

Summary
우리는 우리가 살아있는 세포의 공 촛점 이미징에 의해 소포체 (ER)에 거주 형질 전환 형광 단백질의 분포와 이동을 조사하는 데 사용하는 이미징 방법을 설명합니다. 우리는 또한 ultrastructurally이 세포 내 구획의 구조에 대한 자신의 표현의 효과를 분석 할 수 있습니다.
Abstract
이러한 강렬한 interorganelle 분자 트래픽에도 불구하고 자신의 독특한 구성과 기능을 유지하지만 진핵 세포의 지질과 단백질은 지속적으로, 세포 구획 사이에 교환됩니다. 이 문서에 설명 된 기술은 생체 내에서 자신의 생리적 환경에서 단백질과 지질의 이동성과 인신 매매를 공부하는 강력한 수단입니다. (FLIP)을 photobleaching에에 (FRAP) 및 형광 손실을 photobleaching에 후 형광 복구 널리 엑소-세포 내 이입 통로, 세포 기관 또는 subcompartments 사이의 연속성, 단백질 복합체의 형성, 단백질 지역화보기를 통해 세포 내 인신 매매를 공부 라이브 세포 이미징 기술을 사용하는 지질 마이크로 도메인에서 모두의 생리적 및 병리 적 조건에서 관찰 될 수있다. 이러한 방법의 한계로 인해 형광 융합 단백질의 용도에 주로, 그들의 잠재적 인 단점 과발현 인공적 포함세포에서 이온과 태그 및 기본 단백질의 접힘 및 현지화의 차이의 가능성. 마지막으로, 광학 현미경 (약 200nm)의 해상도의 한계로 스트레스 세포에서 유래 할 수있는 ER 또는 특정 subcompartments의 미세 구조의 조사를 허용하지 않는 (즉, 저산소증, 약물 투여, 횡단의 과발현 ER 상주 단백질) 또는 병적 인 조건에서, 우리는 미세 배양 형질 세포의 라이브 세포 이미징을 결합하여 투과 전자 현미경을 기준으로 분석한다.
Introduction
녹색 형광 단백질 (GFP) 및 스펙트럼 변형, 형광 현미경의 평행 개발의 발견은 세포에있는 단백질 행동의 조사를 위해 완전히 새로운 길을 열었습니다. 때문에 강렬한 조명 아래에서 자신의 형광을 소화하는 형광 물질의 고유 용량이 가능하다 (FLIP)을 photobleaching에에 (FRAP) 및 형광 손실을 photobleaching에 후 같은 형광 복구와 같은 기술은, 공 초점 라이브 세포 이미징을 기반으로하고의 사용은 형질 형광 융합 단백질 1-3. 그들은 널리 자신의 기능 4와 관련된 중요한 단서를 공개 할 수있는 단백질의 현지화뿐만 아니라, 자신의 이동성과 기공을 갖는 전송뿐만 아니라,을 평가하는 데 사용됩니다.
진핵 세포의 고유 기능은 특정 지질과 단백질 성분이 세포 내 구획의 존재입니다. 세포 소기관은 물리적으로 isolat와 있지만ED는 그들이 서로 통신 및 셀룰러 항상성을 유지하기 위해 분자 구성 요소를 공유 할 필요가있다. 분비 경로는 ER에서 합성 단백질과 지질이 그들의 기능을 발휘하는 정확한 최종 목적지에 도달 할 수 있음을 보장합니다. 세포 내 소기관 또한 분자 (지질)에 직접 구획 사이에서 교환 될 수 있도록 동적 접촉 위치에 의해 접속 될 수있다. 또한, 많은 단백질에 큰 이종 체 단지에서 조립 또는 기능적으로 활성화 될 수 또는 최종 목적지로 이송하기 위해 특정 지질 종 (지질 뗏목 / 마이크로 도메인)와 관련있다. 이러한 생물학적 측면 모두는 크게 단백질의 역학적 특성에 영향을 미치는, 따라서 적절하게 아래에 설명 된 기술에 의해 조사 할 수있다.
