

Summary
我们描述了使用调查转染荧光蛋白通过活细胞的共聚焦成像装置驻留在内质网(ER)的分布和流动的成像方法。我们还超微结构分析其表达对这个亚细胞区室的结构的效果。
Abstract
在真核细胞中的脂质和蛋白质细胞区室之间不断交换,但这些保留其独特的组成和功能,尽管激烈的interorganelle分子的流量。在本文中描述的技术是强大的研究蛋白质和脂质的流动性和贩运体内 ,并在其生理环境的手段。光漂白(FRAP)和荧光损失在光漂白(FLIP)后荧光恢复被广泛用于活细胞成像技术通过外 - 内吞途径,细胞器或子隔离之间的连续性,形成蛋白质复合物,以及蛋白质定位研究细胞内运输所有这些脂质微区,可生理和病理条件下观察。这些方法的局限性主要是由于使用了荧光融合蛋白,以及它们的潜在缺点包括伪迹过表达离子在细胞内和在标记的和天然蛋白质的折叠和定位的差异的可能性。最后,由于光学显微镜(约200纳米)的分辨率的极限不允许急诊室或特定的小班,可以在细胞应激状态下起源的精细结构的研究( 即缺氧,药品监督管理,过度表达的跨膜ER常驻蛋白)或病理条件下,我们结合培养的转染细胞的活细胞成像与超微结构分析了基于透射电子显微镜。
Introduction
绿色荧光蛋白(GFP)及其变种光谱和荧光显微镜的并行开发的发现,开辟了全新的途径,在细胞中蛋白质的行为进行调查。技术,如荧光漂白恢复(FRAP)和荧光损失漂白(FLIP),这是因为荧光的内在能力,以扑灭在强光照射下其荧光的可能后,基于共聚焦活细胞成像和使用的转荧光融合蛋白1-3。它们被广泛用于评估蛋白质不仅是本地化,而 且他们的流动性和膜泡运输,它可以揭示有关它们的功能4重要线索。
真核细胞的独特之处在于具有特定的脂质和蛋白组成的细胞内隔室的存在。虽然细胞器是物理隔离器版,它们需要彼此通信,并以维持细胞稳态交流分子成分。分泌途径保证了在ER中合成的蛋白质和脂质到达正确的最终目标,其中它们发挥它们的功能。细胞内细胞器,也可以通过动态接触部位,使分子(脂质)向隔间之间直接交换连接。此外,许多蛋白已到组装在大异聚复合物,或与特定的脂类物质(脂质筏/微区)为了成为功能活性或者被运送到最终目的地相关联。所有这些生物方面极大地影响蛋白质的动力学性质,并因此可以通过如下所述的技术手段进行适当的调查。
我们的团队已经广泛应用于FRAP和FLIP结合电子显微镜来研究内质网的结构和它的不同的子域。内质网是分泌途径的第一站和在蛋白质和脂质排序5起关键作用。这是一个高度动态的细胞器,其独特的子域名反映其许多不同的功能( 如蛋白质和脂质的合成和贩运,蛋白质折叠,Ca 2 +的储存和释放,以及异物代谢)。然而,尽管它们在形态上,空间上,和功能上不同,这些结构域彼此连续,并且它们的相对丰度可在细胞生理和病理条件下进行修改。最有名的和内质网中,通常在空间上分离的结构域是核膜和平滑和粗糙的ER,但是,我们和其他人已经证明,有ER结构具有更复杂的结构和三维组织在各种细胞类型和在生理条件下的组织,也可以由紧张刺激如缺氧,药物诱导的手段管理或过度表达的ER驻留跨膜蛋白2,6(以及其中的参考文献)。
我们最近还展示了这样的结构在人类疾病中1,7的细胞模型的存在。从滑面内质的堆积潴发起,他们分别获得组织光面内质网(OSER)的总称,2003年6,虽然他们也被称为karmellae,片状结晶 ,并且晶体ER他们的架构的基础上,像他们的大小,可以改变。之后,细胞转染绿色荧光蛋白融合到尾锚(TA)ER居民蛋白质(D EGFP-ER),绿色荧光蛋白在转弱二聚化倾向显着地改变了ER的组织和结构的细胞质区域。 FRAP和FLIP实验表明使得 d EGFP-ER是自由OSERs内扩散,并且它从网状ER移动到OSER 反之亦然<事实/ em>的指示聚集是连续的与周围的网状急诊室。超微结构分析使我们能够将荧光数据与OSER体系结构和组织在纳米尺度水平上的详细描述关联:OSERs总是由成堆的光滑ER配对囊泡的,但可能有不同形式的空间组织,如规则排列的正弦数组或轮生,或六角形“晶体”管阵列。这些重排导致立方形态8的,因为它们在细胞中发现的生理条件9和下面的压力,如缺氧10,药物治疗11和癌症9下,可有显 著电位超微结构标记。
