Measurement of parasitemia.
For the measurement of parasitemia, blood cells should first be selected, and noise, debris and platelets excluded, based on FSC/SSC properties (Figure 2A). Depending on the cytometer used, single cells should then be selected based on either trigger pulse width (Figure 2B), or FSC peak height to area ratio (Figure 2C). Remaining events should consist of leukocytes, stained positive for APC eFluor 780, RBC progenitors (including reticulocytes), stained positive for PerCP eFluor 710, and mature RBCs, negative for both these stains (Figure 2D). After selecting the mature RBC population, events should separate into four populations when JC-1 fluorescence is plotted against Hoechst 33342 (Figure 2E,F) or Hoechst 34580 (Figure 2G,H) fluorescence. The dual positive population represents parasitized RBCs, while the remaining populations are either uninfected RBCs (dual negative), HJ-RBCs (Hoechst positive, JC-1 negative), or unknown cells (JC-1 positive, Hoechst negative). Notably, JC-1 fluorescence may shift from a maximum at 529 nm to a broad emission peaking at 590 nm, depending on the mitochondrial membrane potential of the cell. Regardless of this shift JC-1 will fluoresce strongly in the 530/40 nm channel, making this channel most suitable for analysis. However, in some instances it may be worthwhile to additionally record fluorescence in the 585/42 nm channel, which may provide an indication of the mitochondrial membrane potential of cells. In uninfected samples less than 0.007% of events should be dual positive (Figure 2E,G). This result will depend on the cleanliness of the flow cytometer and may vary considerably, extensive cleaning before analysis may improve results. HJ-RBCs account for 0.3 – 0.9% of mature RBCs, making the addition of the JC-1 dye critical for the accurate measurement of low parasitemia.
Assessment of merozoite invasion.
After injecting labeled RBCs into infected mice the relative invasion rates into the two labeled populations can be determined. Blood samples should be taken and prepared as described for the parasitemia measurement with the addition of Streptavidin PE Cy7. The time at which the sample is taken depends on the experimental conditions and desired outcome of the analysis. In general, an earlier time point will best reflect an invasion phenotype, although it may be beneficial to collect multiple time points to increase accuracy. For the analysis of flow cytometry data, mature RBCs should be selected as in Figure 2. From these cells, Atto 633 and biotin labeled RBCs can be identified based on fluorescence in the 670/30 and 750LP channels respectively (Figure 3A). From here on the three RBC populations (Atto 633, biotin, and unlabeled) should be analyzed separately. In each of these populations parasitemia can be determined based on staining with Hoechst and JC-1 (Figure 3B-D).

Figure 1. Schematic of the in vivo parasite invasion assay. An example of how this assay can be used to determine relative invasion rates in RBCs treated according to the user’s biological question is shown. Blood collected from uninfected mice is divided into two tubes. One tube is treated as desired while the other sample is left untreated (A). Tubes are again divided with one labeled with NHS-Biotin and the other with NHS-Atto 633 (B). Samples are then combined in two combinations; Biotin labeled treated RBCs with Atto 633 labeled untreated RBCs and Atto 633 labeled treated RBCs with Biotin labeled untreated RBCs (C). These two combinations are injected separately into two lots of infected mice during schizogony at 2 – 15% parasitemia (D). A total of 6 recipient mice is recommended to gain statistical significance in the result, although more or less may be used if needed. Adapted from Lelliott et al.14, originally published by BioMed Central.