우리 그룹은 ER의 아키텍처와 다른 서브를 연구하기 위해 널리 FRAP를 사용하고 전자 현미경과 결합 플립도메인. ER은 분비 경로의 제 1 국이고, 단백질과 지질 5 선별에 중요한 역할을한다. 그것은 누구의 고유 한 하위 도메인의 여러 가지 기능 (예 : 단백질과 지질의 생합성 및 인신 매매, 단백질 폴딩, 칼슘 2 + 저장 및 해제, 및 생체 이물 대사)를 반영하는 매우 역동적 인 세포 기관이다. 그들이 공간적, 형태 학적, 그리고 기능적으로 구별 그러나, 비록 이러한 도메인은 서로 연속하고, 그들의 상대적인 풍부함 생리 및 병리 적 조건하에 세포에서 변형 될 수있다. 가장 잘 알려진 및 ER의 보통 공간적으로 분리 된 도메인들은 핵막 및 매끄럽고 거친 ER이며 그러나 다른 사람은 ER 구조는 다양한 종류의 세포에서보다 정교한 구조 및 3 차원 조직이 있다는 것을 증명하고있다 또한 저산소증, 약물과 같은 스트레스 자극에 의해 유도 될 수있는 생리 학적 조건 하에서 조직관리, 또는 ER 상주 횡단 단백질 2,6의 과발현 (및 참조 안에).
우리는 또한 최근에 인간의 질병 1,7의 세포 모델에서 이러한 구조의 존재를 증명하고있다. 부드러운 ER의 누적 cisternae에서 발생 그들은 또한 그들의 아키텍처에 기초 karmellae, 얇은 판자, 및 결정상 ER로 알려져 있지만, 그들은 2003 년 6 조직 부드러운 소포체 (OSER)의 집단 이름이 부여 된, 같은 자신 크기는 다를 수 있습니다. 세포는 꼬리 고정 (TA) ER 상주 단백질 (D의 EGFP-ER), 트랜스에서 GFP의 약한 이합체 경향이 극적으로 ER의 조직과 구조를 변경의 세포질 영역에 융합 GFP 형질 전환 된 후. FRAP 및 플립 실험은 D EGFP-ER은 OSERs 내에서 확산 자유롭게 것으로 나타났다하고 OSER에 망상 ER에서 이동하고 그 반대의 경우도 마찬가지 <사실/ 엠> 집계 주변의 망상 ER 연속 있음을 나타냅니다. 미세 분석은 우리가 나노 수준에서 OSER 아키텍처와 조직에 대한 자세한 설명과 함께 형광 데이터의 상관 관계를 할 수있다 : OSERs은 항상 부드러운 ER의 쌍 cisternae의 스택으로 구성되어 있지만, 이러한 정기적 사인 배치로 공간 조직의 다양한 형태를 가질 수 배열이나 윤 생체, 또는 육각형의 "결정체"관의 배열. 이러한 재 배열들은 생리적 조건 9 저산소증 10, 약물 치료 11 암 9로 다음과 같은 스트레스에 따라 세포에서 발견 된 것처럼, 미세 마커 상당한 잠재력을 가지고있다, 입방 형태학 8로 이어집니다.
GFP 융합 단백질을 사용하여이 제 데모 후에, 우리는 약물 치료 12 ASSE에 응답하여 ER 도메인의 증식을 분석하기 위해 이미징 실험을 사용SS 세포 13 oligomerise하고, 그 병원성 1,8 관련된 수도 ER 계 세포 응집체의 형성에 돌연변이, ALS TA-결합 단백질의 역할을 조사하기 위해 형광 단백질의 경향. 그것은 (많은 신경 변성 질환 (14)에서 발생하는) 세포 응집체의 형성이 독성 돌연변이 단백질 및 주변 셀 요소 (15) 사이의 상호 작용을 방지하기위한 보호 메커니즘이 될 수 있다는 것을 제안하고있다.
다음은 그의 C-말단 소수성 도메인은 ER의 세포막 내로 삽입되고, 자신의 동적 거동의 분석 및 ER 도메인에서 자신의 과발현의 효과 구조를 조사하는 광학 및 전자 현미경 방법의 조합에 대하여 설명한다 배양 된 세포의 구조 (실험 프로토콜의 흐름도는 그림 1 참조).
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. ER 형광 단백질 플라스미드, 세포 배양 및 형질
- 본 연구에서 사용 된 플라스미드는 쥐 시토크롬 B의 ER 이소 형의 꼬리 영역과의 C-말단에 융합 된 GFP의 향상된 버전으로 구성 (5) 링커 서열을 통해 (B 등 (5) 여기 기함). 꼬리 영역 (5), 업스트림 및 다운 스트림 극성 시퀀스에 의해 측면에 17 잔류 TMD (횡단 도메인), (UPS 및 DPS) 등의 기본 B의 트립신 절단 후 관련 막 남아있는 전체 시퀀스 (Pro94-Asp134)를 포함 . 링커 [(의 Gly) 4 빼앗아] 3이어서 MYC 에피토프로 구성하고, 전체 cDNA를 포유 동물 발현 벡터의 Hind3 된 pcDNA3-Xba1 사이트에 삽입된다. 이 플라스미드의 구성 정보가 GFP ER-16로 언급되는 이전의 간행물에 기재되어있다.
