).
- Tanouye, M. A., Wyman, R. J. Motor outputs of giant nerve fiber in Drosophila. Journal of. 44, 405-421 (1980).
- Augustin, H., Allen, M. J., Partridge, L. Electrophysiological Recordings from the Giant Fiber Pathway of D. melanogaster. J. Vis. Exp. (47), e2412 (2011).
- Allen, M. J., Godenschwege, T. Drosophila Neurobiology. Zhang, B., Freeman, M. R., Waddell, S. , Cold Spring Harbor Laboratory Press. 215-224 (2010).
- Blagburn, J. M., Alexopoulos, H., Davies, J. A., Bacon, J. P. Null mutation in shaking-B eliminates electrical, but not chemical, synapses in the Drosophila giant fiber system: a structural study. J. Comp. Neurol. 404, 449-458 (1999).
- Thomas, J. B., Wyman, R. J. Mutations altering synaptic connectivity between identified neurons in Drosophila. J. Neurosci. 4, 530-538 (1984).
- Baird, D. H., Schalet, A. P., Wyman, R. J. The Passover locus in Drosophila melanogaster: complex complementation and different effects on the giant fiber neural pathway. Genetics. 126, 1045-1059 (1990).
- Gorczyca, M., Hall, J. C. Identification of a cholinergic synapse in the giant fiber pathway of Drosophila using conditional mutations of acetylcholine synthesis. J. Neurogenet. 1, 289-313 (1984).
- Allen, M. J., Murphey, R. K. The chemical component of the mixed GF-TTMn synapse in Drosophila melanogaster uses acetylcholine as its neurotransmitter. The European journal of neuroscience. 26, 439-445 (2007).
- Mejia, M. A novel approach for in vivo screening of toxins using the Drosophila Giant Fiber circuit. Toxicon. 56, 1398-1407 (2010).
- Stork, T. Organization and function of the blood-brain barrier in Drosophila. J. Neurosci. 28, 587-597 (2008).

{kind=link}