使用GFP融合蛋白第一次演示之后,我们使用了成像实验,分析ER结构域的增殖响应于药物治疗12,ASSESS荧光蛋白的倾向oligomerise在细胞13,并调查的突变,ALS联TA蛋白在ER起源的细胞内聚集,可能是有关其致病1,8的形成中的作用。它已被提出,形成的细胞内聚集物(这发生在许多神经退行性疾病14)可以设计成防止有毒的突变蛋白和周围的细胞成分15之间的相互作用的一种保护机制。
以下是对光学和电子显微镜方法考察构建体的C-末端的疏水性结构域被插入到内质网的膜,并且其动态行为的分析以及其过度表达对ER域的效应的组合的说明在培养的细胞结构(参见图1的实验方案的流程图)。
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1。质粒,细胞培养,转染,并与ER荧光蛋白
- 在本研究中使用的质粒由GFP的增强版本融合在其C-末端的尾部区域的大鼠色素b的ER亚型(5)通过接头序列(如B(5)此处略)。尾部区域包含整个序列(Pro94-Asp134),该保持天然B的胰蛋白酶切割位(5),其中包括17个残基的TMD(跨膜结构域),由上游和下游的极性序列侧接(UPS和DPS)后膜相关。连接器包括myc表后跟[(甘氨酸)4辑] 3,并且整个的cDNA被插入到哺乳动物表达载体pcDNA3的Hind3-Xba1位点。该质粒的构造细节在以前的出版物中,它被称为GFP-ER 16已被描述。
- 生长的COS-7细胞在Dulbecco改进的Eagle培养基(DMEM),补充有10%羊升牛血清,2mM L-谷氨酰胺,1%青霉素/链霉素的培养箱中,在37℃和10%的CO 2。
- 转染。板3×10 5个细胞在6孔板中的圆形玻璃盖玻片,并在次日,如制造商所述转染与JETPEI系统。注意,最优JETPEI / DNA比率,以建立根据所使用的质粒和细胞系的最大转染效率进行了测试:在我们的情况下,JETPEI:2:1的DNA比导致70-80%的转染效率。
2。实时荧光扫描共聚焦显微镜
- 活细胞成像。把在其上的转染细胞接种到一个钢细胞培养室,用于24毫米盖玻片填充用DMEM W / O型酚红,补充有10%FBS,2mM L-谷氨酰胺,1%青霉素/链霉素,25mM的HEPES盖玻片,50微克/毫升放线菌酮和1:100 OxyFluor以防止样品从光漂白。一个SP5共焦microscopë配备有温度控制的CO 2培养箱(37℃和5%CO 2)用于活细胞成像的实验中,使用488nm的激光和一个五十分之五百二十五带通发射过滤器D的GFP-ER被可视化。
- 漂白(FRAP)后荧光恢复。绘制感兴趣区(ROI)的区域,对应于OSER结构,它采用20次迭代和488 nm的组合(30毫瓦氩激光的100%,相当于5.5-6μW在样品)和405 nm的漂白(60%的一个30毫瓦二极管405的激光,对应于11.6μW在样品)激光器,在我们的经验,导致了高效率和快速漂白。
- 通过采取一个单一的框架每10秒,10分钟(像素时间= 1.61微秒/像素)记录荧光的复苏漂白的投资回报。