- 10 % 죽은 태아의 보충 둘 베코의 수정 이글 중간 (DMEM)의 COS-7 세포를 성장L 소 혈청, 2 밀리미터 L-글루타민, 37 ° C에서 인큐베이터에서 1 % 페니실린 / 스트렙토 마이신 및 10 % CO 2.
- 형질. 제조자에 의해 설명 된대로 판 3 × 10 5 6 - 웰 플레이트에 둥근 유리 커버 슬립에 세포와 다음 날에, JetPEI 시스템으로 형질. 최적 JetPEI / DNA 비율은 사용 된 플라스미드 및 세포주에 따라 최대 형질 감염 효율을 확립하기 위해 시험되었다 유의 : 우리의 경우, JetPEI는 2:1의 DNA 비율 70-80 % 년 형질 감염 효율에 이르게한다.
2. 라이브 형광 스캐닝 공 초점 현미경
- 라이브 세포 이미징. 형질 세포가 W / 10 % FBS, 2 밀리미터 L-글루타민, 1 % 펜 / 연쇄상 구균, 25 mM의 HEPES와 보충 페놀 레드, O를 DMEM 가득 24mm 커버 슬립의 강 문화 전지 실에 파종 된에 커버 슬립을 넣어 , photobleaching에에서 샘플을 방지하기 위해 50 ㎍ / ㎖의 사이클로 헥시 미드 1:100 OxyFluor. SP5 공 초점 microscopD의 GFP-ER은 488 nm의 레이저와 50분의 525 대역 통과 방출 필터를 사용하여 시각화되고으로 온도가 조절 CO 2 배양기 (37 ° C, 5 % CO 2)를 탑재 전자는 라이브 셀 이미징 실험에 사용됩니다.
- FRAP (을) photobleaching에 후 형광 복구. OSER 구조에 대응하는, 또한 (ROI)의 영역을 그리고 그것을 (샘플에서 5.5-6 μW에 대응하는, 30 mW의 아르곤 레이저의 100 %) 20 반복 및 488 nm의 조합을 사용하여, 405 nm의 표백 우리의 경험에서, 효율적이고 신속한 광표백에 이르게, (샘플에서 11.6 μW에 해당하는 30 mW의 다이오드 레이저 (405)의 60 %,) 레이저.
- 10 분 (픽셀 시간 = 1.61 마이크로 초 / PX)에 대해 하나의 프레임마다 10 초를 복용하여 표백의 ROI에 형광의 회복을 기록합니다.
- (FLIP)을 photobleaching에 형광 손실. OSER 구조에 따른 투자 수익 (ROI), 전술 한 바와 같이 표백제를 그립니다. 표백은 매 30 초를 반복,사후 표백 이미지는 30 분 (픽셀 시간 = 1.61 마이크로 초 / PX)마다 10 초를 기록한다.
- FRAP 및 플립 분석. 모든 이미지는 ImageJ에 소프트웨어 (사용하여 분석 http://rsbweb.nih.gov/ij/download.html을 ). FRAP 실험에서, 표백 투자 수익 (ROI)의 형광 복구는 시간과 항상 시간에 일정하게 체크 표백 셀의 총 형광, 정규화를 통해 측정된다.
- FLIP 실험의 경우, 표백 OSER 및 전체 세포를 덮고 외부의 투자 수익 (ROI)을 그립니다. 시간이 지남에 따라 그 형광 강도를 측정하고 영상 자체에 기인 한 형광의 어떤 저하를 보정하기 위해 표백 셀에 그려진 ROI의 형광 수준으로 정상화.
- 모든 실험에서의 ROI의 형광 강도로부터 (세포 외부의 영역에서 결정) 배경 신호를 감산. 마지막으로, 할머니를 사용하여 결과를 플롯aphPad 프리즘 소프트웨어.
3. 투과 전자 현미경에 의한 미세 구조 분석
시약의 많은 독성을 감안할 때, 모든 절차는 적절한 실험실 입고 수행되어야한다. 흄 후드 코트와 장갑.
