

Summary
En hurtig
Abstract
Screening af forbindelser for aktivitet in vivo, kan anvendes som et første trin til at identificere kandidater, der kan udvikles til farmakologiske midler 1,2. Vi udviklet et hidtil ukendt nanoinjection / elektrofysiologi assay, som tillader påvisning af bioaktive modulatoriske virkninger af forbindelser på funktionen af en neuronal kredsløb, der medierer flugtrespons i Drosophila melanogaster 3,4. Vores in vivo assay, der anvender Drosophila Giant Fiber System (GFS, figur 1) tillader screening af forskellige typer af forbindelser, såsom små molekyler eller peptider, og kræver kun minimale mængder til at fremkalde en virkning. Desuden tilbyder Drosophila GFS en lang række potentielle molekylære mål på neuroner eller muskler. De Giant Fibers (GFS) synapse elektrisk (gap junctions) samt kemisk (kolinerge) på en ekstern Synapsing Interneuron (PSI) og Tergo trochanterale Muscle neuron (TTMn) 5 6. Endelig neuromuskulære junctions (NMJ) af TTMn og DLMn med spring (TTM) og fly muskler (DLM) er glutamaterge 7-12. Her vil vi vise, hvordan man kan injicere nanoliter mængder af et stof, mens få elektrofysiologiske intracellulære optagelser fra Giant Fiber System 13 og hvordan man kan overvåge virkningerne af forbindelsen på funktionen af dette kredsløb. Vi viser specificiteten af assayet med methyllycaconitine citrat (MLA), en nAChR-antagonist, som forstyrrer PSI DLMn forbindelse, men ikke GF til TTMn forbindelse eller funktionen af NMJ ved hop eller flugt muskler.
Inden du begynder denne video er det vigtigt, at du nøje se og blive fortrolig med JOVE video med titlen "Elektrofysiologiske Optagelser fra Giant Fiber Pathway af D. melanogaster "fra Augustin et al 7, som videoen præsenteres her er tænkt som en udvidelse til den eksisterende teknik. Her bruger vi elektrofysiologiske optagelser metode og fokuserer i detaljer kun på tilsætning af de parrede nanoinjections og overvågning af teknik.
Protocol
1. Elektrofysiologi Rig Set-up
- Det krævede udstyr til elektrofysiologi riggen opsætning er beskrevet i detaljer af Augustin et al. i denne Journal 14. Der henvises til denne artikel for en detaljeret forklaring af de elektrofysiologiske nødvendige apparater.
- Rediger tidligere beskrevet elektrofysiologi rig set-up 14 ved at tilsætte 1/6 mikromanipulator, som holder nanoinjector. For nem adgang til dyrets hoved den skal placeres mellem de to stimulerende elektrode mikromanipulatorer som vist i figur 2.
- Før du starter eksperimentet, sikre, at du har et behageligt sortiment på alle akser i rotation med alle mikromanipulatorer, og at alle elektroder samt injektion mikropipette (se trin 2) kan nå dyret.
2. Nanoinjection Set-up
- Den nanoinjection set-up kræver en Nanoliter2000 (World Precisiom instrumenter,. Sarasota, FL, USA) eller lignende type af injektor, der tillader kontrollerede injektioner i nanoliter mængder
- Forbered injektionsnål ved hjælp af glas mikropipetter leveres med injektoren ved at trække dem til en modstand på 80-100 MQ med en elektrode aftrækker.
- Til glatte injektioner det er nødvendigt at affasningen af mikropipetter til en 11-17 um åbning i en 45 graders vinkel (fig. 3).
- Langsomt efterfylde indsprøjtning mikropipette med syntetisk olie med en Hamilton sprøjte som anvist af Nanoliter2000 manual, der sikrer, at ingen luftbobler er til stede.
- Forsigtigt sikre mikropipette på nanoinjector og forberede den til at indlæse forbindelse ved at tømme overskydende olie, som anvist af Nanoliter2000 manual.