- 荧光损失漂白(FLIP)。绘制对应于一OSER结构的投资回报率,并且如上所述的漂白剂。漂白重复每30秒,和后漂白图像被记录,每10秒进行30分钟(像素时间= 1.61微秒/像素)。
- FRAP和FLIP分析。所有的图像都使用ImageJ软件(分析http://rsbweb.nih.gov/ij/download.html )。在FRAP实验中,漂白ROI的荧光恢复测量随着时间的推移和归一化到漂白细胞的总荧光,它总是检查,以随时间恒定。
- 对于FLIP实验中,得出一个投资回报率的漂白OSER和覆盖整个细胞外。测量其荧光强度随着时间的推移和正常化到ROI的荧光水平,以纠正在荧光任何减少所造成的成像本身上绘制一个未漂白细胞。
- 在所有的实验中,从感兴趣区域的荧光强度中减去背景信号(在细胞外的区域确定)。最后,使用石墨绘制的结果aphPad Prism软件。
3。超微结构分析透射电子显微镜的手段
鉴于许多试剂的毒性,所有的程序都要进行穿着适当的实验室。大衣和手套在通风橱内。
- 从培养皿中取出盖玻片后,固定生长在培养皿如使用过滤的2%戊二醛的0.1M二甲胂酸盐缓冲液,pH 7.4的单层的底部,在室温下10分钟,剩余的细胞。
- 采用聚四氟乙烯刮板刮去细胞,并将它们转移到1.5ml离心管中。通过离心的方法使细胞沉淀以9,000 g下10分钟。除去上清液,加入新鲜固定液,并留下过夜,4°C。
- 清洗用缓冲液的颗粒,然后用1%四氧化锇在二甲胂酸盐缓冲液1小时,在室温下的溶液中后固定。
- 冲洗用超纯水,以及整块污渍1%乙酸双氧铀在蒸馏水中20-60分钟之间。
- 脱水的样品中增加的乙醇系列(70%,80%,90%,100%,100%,每次10分钟),并在两次环氧丙烷简要地洗涤(每次15分钟)。
- 渗入样品中的氧化丙烯的Epon +(1:1)的混合物(从2小时到过夜)。
- 嵌入的Epon环氧树脂固化物在60℃下至少24小时。
- 部分使用超薄切片机LEICA UC6配备有45°的金刚石刀,以获得区段的厚度为60-70纳米的手动修整树脂集团。收集300目铜网上的部分。
- 染色对电网的部分用醋酸铀(20分钟)的饱和溶液和柠檬酸铅(7分钟),通过浸在双蒸过滤水彻底清洗网格,并允许他们在室温下干燥。
- 染色的网格使用的是TECNAI G2的透射电子显微镜观察和图像是使用博特捕获OM式CCD摄像头在不同的最终放大倍率(一般为6,000-39,000倍)。
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
图2显示了蛋白质的流动性的一个例子FRAP研究。 ðEGFP-ER蛋白的迁移率证明了在漂白漂白OSERs后迅速荧光恢复。对于定量分析,在中场休息时和移动部分均来自实验测得的数据进行拟合以下单指数方程:
F(T)= F + 后 (F REC-F后 )(1-E-T /τ)
其中F是后荧光信号漂白后,F REC是漂白后到达,T登记时间和τ时间常数的最大荧光回收价值。
请注意,获取图像不饱和的像素,可以改变荧光的恢复,因此,该蛋白质迁移率分析的重要性。据我S也必须始终正常化的漂白ROI的荧光信号,以相同的细胞,以便考虑图像采集或小的变化在聚焦平面时,由于漂白荧光强度变化的总荧光。
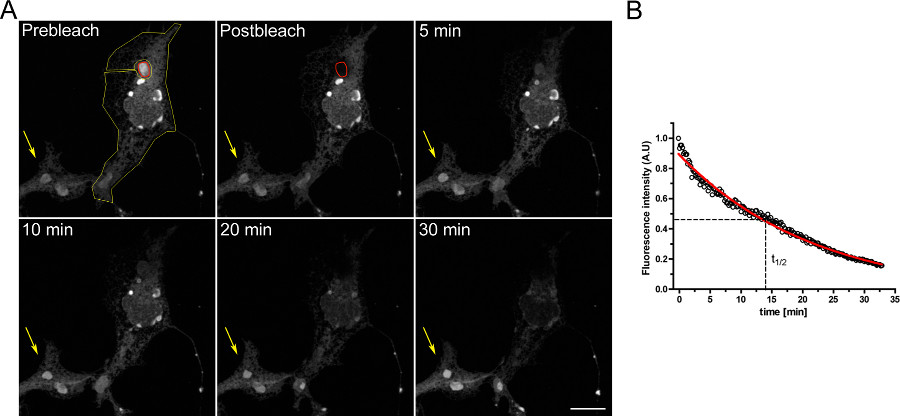
倒装实验来研究细胞内隔室之间的连续性的一个例子示于图3。 OSERs物理与ER的其余部分连接就证明了当OSER领域不断漂白急诊室的逐渐排空。
一个适当的分析,必须避免采集饱和像素(见上文),此外,采集参数必须设置与激光功率尽可能地低,以避免光漂白由于图像采集。出于这个原因,强烈建议到图像中的同一种漂白细胞将被用于归一化的漂白C的荧光信号得寸进尺。