- 페트리 접시에서 커버 슬립을 제거한 후, 실온에서 10 분 동안 0.1 M cacodylate 완충액, pH 7.4의 여과 2 % 글루 타르 알데히드를 사용하는 단일 층으로서 접시의 바닥에 성장 남은 세포를 고정한다.
- 테플론 스크레이퍼를 사용하여 세포를 긁고 1.5 ㎖의 에펜 도르프 튜브에 전송할 수 있습니다. 10 분간 9,000 g에서 원심 분리에 의해 세포 펠렛. , 뜨는을 제거 신선한 정착액을 추가하고, 4 ℃에서 하룻밤 남겨
- 버퍼와 펠렛을 씻으, 실온에서 1 시간 동안 cacodylate 버퍼에 1 % 오스뮴의 용액으로 후 고정합니다.
- 를 MilliQ의 물로 씻어하고있는 블록 얼룩 엔20 ~ 60 사이의 분 동안 증류수에 1 % 우라 닐 아세테이트.
- 에탄올 시리즈 (70 %, 80 %, 90 %, 100 %, 10 분마다 100 %) 증가에 샘플을 탈수하고, (각 15 분) 프로필렌 옥사이드에 짧게 두번 세척 하였다.
- (2 시간 내지 하룻밤) 프로필렌 옥 시드 +의 Epon (1:1)의 혼합물에 샘플을 침투.
- 최소 24 시간 동안 60 ° C에서 경화의 EPON 에폭시 수지에 포함.
- 제 60 ~ 70 ㎚의 두께를 갖는 섹션을 구하는 45 ° 다이아몬드 나이프 탑재 인해 Ultramicrotome LEICA UC6를 사용하여 수동으로 트리밍 수지 진영. 300 메쉬 구리 그리드에 대한 부분을 수집합니다.
- 우라 닐 아세테이트 (20 분)의 포화 용액으로 그리드 섹션을 얼룩과 구연산 (7 분)를 이끌어, 철저하게 이중 증류수 여과 물을 침지하여 그리드를 세척하고 실온에서 건조 할 수 있습니다.
- 더러워 그리드는 TECNAI G2 투과형 전자 현미경을 사용하여 관찰되고, 이미지는 BOTT를 사용하여 캡처되는(일반적으로 6,000-39,000 X에 이르기까지) 다른 최종 배율에서 OM-마운트 CCD 카메라.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
그림 2는 단백질 이동의 예 FRAP 연구 결과를 보여줍니다. D EGFP-ER 단백질의 이동성은 표백 OSERs에 photobleaching에 후 빠른 형광 복구에 의해 증명된다. 정량 분석을 위해, 하프 타임 및 모바일 분획은 다음 방정식 단지 수 피팅에 의해 실험적으로 측정 된 데이터로부터 유도 하였다 :
F (t) = F 포스트 + (F의 REC-F의 게시물) (1-e-t / τ)
F 포스트 photobleaching에 후 형광 신호이고, F의 REC 등록시 t, 표백 후의 도달 시간 상수 τ되는 형광 극대 복구 값이다.
형광 복구를 변경하고, 결과적으로, 단백질의 이동성을 분석 할 수 포화 픽셀 않고 이미지를 획득의 중요성을 양해 해 주시기 바랍니다. 그것은 내가항상 초점 평면 내의 이미지 획득 또는 작은 변화 동안 표백에 의한 형광 강도의 변화를 고려하기 위해 동일 셀의 전체에 형광 표백 ROI의 형광 신호를 정규화하는 것도 필수적들.
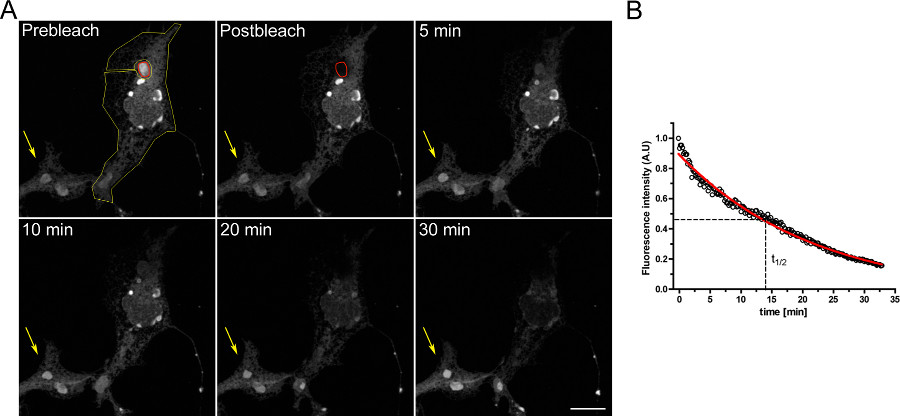
세포 내 구획들 사이의 연속성을 연구 FLIP 실험의 예는도 3에 도시된다. OSER 도메인이 지속적으로 표백 ER의 진보적 비우는에 의해 입증으로 OSERs 물리적 ER의 나머지 부분과 연결되어있다.