- Sæt injektor på mikromanipulator og indlæse forbindelsen, som instrueret i Nanoliter2000 manual. At sikre, at spidsen af mikropipetten ikke gå itu under disseprocedure.
- Indstil den ønskede mængde nanoliters skal indsprøjtes i injektoren kontrol boks som anvist af Nanoliter2000 manual. Bemærk venligst, at det samlede beløb injicerede ikke bør overstige 100 nl. Vi fandt, at større mængder af saltvand kontrolopløsninger kan påvirke funktionen af GFS kredsløbet.
- Det er vigtigt at have injektoren taget ud fra styreboksen under optagelse erhvervelse med undtagelse af injektionen selv, eftersom strømmen af nanoinjector interfererer med optagelser, som er synlig som baggrundsstøj (figur 4). Du skal dog ikke afbryde strømforsyningen til kontrolboksen, som igen vil sætte den.
3. Drosophila melanogaster Fremstilling
- Bedøve 2 til 6 dage gamle fluer med CO2 eller på is som tidligere beskrevet 14,15.
- Gang ubevægelig, anvende en pincet til at overføre dyret til en lille plade with blød dentalvoks ved at tage den op fra sine ben. Bemærk, at en mandlig bananfluen vejer cirka 1,0 mg og en kvindelig bananfluen vejer omkring 1,2 mg, hvilket stof-til-kropsvægt forholdet er anderledes i mandlige versus kvindelige fluer. Derfor anbefales det at anvende én køn for eksperimenterne.
- Som tidligere beskrevet 14,15, forsigtigt montere fluen dorsale side op og sikre, at brystkassen og hovedet er immobiliseret med blødt dentalvoks placeret rundt i kroppen. Forsigtigt at vingerne ud, så at de lægger vinkelret på thorax (fig. 2, C). Fluen skal monteres med så lidt skade som muligt.
4. Parret Nanoinjection / elektrofysiologi
- Placer monteret flue på elektrofysiologi rig med sit hoved mod stimulerende elektroderne.
- Spidde dyret med det tilsvarende stimulerende, formalet og registrerende elektroder som tidligere beskrevet 14,15 (FIGUR 2, C). Medmindre andet er ønsket, sted en optagelse elektrode i DLM og den anden i TTM musklen. Det DLM muskel er placeret i brystkassen mellem de forreste dorso-Central hår og midterlinjen af fluen. Den TTM musklen er beliggende nær fløj vedhæftede filer, mellem den bageste og forreste Supra-Alar hår af fluen 7.
- Bringe injektionen mikropipetten indeholdende forbindelsen med centrum af de tre ocelli placeret på den mediale posteriore del af hovedet, men ikke injicere endnu (fig. 2, C).
- Før sammensatte injektion opnå en baseline optagelse af GF til TTM og GF til DLM veje i Giant Fiber System (GFS, figur 1) via brain stimulation. For at gøre dette, aktivere Giant Fibers (GFS) med 10 tog på 10 stimuli (40-50 mV) hver gives for en varighed på 0,03 ms ved 100 Hz med en 1 sekunders forsinkelse mellem togene 14,15 (Figur 4). Vildtype flyver shOuld være i stand til at følge en-til-en ved denne hastighed på stimulation for både DLM og TTM veje. Kassér fluen hvis GF til DLM og GF til TTM veje ikke følger 100 Hz stimulation i forholdet 1:1.
- Skifte til kontinuerlig stimulering af GF med enkelt pulser ved 1 Hz (figur 4).
- Hurtigt sætter injektoren ind i styreboksen. Selvom baggrundsstøj vil forstyrre med 1 Hz stimulering optagelser, ikke afbryde den.