所有的实验都必须在放线菌酮,翻译抑制剂的存在下进行,以避免在ER中的荧光信号的任何增加(并因此总荧光)由于蛋白质的生物合成。
透射电子显微镜显示出,在转染以d EGFP-ER的培养的细胞中观察到的荧光聚集体代表的光滑补丁和扁平空间上自行组织分类的模式的基础上,定义良好的三维几何形状的ER囊泡:线性的或弯曲的叠层(常与核膜,未示出)( 图4A和B),其可以是连续的正弦ER的区域( 图4A)相关联,在某些区域中的细胞膜被组织成格子具有正方形或六边形对称性(晶体ER,图中未示)。相邻的囊泡是由一层薄的分离略微电子基茂密的细胞质约11纳米厚是连续的周围聚集细胞质中。

图1。流程图实验过程中,将培养的细胞,以过表达所关心的荧光融合蛋白先转jetPEI转染(见协议)。 24小时后,活转染的细胞可视化和FRAP和FLIP实验使用共聚焦显微镜配有控制温度的CO 2培养箱中进行,并且所记录的图像中导出,并用适当的软件( 例如 ImageJ的)分析。用于超微结构分析,转染的细胞被固定,沉淀,并嵌入在EPON环氧树脂集团。超薄切片使用钻石刀获得,收集的关于c奥珀网格,并在透射电子显微镜下观察。 点击这里查看大图 。

图2。 FRAP实验使用COS-7细胞中瞬时转染以d EGFP-ER。 a)两个OSER结构(红色的ROI)进行漂白和荧光恢复,录得超过时间。清晰的荧光恢复,可以检测1分钟后的脱色,且信号进一步增大4分钟后(比例尺为10μm)B):在FRAP实验示出了恢复半时间的定量分析与d EGFP的移动部分-ER蛋白。 氯ICK这里查看大图

图3。 FLIP实验使用COS-7细胞中瞬时转染以d EGFP-ER。 A)一种OSER(由红色ROI表示)的连续漂白导致逐步减少在荧光在ER中的其余部分,并在同一小区内的其他OSER结构(由黄色ROI表示)。黄色箭头表示漂白电池的一部分,其中的荧光信号随时间保持恒定。 (比例尺10微米)。 二)FLIP实验的定量分析。 点击这里查看大图
图4。固定和包埋后,将细胞表达高水平ðEGFP-ER,其中OSER结构可以由荧光显微镜的方法进行检测,通过透射型电子显微镜进行观察。包含OSER由堆叠囊泡和起伏的正弦膜细胞的细胞质的一部分A)低倍率看法。线粒体(M)可以看到周围的OSER结构聚集,而核糖体装饰最外层的囊泡(箭头和插图),只有膜。膜之间的11纳米厚的电子密度的空间是连续的细胞质(箭头和插图)(L =溶酶体/(自动)吞噬体)(比例尺为1.5μm;插页0.25微米)。 乙 )一个OSER可以形成由层状ER: 即扁平ER潴栈,可连续或fragme与指向其在薄片的外观。囊泡从堆栈的最外囊泡出芽偶尔可以观察到(星号)(PM,质膜)(比例尺为150nm)。
Subscription Required. Please recommend JoVE to your librarian.
Discussion
本文所描述的协议和成像方法已被用于研究转助教荧光蛋白居民在活细胞的内质网的分布和流动。我们也分析了这个亚细胞区室中通过超微结构分析装置的体系结构中的过度表达,这些蛋白质的效果。
活细胞共聚焦成像和电子显微镜的组合代表是一个非常强大的研究蛋白质的动态性能的手段,并可能提供有关蛋白质功能的重要信息。所描述的方法不是费时(一般工作三天),以及许多用户友好的软件应用程序的开发,用于图像采集和分析,使漂白为主,活细胞成像比较简单。