적절한 분석을 위해, 포화 픽셀의 취득 (위 참조) 피해야한다, 또한, 획득 매개 변수는 이미지 수집으로 인해 photobleaching에 방지하기 위해 가능한 한 낮은 레이저 힘을 설정해야합니다. 이러한 이유로 강하게 화상 표백 C의 형광 신호를 정규화하기 위해 사용될 동일 필드 표백 셀 추천엘.
모든 실험 인해 단백질 생합성 어떤 ER 형광 신호의 증가 (및 따라서 전체 형광)을 피하기 위하여, 사이클로 헥시 미드, 번역 억제제의 존재하에 수행 할 수있다.
투과 전자 현미경은 디 EGFP-ER로 형질 감염된 배양 된 세포에서 관찰 된 형광 골재 매끄러운의 패치를 나타낸다는 것을 입증하고 공간적으로 그 패턴에 기초하여 분류되어 잘 정의 된 3 차원 형상으로 스스로를 구성 ER의 cisternae 평평 : 종종 선형 또는 곡선 스택 ( 정현파 ER의 영역으로 연속 될 수있는 핵 봉투, 도시하지 않음) (그림 4A 및 B) (그림 4A)와 관련, 일부 지역에서 세포막이 사각형 또는 육각형 대칭 (결정상 ER과 격자로 구성되어, 도시하지 않음 ). 인접 cisternae는의 얇은 층으로 분리되어 약간 전자밀도 세포질 두께 약 11 nm의 그 집계를 둘러싼 세포질 연속이다.

그림 1. 실험 절차의 흐름도. 배양 세포가 처음으로 관심의 형광 융합 단백질을 발현 과다에 위해 jetPEI (프로토콜 참조)로 형질됩니다. 24 시간 후, 라이브 형질 전환 세포를 시각화하고 FRAP 및 플립 실험은 온도가 제어되는 CO 2 배양기를 갖춘 공 초점 현미경을 사용하여 수행하며, 기록 된 이미지는 수출과 적절한 소프트웨어 (예 : ImageJ에)를 사용하여 분석 하였다. 미세 분석을 위해, 형질 감염된 세포를 고정 펠릿 및 EPON 에폭시 수지 진영에 매립된다. 얇은 부분은 C에서 수집, 다이아몬드 나이프를 사용하여 얻을 수있다opper 그리드 및 투과 전자 현미경으로 관찰. 큰 그림을 보려면 여기를 클릭하십시오 .

그림 2. FRAP 실험 일시적으로 D의 EGFP-ER로 형질 COS-7 세포를 사용하여. A) 두 OSER 구조 (빨간색의 ROI)을 표백하고, 형광 복구는 시간이 지남에 기록되었다. 명확한 형광 복구는 1 분 후 표백을 감지 할 수있는 신호를 더 증가 4 분 이상 (스케일 바 10 μm의) B). 복구 반 시간을 보여주는 FRAP 실험의 정량 분석 및 개발 EGFP의 이동 비율 -ER 단백질. 망할 CIA큰 그림을 보려면 여기를 ICK

그림 3. 일시적 디 EGFP-ER로 형질 COS-7 세포를 이용한 실험 FLIP. A)로 나타낸 적색 ROI OSER ()의 연속적인 표백을 황색 ROI로 나타낸 ER의 나머지 부분과 동일한 셀 내의 다른 OSER 구조에서 형광의 점진적인 감소 ()시킨다. 노란 화살표는 형광 신호가 시간에 걸쳐 일정되는 표백 셀의 부분을 나타낸다. (스케일 바 10 μm의). B) FLIP 실험의 정량 분석. 큰 그림을 보려면 여기를 클릭하십시오
그림 4. 고정 및 매립 후 OSER 구조물 형광 광학 현미경에 의해 검출 될 수있는 디 EGFP-ER의 높은 수준을 발현하는 세포를 투과 전자 현미경으로 관찰 하였다. 적층 cisternae 및 기복 정현파 세포막 이루어진 OSER 함유 세포의 세포질 부분의 A) 저배율 뷰. 리보솜이 최 cisternae (화살촉과 삽입물)만을 세포막을 장식 반면 미토콘드리아 (M)은, OSER 구조 주위에 클러스터 된 알 수있다. 막 사이에 11 nm 두께의 전자 밀도가 높은 공간은 세포질 (화살표를 삽입) (L = 리소좀 / (자동) 된 phagosomes)과 연속 (스케일 바 1.5 μm의, 삽입 된 0.25 μm의). B) OSER을 형성 할 수있다 연속 또는 fragme 될 수 있습니다 평평 ER의 cisternae의 즉 스택 : 층상 ER로얇은 섹션에있는 자신의 모습에 nted. 스택의 최 cisternae에서 신진 소체 가끔 (별표) (PM, 세포막) (스케일 바을 150 nm)를 관찰 할 수있다.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
이 문서에 설명 된 프로토콜 및 이미징 방법은 살아있는 세포의 ER에 거주 형질 TA 형광 단백질의 분포와 이동을 조사하는 데 사용되었다. 우리는 또한 미세 구조 분석에 의해이 세포 내 구획의 구조에서 이러한 단백질의 과발현의 효과를 분석 하였다.