- Indsæt forsigtigt injektion mikropipetten i hovedet kapsel fluen lige under cuticule og injicere den ønskede mængde forbindelse i hæmolymfe af flyve under opretholdelse af en Hz stimulation (figur 4). På grund af den åbne kredsløbssystem flue, vil hele nervesystemet udsættes for forbindelsen i sekunder. Selvom en bestemt injektionsstedet ikke er kritisk at afgive forbindelser til hæmolymfe, finder vi det område af ocelli, som lokalisererd medial på den mest dorsale side af hovedet kapsel, er et passende sted, der tillader en nem injektion, der fører til hurtig og jævn fordeling af forbindelsen.
- Straks at fjerne injektionsstedet mikropipette fra injektionsstedet og tag injektoren fra styreboksen samtidig fortsætte 1 Hz GF stimulation op til 1 min efter injektion (figur 4).
- For at afsløre mere subtile effekter af forbindelser på GFS, understrege Giant Fibers (GFS) med 10 tog på 10 stimuli ved 100 Hz med en 1 sekunders forsinkelse mellem togene. Fortsætter med at teste funktionen af GF veje dette paradigme hvert 5. minut op til 15 minutter (figur 4). Men kortere intervaller eller længere overvågning perioder er også mulige.
- For at teste, hvorvidt forbindelsen har en virkning på den neuromuskulære forbindelse (NMJs) af GFS og eventuelt indsnævre virkningerne af forbindelsen, gå til at aktivere motoren neuroner retningtly af thorax stimulation. Til dette fjerne de stimulerende elektroder fra øjnene og erstatte dem på de forreste sider af thorax med henblik på at stimulere de motoriske neuroner med 10 tog på 10 stimuli ved 100 Hz.
Bemærk: elektrofysiologi spor vist i videoen, ikke svarer til virkningerne af rene farvestof injektion.
5. Repræsentative resultater
Effekt af en antagonist på PSI til DLM synapse af Giant Fiber System
Methyllycaconitine citrat (MLA) er en nAChR-antagonist, der er specifik for a7 nAChR-undertype. PSI til DLMn synapse i GF-DLM pathway er afhængig af Dα7 nAChR-undertypen for korrekt funktion, mens genetiske fjernelse af Dα7 nAChR-undertypen har ingen virkning på GF-TTM pathway 5,6. For at demonstrere specificiteten og følsomheden af vort assay vi injiceret MLA ved forskellige koncentrationer (0, 0,02, 0,04, 0.08, 0,12 ng / mg, 46 nl injiceret) ind i hovedet af dyret (n = 10 pr behandling med forbindelsen, n = 15 for saltvand behandling). Kun mandlige fluer (for vildtype genotypen vild 10E) blev anvendt, og virkningen af forbindelsen blev overvåget i alt 15 minutter efter injektion.
Figur 5 viser forskellen mellem basislinieoptegnelser opnået før injektion, og de fremstilles efter injektion som reaktion på MLA og saltvandskontrol opløsning. Vi fandt, at injektion af MLA resulterede i manglende evne til GF-DLM pathway at følge en-til-en ved 100 Hz ved stimulering af GF'er i hjernen, mens GF-TTM pathway forblev upåvirket. (Figur 5, top-og mellemledere spor, t-test udført mellem saltvand kontrol [0 ng / mg] og de forskellige koncentrationer af MLA på hvert tidspunkt, medmindre data er ikke-parametrisk [normalitet og ens varianser testet], ellers bruger vi en Mann-Whitney Rank Sum Test. * p <0,001). En én-til-én response af DLM blev observeret, når de motoriske neuroner blev stimuleret direkte (figur 5, nederste spor), hvilket viser, at NMJ funktion DLM og TTM ikke påvirkes af MLA. MLA syntes at nå sin maksimale effekt 1 minut efter injektion af 0,04, 0,08 og 0,12 ng / mg MLA injiceret, som ikke yderligere signifikante ændringer blev noteret i de følgende 15 minutter testperiode. Endvidere kan forbindelsen opnået en maksimal virkning ved 0,08 ng / mg, idet større responser ikke blev observeret med den højere dosis på 0,12 ng / mg.