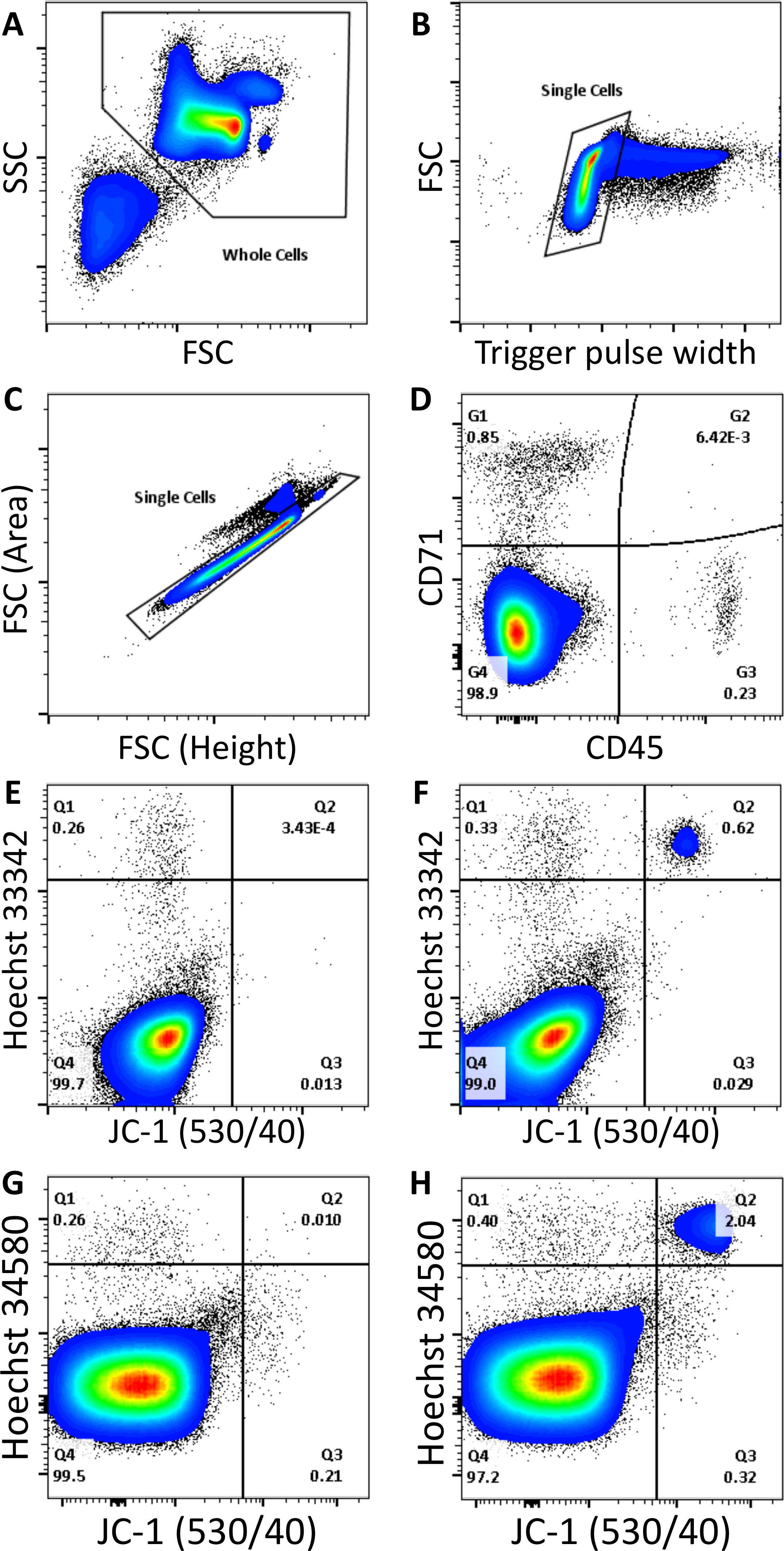
Figure 2. Measurement of parasitemia by flow cytometry. Debris, noise, and platelets are removed from analysis based on FSC/SSC properties (A). Single cells are then selected from the remaining cells based on either trigger pulse width (B), or FSC peak to area ratio (C). Cell types can then be distinguished based on positive staining with CD45 APC eFluor 780 (leukocytes), CD71 PerCP eFluor 710 (RBC progenitors), or negative staining (mature RBCs) (D). After selecting mature RBCs, cells are either: Hoechst and JC-1 positive (parasitized RBCs), Hoechst positive and JC-1 negative (RBCs containing Howell Jolly bodies), JC-1 positive and Hoechst negative (unknown cells), or dual negative (uninfected RBCs). Uninfected and P. chabaudi infected samples stained with Hoechst 33342 (E,F), or Hoechst 34580 (G,H), are shown. Please click here to view a larger version of this figure.

Figure 3. Assessment of merozoite invasion into two labeled RBC populations in vivo. Plots are of mature RBCs, gated as in Figure 2. The two labeled RBC populations can be selected based on their label (A). Atto 633 labeled cells fluoresce in the 670/30 channel (L1), biotin labeled cells bind to streptavidin and fluoresce in the PE-Cy7 channel (L3), and unlabeled cells are negative for these stains (L4). In each of these populations parasitized RBCs can be identified based on Hoechst and JC-1 positive staining (Q2) (B-D). Please click here to view a larger version of this figure.