살아있는 세포 공 촛점 이미징 및 전자 현미경의 조합이 단백질의 동적 특성을 조사의 매우 강력한 수단이다 나타내고, 단백질 기능에 관한 중요한 정보를 제공 할 수있다. 한 방법은 시간 (일반적으로 일 사흘)을 소비하지 않고, 이미지 수집 및 분석을위한 많은 사용자 친화적 인 소프트웨어 응용 프로그램의 개발이 photobleaching에 기반, 라이브 세포 이미징 비교적 간단합니다.
이러한 기술의 주요 제한은 형광 융합 단백질의 사용이다 때문에형광 태그는 그 단백질의 적절한 폴딩 및 / 또는 어셈블리에 영향을 미칠 수있다. 또, 과다 발현은 형질 감염, 형광 태그로 단백질의 동작을 변경할 수 있으므로, 내인성 단백질의 실제 특성을 반영하지 않을 수 있지만, 이는 발현 수준이 될 수있는 유도 안정적 형질 감염된 세포를 사용하여 극복 될 수있다 정확하게 내인성 단백질 1,7의 것과 유사한 수준을 얻기 위해 변조. oligomerise에 FPS의 경향은 광범위하게 문서화되어 상당히 메릭 단백질의 거동 (즉, 반응 속도, 불필요한 단백질 - 단백질 상호 작용과 응집체의 형성)을 변경할 수있다. 최적화 단량체 형광 단백질의 사용은 따라서 17 고려되어야한다.
형광 및 광표백을 사용하여 동적 영상 연구의 또 다른 중요한 측면은 효율적으로 형광 표백제와 형광 재를 측정하기 위해 필요한 시간이다covery (따라서 단백질의 이동성) 정확하게, 또한 투자 수익 (ROI) 및 지방 세포 두께의 영역에 따라 달라집니다. 관련 GFP-태그 된 단백질이 높은 확산 계수를 갖는 경우에, 확산은 표백 동안 발생하기 때문에 복구 시간 측정을 방해 할 수있다. 신속하고 효율적인 표백을 얻기 위해서는, 강한 함수 "줌인"(사용 가능한 경우) 및 하나 이상의 레이저 선을 사용하는 것이 권장된다. 고속 스캔 모듈 (즉 공진 스캐너)의 사용이 크게 실험 회복기 동안 촬상의 속도를 향상시킬 수 있지만, 우리 손에 또한 꽤 표백 효율을 감소시킨다. 그러나, (예컨대 전용 광표백 장치를 구비 회전 디스크와 같은) 다른 스캐닝 시스템, 그리고 더 강력한 레이저 표백 효율 및 수집 속도를 모두 향상시킬 수있다.