Figur 1. Den Giant Fiber System Diagram af Giant Fiber System (GFS). The Giant Fibers (GFS, vist med rødt) synapse elektrisk (gap junctions) samt kemisk (kolinerge) på en ekstern Synapsing Interneuron (PSI, der er vist i grøn) og den Tergo trochanterale Muscle neuron (TTMn, vist med gult) 5. PSI DLMn (Dorsal longitudinal muskel neuron, der er vist i blå) forbindelsen er afhængig Dα7 nAChR-undertypen 6. Endelig neuromuskulære junction (NMJ) af TTMn og DLMn på spring (TTM, vist i lilla) og fly muskler (DLM, vist i lilla) er glutamaterge.
Bemærk: GF til PSI forbindelse er både elektrisk og kemisk. I shakB mutanter (som mangler gap-junctions), kan der ikke noget svar noteres fra DLM efter stimulering af GF'er i hjernen, hvilket viser, at den kemiske komponent i fravær af elektriske forbindelser ikke er tilstrækkelig til at fremkalde et aktionspotentiale i PSI 5,16-18. Fordi GF til PSI-forbindelsen er gap junction afhængig, viser denne figur kun GAP krydset ved synapsen for enkelthedens skyld.

Figur 2.
Mikromanipulatorer set op.
- En modificeret opstilling af en tidligere offentliggjort protokol 14 bruges til at passe til injektion mikromanipulator for parrede GFS optagelser med samtidige nanoinjections. Den monterede fly Præparatet er orienteret med hovedet af fluen mod forsøgslederen. Injektion mikromanipulator (# 1) er placeret foran eksperimentatoren mellem de to manipulatorer for wolfram stimulerende elektroder (2 # og # 3). De to mikromanipulatorer til glas registrerende elektroder (# 4 og # 5) er anbragt på venstre og højre side, henholdsvis. Den mikromanipulator for wolfram jordforbindelseselektrode (# 6) er placeret længst på bagsiden enten på den venstre side (vist her) eller til højre.
- Et nærbillede fra toppen af arrangementer af de forskellige elektroder og injektion mikropipette.
- En korrekt monteret D. melanogaster spiddet med elektroder og injektion mikropipette klar til injektion. Bemærke, at dyrets kropmonteres med sin thorax vandret og vingerne spredes ud. Voks er forsvarligt viklet omkring sin krop forhindrer dyret i at bevæge sig. Derudover jordforbindelseselektrode (# 6 i maven), glas registrerende elektroder (# 4 og # 5 i thorax, fremhævet mørke linjer), og stimulerende elektroder (# 2 og # 3, et i hvert øje ) er spiddet på plads, som tidligere beskrevet 14. Injektion mikropipetten (# 1) er korrekt justeret med midten af de tre ocelli (cirkel). Indføring af injektionen mikropipetten skal placeres i dette område.

Figur 3. Skrå injektion mikropipette. Et diagram af en korrekt skrå mikropipette vises her. Elektroden Åbningen skal affaset ved en 45 graders vinkel og har en åbning mellem 11 til 17 um. En ordentlig skrå injektion mikropipette er afgørende for en glat injektion med minimal dAmage til fluen.

