We first tested if the antibodies did not interfere with the barrier function of the endothelium. We measured the resistance of endothelial monolayers by using electrical cell-substrate impedance sensing (ECIS). For details, see Van Buul et al.13 No change in resistance was observed when the anti-VE-cadherin fluorescently-labeled antibody was added to the cells (Figure 2A). An anti-VE-cadherin antibody that is well-recognized to block VE-cadherin function reduced the resistance dramatically (Figure 2A). Also, the antibodies used for imaging do not alter the dynamics of VE-cadherin, as was assessed by measuring fluorescent recovery after photo-bleaching (FRAP) of VE-cadherin-GFP (Figure 2B).
After 1-2 min, neutrophils adhered to the activated endothelial monolayer as could be visualized in the DIC channel (Figure 3A). After crawling for 5-30 sec, the vast majority of neutrophils transmigrated through the endothelial monolayer through the cell-cell junctions that were labeled with the antibodies directed against PECAM-1 and VE-cadherin. During the process of diapedesis, the distribution of VE-cadherin and PECAM-1 was followed in real-time (Figure 3B). At sites of neutrophil diapedesis, VE-cadherin was locally disrupted and PECAM-1 showed a more ring-like structure. After completion of diapedesis, the junctions close and VE-cadherin and PECAM-1 clearly relocated at the sites of diapedesis (Figure 3C). Note that parts of the anti-PECAM-1 antibody were detected on the surface of the neutrophil once the neutrophil reached the baso-lateral side of the endothelium. However, this did not prevent neutrophils from crossing the endothelium. The observed dynamics of VE-cadherin in real-time during neutrophil transendothelial migration were in agreement with the work by Shaw and colleagues who showed, using confocal microscopy and VE-cadherin-GFP-transfected endothelial cells, that VE-cadherin-GFP diffused laterally when leukocytes crossed the endothelial cell-cell junctions14. Also work by Su and coworkers underscored our observations of PECAM-1. They showed that, next to lateral VE-cadherin diffusion upon leukocyte passage, PECAM-1 was locally redistributed into a ring around the transmigrating neutrophil15.

Figure 1. In vitro flow chamber. (A) Arrow indicates the reservoir from which the medium needs to be refreshed. (B) Silicone tubing that connects the flow chamber with the in-line Luer injection port used for injecting PMNs without disconnecting the tubing (arrowhead). (C) Connection of an empty flow-chamber to the reservoir flask filled with 37 °C flow-buffer (arrowhead). (D) Connection of the flow-chamber with the TNF-α-treated HUVECs to the flow-buffer-containing tubing and placed in the microscope stage (arrowhead).

Figure 2. VE-cadherin antibodies do not interfere with junctional dynamics or function. (A) Endothelial cell monolayer impedance is measured using ECIS. Y-axis expresses impedance in Ohms and x-axis represents time in hours. VE-cadherin antibody clone 55-7H1, labeled with ALEXA647 (blue line) or isotype IgG-ALEXA647 control (red line) do not alter endothelial cell monolayer impedance, whereas the VE-cadherin blocking antibody cl75 (black line) did reduce the impedance. (B) VE-cadherin-GFP was expressed in HUVEC and FRAP analysis using confocal microscopy revealed no change in fluorescence recovery in the presence or absence of the VE-cadherin 55-7H1 antibody.
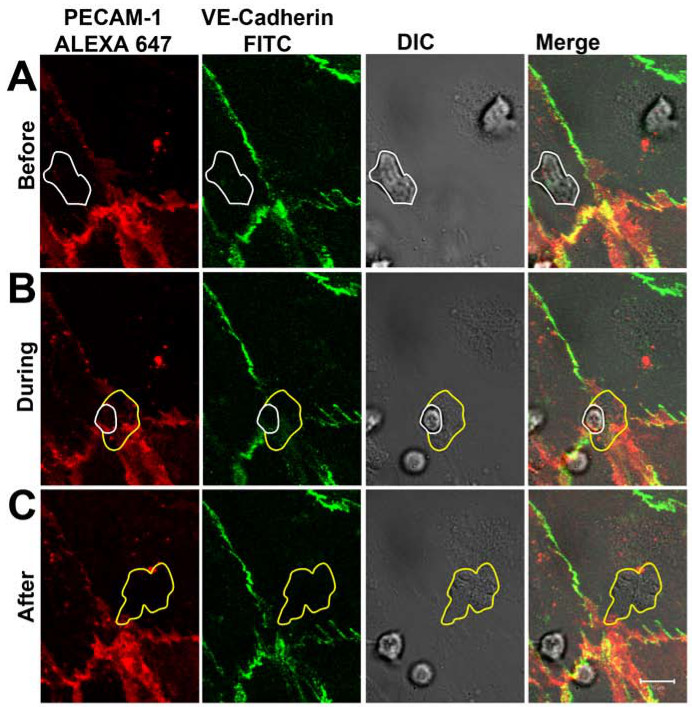
Figure 3. VE-cadherin and PECAM-1 distribution during neutrophil TEM in real-time. (A) Neutrophil (marked with white line) adhering on the endothelium and crossing the cell-cell junction without affecting the distribution of VE-cadherin and PECAM-1. (B) Neutrophil crossing the endothelial monolayer through the cell-cell junctions. A local dispersion of VE-cadherin and PECAM-1 can be observed when a neutrophil protrudes through the cell-cell junctions. White line illustrates neutrophil presence still on top of the endothelium. Yellow line shows neutrophil membrane that is already underneath endothelium. (C) Neutrophil has fully crossed the endothelial monolayer. VE-cadherin and PECAM-1 are relocated at sites of diapedesis. Yellow line illustrates borders of the transmigrated neutrophil. Please click here to view a larger version of this figure.