这些技术的主要限制是在使用荧光融合蛋白,因为荧光标记可以影响正确折叠所感兴趣的蛋白和/或组件。此外,过表达可改变转染,荧光标记的蛋白质的行为,并且因此可能不反映内源蛋白质的真实属性,但是,这可以通过使用可诱导的和稳定转染的细胞中的表达水平可以是可以克服精确调制,以获得与内源性蛋白1,7水平相当。 FP的给oligomerise的倾向已被广泛记载并且可能显著改变嵌合蛋白的行为( 即动力学,不需要的蛋白-蛋白相互作用,并形成聚集体)。采用优化的单体荧光蛋白,因此应考虑17。
利用荧光和光漂白动态成像研究的另一个重要方面是有效地漂白的荧光,测量荧光重所需的时间covery(并因此蛋白质迁移率)精确地说,它也依赖于ROI和本地单元厚度的区域。如果一个给定的GFP标记的蛋白质具有高的扩散系数,可能会发生在漂白过程中的扩散,从而干扰恢复时间的测量。为了获得快速,高效漂白,强烈建议功能的“放大”(如果有的话)和一个以上的激光线使用。虽然采用了快速扫描模块( 即共振扫描器)的可以在实验的恢复阶段大大提高成像的速度,拿在手上也大大降低漂白效率。然而,另一种扫描系统(如配备了专用的漂白装置一个旋转的圆盘),和更强大的激光器能同时提高漂白效率和采集速度。
在大多数FRAP和FLIP实验中使用的荧光蛋白表现出一定程度的可逆性漂白和劳保局nking进行定量分析时必须考虑的。荧光和暗态之间的波动发生在第二至分钟的时间尺度。对于EGFP,它已经表明,在漂白实验,荧光的变化可能涉及的分子的10%以内,因此本协议中这种现象可以忽略不计。如果所有的条件都保持不变,这将在结果引入一个恒定偏压。如果其它荧光蛋白的使用,其中所述可逆分数是显著更高( 即 YFP),或检测和评估漂白可逆性,这可以通过测量荧光漂白恢复在整个活细胞之后进行,如果恢复是观察这可以仅是光漂白可逆性18的结果。
光在实验过程中的潜在毒性是另一个关键因素,特别是因为漂白需要强大的照明。这是WEL升已知激发的荧光团易与氧发生反应,产生自由基,可影响多种细胞内过程,甚至细胞活力19,因此,有必要建立有效的漂白和最小的光毒性之间的平衡,此外,细胞存活率应始终检查后活细胞成像实验。由于录制时间短,我们没有考虑短波长光(405纳米)的遗传毒性效应在本文中所描述的例子,但是,如果需要一个较长的实验,不应该使用一个405 nm的激光线。
我们选择不使用,因为OSER架构的异构性,但我们希望观察尽可能多的细胞(和结构)尽可能的其实是一个相关的方法来透射电子显微镜。蛋白质聚集在细胞的精细结构的多样性可能是不同疾病的主要特征,我们有兴趣获得广泛的samples,而相关的方法可以让更少的事件在同一时间内观察。然而,相关的光致电子显微镜(CLEM)应该调查,不能容易地确定(例如,较不突出的ER子域),或在细胞中的有限数量的(如微注射的细胞)中的结构的事件时是第一选择。值得注意的是,我们的实验,其特征在于,转染效率高的程度(在细胞中的至少30%的转染),否则观察OSER结构noncorrelatively的可能性是非常有限的。
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
作者什么都没有透露。
Acknowledgments
作者感谢基金会Filarete提供的帮助和支持,在此文章的发表。我们还要感谢炫酷那么Europeo二Nanomedicina为使用该Tecnai G2透射电子显微镜。
Materials
| Name | Company | Catalog Number | Comments |
| Dulbecco’s Modified Eagle Medium (DMEM) | Invitrogen | 41966029 | |
| Dulbecco’s Modified Eagle Medium (DMEM) w/o phenol red | Invitrogen | 31053028 | |