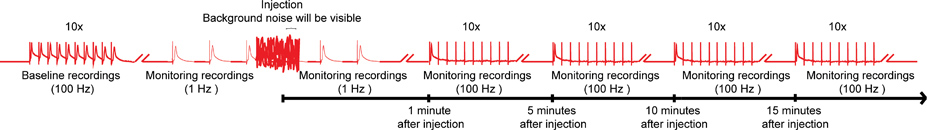
Figur 4. Samlet ordning for nanoinjection / elektrofysiologi-protokollen. En repræsentant diagram over den samlede ordning for nanoinjection / elektrofysiologi protokol. Start med at opnå en baseline optagelse ved at stimulere Giant Fibers (GFS) ved 100 Hz med 10 tog på 10 stimuli hver (kun ét tog vist her). Før injektion, begynder de 1 Hz stimulationer en sekund fra hinanden. Under injektion tid (mens injektor er sluttet til styreboksen), vil du observere betydelig baggrundsstøj, men ikke stoppe optagelserne. Efter injektion (og injektor er trukket fra kontrol boks), fortsætter 1 Hz stimulation for omkring 1 flere minutter. Endelig videre til understrege garantifonde med 10 tog på 10 stimuli ved 100 Hz og fortsætte med at teste funktionen af GF veje med dette paradigme hvert 5. minut op til 15 minutter. Bemærk: optagelser wførend manipuleret til at skabe den samlede ordning og ikke repræsenterer et bestemt resultat. Ikke at skala, er det ikke alle spor vises. Klik her for større billede .

Figur 5. Virkningerne af MLA i GFS.
- En grafisk afbildning af virkningerne af a7 nAChR-antagonisten Methyllycaconitine citrat (MLA) på GF-DLM pathway af frugtflue ved forskellige koncentrationer (0, 0,02, 0,04, 0,08, 0,12 ng / mg n = 10 pr behandling med forbindelsen. n = 15 for saltvand behandling). Kun et minut efter injektion, blev en betydelig og øjeblikkelig virkning ses med 0,04 ng / mg MLA. En væsentlig effekt blev også set med 0.8ng/mg og 0.12ng/mg i MLA ved 100 Hz stimulering af GFS. Ingen signifikant forskel blev bemærket mellem saltvandskontroller og 0,02 ng / mg MLA. Derudover ingen ændring in virkning blev set efter 1 minut efter injektion i det tidsrum testet (15 minutter). En t-test blev udført mellem saltvandskontroller (0 ng / mg) og forskellige koncentrationer af MLA på hvert tidspunkt. Niveauer er rapporteret som middelværdi + / - SEM, * p <0,001.
- Eksempler på spor af DLM reaktioner på 100 Hz stimulation. Øverste spor viser responser i musklen før MLA injektion af GF stimulering i hjernen. Bemærker, at musklen er i stand til at reagere på hver stimulus én-til-en ved 100 Hz. Mellemøsten spor viser svarene fra DLM efter MLA injektion (0,12 ng / mg). Bemærker, at musklen ikke er i stand til at reagere på hver stimulus én-til-en ved 100 Hz. (Asterisker). Nederst spor viser svarene fra DLM af samme præparat (0,12 ng / mg) ved direkte stimulering af de motoriske neuroner i brystkassen. Fordi DLM reagerer én-til-en ved 100 Hz med torakal stimulation, kan svigt af reaktioner med hjernetumorer stimuleringer tilskrives cholinerge PSI-DLMn forbindelse.
- En grafisk afbildning af virkningerne med forskellige MLA koncentrationer (0, 0,02, 0,04, 0,08, 0,12 ng / mg) på GF-TTM pathway. Ingen signifikante virkninger blev set mellem saltvand (0 ng / mg) og forbindelsen injektioner på ethvert tidspunkt. En t-test blev udført mellem saltvandskontroller (0 ng / mg) og forskellige koncentrationer af MLA på hvert tidspunkt, * p <0,001.
- Eksempler på spor af de TTM reaktioner på 100 Hz stimulation. Øverste spor viser responser i musklen før MLA injektion ved GF aktivering med stimulering af hjernen. Bemærk, at musklen er i stand til at reagere på alle stimuli ved 100 Hz. Mellemøsten spor viser svarene fra DLM efter MLA injektion (0,12 ng / mg). Reaktioner fra TTM musklen til stimulering af GF i hjernen forblive en-til-en. Nederste spor viser responser i TTM af samme præparat til 100 Hz stimulering af motorneuroner i thorax (0,12 ng / mg).