FRAP 및 플립 실험에 사용되는 대부분의 형광 단백질은 가역적 광표백 및 BLI의 어느 정도를 표시정량 분석을 수행 할 때 nking이 고려되어야한다. 형광등과 어두운 상태 사이의 변동 분 시간 단위로 두 번째로 발생합니다. EGFP를 들면, 표백 실험 중에, 형광 바리에이션 따라서 본 프로토콜에이 현상을 무시할, 분자의 10 % 미만을 포함 할 수 있음을 보여왔다. 모든 조건이 일정하게 유지되는 경우,이 결과에 일정한 바이어스를 소개 할 것이다. 다른 형광 단백질 리버시블 분획 (즉 YFP) 상당히 높은 경우, 또는 photobleaching에 가역성을 검출하고 평가할 수있는, 사용되는 경우,이 전체를 살아있는 세포에 photobleaching에 후 형광 복구를 측정함으로써 수행 할 수 있으며, 회복이 관찰 된 경우이 수 만 가역성 18 photobleaching에의 결과.
실험 기간 동안 빛의 잠재적 인 독성은 광표백 강한 조명을 필요로 특히 때문에, 또 다른 중요한 요인이다. 그것은 아입니다L 여기 형광체는 다양한 세포 내 과정에서도 세포 생존율 (19)에 영향을 미칠 수있는 자유 라디칼을 생성하도록 산소와 반응하는 경향이 있다고 알려져 있으며, 그래서 효율적인 표백 최소한 광독성 간의 균형을 확립 할 필요가있다, 또한, 세포 생존율은 항상 점검해야 라이브 세포 이미징 실험 후. 짧은 촬영 시간을 감안할 때, 우리는이 문서에 설명 된 예에서 짧은 파장의 빛 (405 NM)의 유전 독성 효과를 고려하지 않았지만, 더 이상 실험이 필요한 경우, 405 nm의 레이저 라인은 사용하지 말아야합니다.
때문에 우리는 OSER 아키텍처의 이기종 자연과 우리는 가능한 한 많은 세포 (구조물)를 관찰하고 싶어한다는 사실을 투과 전자 현미경에 상관 방식을 사용하고 싶지 않아하십니다. 세포에있는 단백질 응집체의 미세 구조의 다양성은 다른 질병의 핵심 기능이 될 수 있으며, 우리는 SA의 넓은 범위를 얻기에 관심이 있었다mples, 상호 접근은 동일 기간의 시간 동안 적은 이벤트의 관측을 허용하는 반면. 쉽게 (예 : 작은 눈에 띄는 ER의 하위로) 또는 (예 : 마이크로 주입 된 세포와 같은) 세포의 제한된 수의에 식별 할 수없는 구조에서 이벤트를 조사 할 때 그러나, 상호 가벼운 전자 현미경 (클레멘 타인)의 첫 번째 선택해야합니다. 그것은 그렇지 noncorrelatively OSER 구조를 관찰 할 가능성이 매우 제한적이며, (세포의 적어도 30 %가 형질 감염시켰다) 우리의 실험은 형질 전환 효율이 높은 것이 특징되었다는 것을 주목할 가치가있다.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
저자가 공개하는 게 없다.
Acknowledgments
저자는이 문서의 게시에있는 그것의 도움과 지원을위한 폰다 치오 Filarete에게 감사의 말씀을 전합니다. 우리는 또한 TECNAI G2 투과 전자 현미경의 사용 센트로 피오 디 Nanomedicina에게 감사의 말씀을 전합니다.
Materials
| Name | Company | Catalog Number | Comments |
| Dulbecco’s Modified Eagle Medium (DMEM) | Invitrogen | 41966029 | |
| Dulbecco’s Modified Eagle Medium (DMEM) w/o phenol red | Invitrogen | 31053028 | |
| Fetal Bovine Serum (FBS) | Invitrogen | 10270106 | |
| Pen/Strep | Invitrogen | 15140-122 | |
| L-Glutamine 200 mM solution | Invitrogen | 25030-024 | |
| jetPEI | Polyplus Transfection | PP10110 | |
| OxyFluor | Oxyrase Inc. | OF-0005 | |
| Glutaraldehyde Grade I | Sigma Aldrich | G5882 | |
| Sodium Cacodylate Trihydrate | Sigma Aldrich | C0250 | |
| Osmium Tetroxide 4% solution | Electron Microscopy Science | 19150 | |
| Uranyl Acetate Dihydrate | Sigma Aldrich | 73943 | slightly radioactive |
| Propylene Oxide | Sigma-Aldrich | 82320 | |
| EPON embedding medium kit | Sigma-Aldrich | 45359-1EA-F | |
| Lead Citrate | Electron Microscopy Science | 17800 | |
| Bench top centrifuge | Eppendorf | 5415 D | |
| Spectral Confocal Microscope | Leica Microsystems | TCS SP5 | |
| CO2 Microscope Cage Incubation System | OkoLab | ||
| Ultramicrotome | Leica Microsystems | UC6 | |
| Diamond knife | Diatome | Ultra 45 ° | |
| Transmission Electron Microscope | FEI | Tecnai G2 | |
| GraphPad Prism Software | GraphPad Software, Inc | ||
| Steel culture cell chamber for 24 mm coverslip | Bioscience Tools | CSC-25 | |
| Electron Microscopy grids | Electron Microscopy Science | G300Cu |
References
- Fasana, E., et al. A VAPB mutant linked to amyotrophic lateral sclerosis generates a novel form of organized smooth endoplasmic reticulum. FASEB J. 24, 1419-1430 (2010).