| Fetal Bovine Serum (FBS) | Invitrogen | 10270106 | |
| Pen/Strep | Invitrogen | 15140-122 | |
| L-Glutamine 200 mM solution | Invitrogen | 25030-024 | |
| jetPEI | Polyplus Transfection | PP10110 | |
| OxyFluor | Oxyrase Inc. | OF-0005 | |
| Glutaraldehyde Grade I | Sigma Aldrich | G5882 | |
| Sodium Cacodylate Trihydrate | Sigma Aldrich | C0250 | |
| Osmium Tetroxide 4% solution | Electron Microscopy Science | 19150 | |
| Uranyl Acetate Dihydrate | Sigma Aldrich | 73943 | slightly radioactive |
| Propylene Oxide | Sigma-Aldrich | 82320 | |
| EPON embedding medium kit | Sigma-Aldrich | 45359-1EA-F | |
| Lead Citrate | Electron Microscopy Science | 17800 | |
| Bench top centrifuge | Eppendorf | 5415 D | |
| Spectral Confocal Microscope | Leica Microsystems | TCS SP5 | |
| CO2 Microscope Cage Incubation System | OkoLab | ||
| Ultramicrotome | Leica Microsystems | UC6 | |
| Diamond knife | Diatome | Ultra 45 ° | |
| Transmission Electron Microscope | FEI | Tecnai G2 | |
| GraphPad Prism Software | GraphPad Software, Inc | ||
| Steel culture cell chamber for 24 mm coverslip | Bioscience Tools | CSC-25 | |
| Electron Microscopy grids | Electron Microscopy Science | G300Cu |
References
- Fasana, E., et al. A VAPB mutant linked to amyotrophic lateral sclerosis generates a novel form of organized smooth endoplasmic reticulum. FASEB J. 24, 1419-1430 (2010).
- Borgese, N., Francolini, M., Snapp, E. Endoplasmic reticulum architecture: structures in flux. Curr. Opin. Cell Biol. 18, 358-364 (2006).