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Den nanoinjection / elektrofysiologi bioassay præsenteret her tillader en hurtig screening af forbindelser i nervesystem frugtflue. Dette er en hidtil ukendt in vivo teknik, der kræver små mængder af en forbindelse til at fremkalde en effekt på en række forskellige molekylære mål i en velkarakteriseret neuronal kredsløb. Denne fremgangsmåde kan anvendes til at teste den biologiske aktivitet af forskellige forbindelser, fra ukendte toksiner til kommercielt tilgængelige farmakologiske midler.
Her viste funktion i vores assay under anvendelse af MLA, som havde en virkning på Giant Fiber System (GFS) af frugtflue (figur 5). Vi fandt, at det selektivt forstyrret GF til DLM vej, men ikke GF at TTM vej. Aktivering af motorneuroner direkte via thorax stimulering viste, at defekten i GF til DLM pathway ikke skyldtes en dysfunktion ved den neuromuskulære junction (NMJ), men var i overensstemmelse med antagonistisk effektivt MLA ved Dα7 nAChR undertyper stede i PSI-DLMn synapse (figur 1). Selv om GF til TTMn forbindelse blev vist at være cholinerge, er det uvist, om Dα7 nAChR-underenheder er til stede ved denne synapser. Endvidere genetiske fravær af cholinacetyltransferase (Cha) genet eller Dα7 nAChR-undertypen (Dα7) genet ikke forstyrrer funktionen af GF-TTMn forbindelse på grund af den samtidige tilstedeværelse af et elektrisk forbindelseselement 5,6,17,19, 20, hvilket gør vej forventes at blive påvirket af MLA.
Efter forbindelsen injektion, bør opløsningen umiddelbart nedsænke hele nervesystemet af dyret på grund af sin åbne kredsløbet 21. Hvis de er korrekt injiceret forbindelsen sædvanligvis når thorax og maven inden for sekunder, men en homogen dispersion kan tage op til et minut. Hvis forbindelsen ikke injiceres korrekt i hæmolymfe (dvs. injektion af mikropipetten for deep gå ind i hjernevævet) og derefter langsommere dispersion i hele dyrets observeres. Medens farvestof kan anvendes til at udøve en korrekt injektionsteknik, som vist i videoen, anbefales det ikke at co-injicere blå levnedsmiddelfarve med en forbindelse, der skal testes, da det kan ændre egenskaberne af forbindelsen og dermed dens bioaktivitet. Derudover, eftersom de fleste opløsninger, der anvendes som opløsningsmiddel, er det klart i farve (saltvand, DMSO, etc.), er det vanskeligt at se, hvorvidt forbindelsen blev udstødt fra kanylen. Derfor, når opløsning af en specifik forbindelse er det vigtigt at sikre, at det går fuldstændigt i opløsning, ellers uopløste partikler vil hurtigt tilstoppe injektion spids, hvilket forhindrer enhver udstødning af væske. Selv forbindelse dispersion kan være direkte gennem hæmolymfe, når målene i centralnervesystemet, såvel som når sin maksimale dosering, kan tage længere tid baseret på forbindelsens kemiske egenskaber, såsom størrelse og Polarity og dens evne til at trænge fluen blod-hjerne-barrieren. 22 Det er således vigtigt at overvåge eventuelle virkninger af ukendte forbindelser adskillige minutter efter injektionen, da forskellige forbindelser kan have variationer i tid efter starten virkninger, hvilket kan øge tiden i nogle tilfælde . Stærke og umiddelbare virkninger af forbindelsen, der fuldstændigt blokere funktionen af neuroner allerede kan ses med det udløste responser på 1 Hz, mens stimulation af GFS ved højere frekvenser (100 Hz) anvendes til at detektere mere subtile virkninger forårsaget lavere dosis eller Styrken af en forbindelse. Hvis ingen virkninger observeres efter forbindelsen injektion kan skyldes enten lille lægemiddeldosis, eller at forbindelsen specifikke molekylære mål ikke er til stede i GFS.