- Borgese, N., Francolini, M., Snapp, E. Endoplasmic reticulum architecture: structures in flux. Curr. Opin. Cell Biol. 18, 358-364 (2006).
- Ronchi, P., Colombo, S., Francolini, M., Borgese, N. Transmembrane domain-dependent partitioning of membrane proteins within the endoplasmic reticulum. J. Cell Biol. 181, 105-118 (2008).
- Lippincott-Schwartz, J., Snapp, E., Kenworthy, A.
- Lee, M. C., Miller, E. A., Goldberg, J., Orci, L., Schekman, R. Bi-directional protein transport between the ER and. 20, 87-123 (2004).
- Snapp, E. L., et al. Formation of stacked ER cisternae by low affinity protein interactions. J. Cell Biol. 163, 257-269 (2003).
- Papiani, G., et al. Restructured endoplasmic reticulum generated by mutant amyotrophic lateral sclerosis-linked VAPB is cleared by the proteasome. J. Cell Sci. 125, 3601-3611 (2012).
- Almsherqi, Z. A., Kohlwein, S. D., Deng, Y. Cubic membranes: a legend beyond the Flatland* of cell membrane organization. J. Cell Biol. 173, 839-844 (2006).
- Federovitch, C. M., Ron, D., Hampton, R. Y. The dynamic ER: experimental approaches and current questions. Curr. Opin. Cell Biol. 17, 409-414 (2005).
- Takei, K., Mignery, G. A., Mugnaini, E., Sudhof, T. C., De Camilli, P. Inositol 1,4,5-trisphosphate receptor causes formation of ER cisternal stacks in transfected fibroblasts and in cerebellar Purkinje cells. Neuron. 12, 327-342 (1994).
- Feldman, D., Swarm, R. L., Becker, J. Ultrastructural study of rat liver and liver neoplasms after long-term treatment with phenobarbital. Cancer Res. 41, 2151-2162 (1981).
- Sprocati, T., Ronchi, P., Raimondi, A., Francolini, M., Borgese, N. Dynamic and reversible restructuring of the endoplasmic reticulum induced by PDMP in cultured cells. J. Cell Sci. 119, 3249-3260 (2006).
- Costantini, L. M., Fossati, M., Francolini, M., Snapp, E. L. Assessing the tendency of fluorescent proteins to oligomerize under physiologic conditions. Traffic. 13, 643-649 (2012).
- Pyszniak, A. M., Welder, C. A., Takei, F. Cell surface distribution of high-avidity LFA-1 detected by soluble ICAM-1-coated microspheres. J. Immunol. 152, 5241-5249 (1994).
- Taylor, J. P., Hardy, J., Fischbeck, K. H.
- Winklhofer, K. F., Tatzelt, J., Haass, C. The two faces of protein misfolding: gain- and loss-of-function in neurodegenerative diseases. EMBO J. 27, 336-349 (2008).
- Borgese, N., Gazzoni, I., Barberi, M., Colombo, S., Pedrazzini, E. Targeting of a tail-anchored protein to endoplasmic reticulum and mitochondrial outer membrane by independent but competing pathways. Mol. Biol. Cell. 12, 2482-2496 (2001).
- Chudakov, D. M., Matz, M. V., Lukyanov, S., Lukyanov, K. A. Fluorescent proteins and their applications in imaging living cells and tissues. Physiol Rev. 90, 1103-1163 (2010).
- Bancaud, A., Huet, S., Rabut, G., Ellenberg, J. Fluorescence perturbation techniques to study mobility and molecular dynamics of proteins in live cells FRAP, photoactivation, photoconversion, and FLIP. Cold Spring Harb. Protoc. 2010, (2010).
- Michida, T., et al. Role of endothelin 1 in hemorrhagic shock-induced gastric mucosal injury in rats. Gastroenterology. 106, 988-993 (1994).

{kind=link}