- Ronchi, P., Colombo, S., Francolini, M., Borgese, N. Transmembrane domain-dependent partitioning of membrane proteins within the endoplasmic reticulum. J. Cell Biol. 181, 105-118 (2008).
- Lippincott-Schwartz, J., Snapp, E., Kenworthy, A. Studying protein dynamics in living cells. Nat. Rev. Mol. Cell Biol. 2, 444-456 (2001).
- Lee, M. C., Miller, E. A., Goldberg, J., Orci, L., Schekman, R. Bi-directional protein transport between the ER and. 20, 87-123 (2004).
- Snapp, E. L., et al. Formation of stacked ER cisternae by low affinity protein interactions. J. Cell Biol. 163, 257-269 (2003).
- Papiani, G., et al. Restructured endoplasmic reticulum generated by mutant amyotrophic lateral sclerosis-linked VAPB is cleared by the proteasome. J. Cell Sci. 125, 3601-3611 (2012).
- Almsherqi, Z. A., Kohlwein, S. D., Deng, Y. Cubic membranes: a legend beyond the Flatland* of cell membrane organization. J. Cell Biol. 173, 839-844 (2006).
- Federovitch, C. M., Ron, D., Hampton, R. Y. The dynamic ER: experimental approaches and current questions. Curr. Opin. Cell Biol. 17, 409-414 (2005).
- Takei, K., Mignery, G. A., Mugnaini, E., Sudhof, T. C., De Camilli, P. Inositol 1,4,5-trisphosphate receptor causes formation of ER cisternal stacks in transfected fibroblasts and in cerebellar Purkinje cells. Neuron. 12, 327-342 (1994).
- Feldman, D., Swarm, R. L., Becker, J. Ultrastructural study of rat liver and liver neoplasms after long-term treatment with phenobarbital. Cancer Res. 41, 2151-2162 (1981).
- Sprocati, T., Ronchi, P., Raimondi, A., Francolini, M., Borgese, N. Dynamic and reversible restructuring of the endoplasmic reticulum induced by PDMP in cultured cells. J. Cell Sci. 119, 3249-3260 (2006).
- Costantini, L. M., Fossati, M., Francolini, M., Snapp, E. L. Assessing the tendency of fluorescent proteins to oligomerize under physiologic conditions. Traffic. 13, 643-649 (2012).
- Pyszniak, A. M., Welder, C. A., Takei, F. Cell surface distribution of high-avidity LFA-1 detected by soluble ICAM-1-coated microspheres. J. Immunol. 152, 5241-5249 (1994).
- Taylor, J. P., Hardy, J., Fischbeck, K. H. Toxic proteins in neurodegenerative disease. Science. 296, 1991-1995 (2002).
- Winklhofer, K. F., Tatzelt, J., Haass, C. The two faces of protein misfolding: gain- and loss-of-function in neurodegenerative diseases. EMBO J. 27, 336-349 (2008).
- Borgese, N., Gazzoni, I., Barberi, M., Colombo, S., Pedrazzini, E. Targeting of a tail-anchored protein to endoplasmic reticulum and mitochondrial outer membrane by independent but competing pathways. Mol. Biol. Cell. 12, 2482-2496 (2001).
- Chudakov, D. M., Matz, M. V., Lukyanov, S., Lukyanov, K. A. Fluorescent proteins and their applications in imaging living cells and tissues. Physiol Rev. 90, 1103-1163 (2010).
- Bancaud, A., Huet, S., Rabut, G., Ellenberg, J. Fluorescence perturbation techniques to study mobility and molecular dynamics of proteins in live cells FRAP, photoactivation, photoconversion, and FLIP. Cold Spring Harb. Protoc. 2010, (2010).
- Michida, T., et al. Role of endothelin 1 in hemorrhagic shock-induced gastric mucosal injury in rats. Gastroenterology. 106, 988-993 (1994).

{kind=link}