Endvidere, ved anvendelse af bioassayet præsenteret her som et screeningsværktøj til hidtil ukendte forbindelser (såsom conotoxiner) er det vigtigt at bemærke, at assayet er begrænset til de molekylære mål, der findes iGFS af fluen. Selv om analysen i sig selv ikke tillader at placere de faktiske molekylære mål af den injicerede stof, betyder det mulighed for indsnævring af potentielle mål i GFS. Yderligere tests, såsom patch clamp på neuroner eller muskler eller genetiske interaktionsstudier med Drosophila melanogaster mutanter, kan gøres for at afgøre det specifikke mål for disse forbindelser. Endelig blev præsenteret optagelse protokol til påvisning antagonistiske virkninger på funktionen af GFS. Imidlertid kan optagelsen protokol kan let justeres for at kontrollere for agonistiske virkninger af passivt at overvåge for responser induceret af forbindelsen i stedet prøvning, hvis GFS ikke er i stand til at reagere pålideligt, når kredsløbet er stimuleret af eksperimentatoren.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Ingen interessekonflikter erklæret.
Acknowledgments
Vi vil gerne takke medlemmerne af Mari laboratoriet og Godenschwege laboratoriet, især Aline Yonezawa, for kommentarer og hjælpe med denne protokol. Dette arbejde blev finansieret af det nationale institut for neurologiske sygdomme og slagtilfælde tilskud R21NS06637 til FM og TAG, AB blev finansieret af National Science Foundation prisen nummer 082925, URM: Integrative Biology for fremtidens forskere.
Materials
| Name | Company | Catalog Number | Comments |
| Recording glass electrodes: borosilicate glass capillaries | World Precision Instruments, Inc. | 1B100F-4 | 1.0mm OD, 0.58mm ID |
| Stimulator | Grass Technologies | Model S48 | |
| Amplifier | Getting Instruments, Inc. | Model 5A | |
| Data acquisition Software: Digidata | Molecular Devices | Model 1440A | |
| Data collection software: pCLAMP | Molecular Devices | Version 10 | |
| Stereomicroscope with fiber optic microscope ring illuminator | AmScope | SM-4T Model HL250-AR | |
| Dissecting scope for mounting | AmScope | SM-2TZ | |
| Kite Manual Micromanipulator & Tilting Base | World Precision Instruments, Inc. | Model # M3301 Kite: Model # KITE-M3-L | |
| Drosophila melanogaster Wild 10E genotype (wild type strain) | Bloomington Stock center | Stock # 3892 | |
| Vertical pipette puller | David Kopf Instruments | Model 700c | |
| Injection glass micropipettes: Borosilicate glass capillaries | World Precision Instruments, Inc. | Catalogue # 4878 | 1.14mm OD, 0.5mm ID |
| Silicon oil | Fisher Scientific | Catalogue # S159-500 | |
| Beveler | Sutter Instrument Co. | K.T. Brown Type Model # BV-10 | |
| Nanoliter2000 | World Precision Instruments, Inc. | Catalogue # B203XVY | |
| Blue food coloring | McCormick & Co. | N/A | Ingredients: Water, Propylene Glycol, FD&C Blue 1, and 0.1% Propylparaben (preservative). |
| Methyllycaconitine citrate (MLA) | Tocris Bioscience | Catalogue # 1029 | |
| Plastic wax sticks | Hygenic Corporation (Akron Ohio USA) |
References
- Koehn, F. E., Carter, G. T. The evolving role of natural products in drug discovery. Nat. Rev. Drug Discov. 4, 206-220 (2005).
- Miljanich, G. P. Ziconotide: neuronal calcium channel blocker for treating severe chronic pain. Curr. Med. Chem. 11, 3029-3040 (2004).
- Layer, R. T., Wagstaff, J. D., White, H. S. Conantokins: peptide antagonists of NMDA receptors. Curr. Med. Chem. 11, 3073-3084 (2004).
- Lewis, R. J. Conotoxins as selective inhibitors of neuronal ion channels, receptors and transporters. IUBMB Life. 56, 89-93 (2004).
- Allen, M. J., Godenschwege, T. A., Tanouye, M. A., Phelan, P. Making an escape: development and function of the Drosophila giant fibre system. Semin. Cell Dev. Biol. 17, 31-41 (2006).
- Fayyazuddin, A., Zaheer, M. A., Hiesinger, P. R., Bellen, H. J. The nicotinic acetylcholine receptor Dalpha7 is required for an escape behavior in Drosophila. PLoS biology. 4, e63 (2006).
- Jan, L. Y., Jan, Y. N. L-glutamate as an excitatory transmitter at the Drosophila larval neuromuscular junction. The Journal of physiology. 262, 215-236 (1976).
- Usherwood, P. N., Machili, P., Leaf, G. L-Glutamate at insect excitatory nerve-muscle synapses. Nature. 219, 1169-1172 (1968).
- Marrus, S. B., Portman, S. L., Allen, M. J., Moffat, K. G., DiAntonio, A. Differential localization of glutamate receptor subunits at the Drosophila neuromuscular junction. The Journal of neuroscience : the official journal of the Society for Neuroscience. 24, 1406-1415 (2004).
- Petersen, S. A., Fetter, R. D., Noordermeer, J. N., Goodman, C. S., DiAntonio, A. Genetic analysis of glutamate receptors in Drosophila reveals a retrograde signal regulating presynaptic transmitter release. Neuron. 19, 1237-1248 (1997).
- Qin, G. Four different subunits are essential for expressing the synaptic glutamate receptor at neuromuscular junctions of Drosophila. The Journal of neuroscience : the official journal of the Society for Neuroscience. 25, 3209-3218 (2005).
- Schuster, C. M. Molecular cloning of an invertebrate glutamate receptor subunit expressed in Drosophila muscle. Science. 254, 112-114 (1991).
- Tanouye, M. A., Wyman, R. J. Motor outputs of giant nerve fiber in Drosophila. Journal of. 44, 405-421 (1980).
- Augustin, H., Allen, M. J., Partridge, L. Electrophysiological Recordings from the Giant Fiber Pathway of D. melanogaster. J. Vis. Exp. (47), e2412 (2011).
- Allen, M. J., Godenschwege, T. Drosophila Neurobiology. Zhang, B., Freeman, M. R., Waddell, S. , Cold Spring Harbor Laboratory Press. 215-224 (2010).
- Blagburn, J. M., Alexopoulos, H., Davies, J. A., Bacon, J. P. Null mutation in shaking-B eliminates electrical, but not chemical, synapses in the Drosophila giant fiber system: a structural study. J. Comp. Neurol. 404, 449-458 (1999).
- Thomas, J. B., Wyman, R. J. Mutations altering synaptic connectivity between identified neurons in Drosophila. J. Neurosci. 4, 530-538 (1984).
- Baird, D. H., Schalet, A. P., Wyman, R. J. The Passover locus in Drosophila melanogaster: complex complementation and different effects on the giant fiber neural pathway. Genetics. 126, 1045-1059 (1990).
- Gorczyca, M., Hall, J. C. Identification of a cholinergic synapse in the giant fiber pathway of Drosophila using conditional mutations of acetylcholine synthesis. J. Neurogenet. 1, 289-313 (1984).
- Allen, M. J., Murphey, R. K. The chemical component of the mixed GF-TTMn synapse in Drosophila melanogaster uses acetylcholine as its neurotransmitter. The European journal of neuroscience. 26, 439-445 (2007).
- Mejia, M. A novel approach for in vivo screening of toxins using the Drosophila Giant Fiber circuit. Toxicon. 56, 1398-1407 (2010).
- Stork, T. Organization and function of the blood-brain barrier in Drosophila. J. Neurosci. 28, 587-597 (2008).

{kind=link}