

ERRATUM NOTICE
Important: There has been an erratum issued for this article. Read more …
Summary
Vi presenterer en protokoll for klassisk betinging av utnyttet maur som tillater forskere å studere visuelle læring og hukommelse formasjon på et analyse ikke mulig med fritt flytte personer.
Abstract
Flere arter av insekter har blitt modellsystemer for å studere læring og hukommelse formasjon. Selv om mange studier fokuserer på fritt flytte dyr, studier implementere klassisk betinging paradigmer med utnyttet insekter har vært viktig for å undersøke nøyaktig signaler som enkeltpersoner lære og nevrale mekanismene bak læring og minne formasjon. Her presenterer vi en protokoll for fremkaller visuelle associative læring i tre maur gjennom klassisk betinging. I dette paradigmet, er maur utnyttet og presentert med en visuell pekepinn (blå papp), betinget stimulans (CS), sammen med en appetitive sukker belønning, ubetinget stimulans (USA). Maur utføre en Maxilla-Labium forlengelsen refleks (MaLER), ubetinget svaret (UR), som kan brukes som en avlesning for læring. Treningen består av 10 prøvelser, atskilt av en 5-minutters intertrial intervall (ITI). Maur er også testet for minne bevaring 10 minutter eller 1 time etter trening. Denne protokollen har potensial til å tillate forskere å analysere, i en presis og kontrollert måte, detaljer om visuelle minne formasjon og nevrale grunnlaget for læring og hukommelse formasjon i tre maur.
Introduction
Insekter har vært mye brukt som modeller for å studere læring og hukommelse formasjon1. En særlig vellykket åre forskning innebærer å bruke klassisk betinging behersket dyr, som gir presis kontroll over signaler blir lært og tillater forskere å undersøke nevrale mekanismer underbygger læring og hukommelse. Fleste studier har fokusert på appetitive klassisk betinging honeybee arbeidere, APIene mellifera. Honeybee arbeiderne er opplært til å knytte en CS en amerikansk som fremkaller UR. I dette paradigmet, opprinnelig utviklet av Takeda2 og Bitterman et al. 3, UR er snabel forlengelsen Reflex (PER), USA er sukker og CS er en lukt. Bier Lær tilknytningen mellom CS og USA og kan danne en langsiktig hukommelse av denne tilknytningen.
Den opprinnelige paradigmet bruker PER som UR er brukt til å løse nervebaner underbygger olfactory læring i bier4. Den har blitt modifisert på flere måter å teste læring av ulike sensoriske stimuli, inkludert visuelle stimuli5,6,7, å innlemme aversive læring med undertrykkelse av PER8 eller Sting forlengelsen Reflex (SER)9 som UR, og teste læring i andre arter, som Humler10 og frukt fluer11. Selv om minne formasjon med flere metoder har blitt studert gjennom klassisk betinging, visuelle lære er fortsatt vanskelig å observere bruker denne tilnærmingen, selv i arter som viser en høy grad av visuelle læringsevne under foraging, som honeybees.
Nyere studier har brukt en lignende tilnærming til insekter som ikke har en snabel, som gresshopper, som utfører de Palp åpning svar (POR)12, og maur, som utfører Maxilla-Labium forlengelsen Reflex (MaLER)13. Dette har allerede avdekket fase-spesifikke læreevne som samsvarer med fase-spesifikke fôring strategier av to forskjellige ørkenen locust fenotyper, selskapelig og solitarious former, re-håndheving ideen om at minne formasjon må samsvare med økologiske behov14. Videre studier på olfactory lære om maur har vist likheter mellom maur og honeybees minne formasjon og oppbevaring, med langsiktige minne bevaring, 72 timer etter trening, er avhengig av protein syntese15.
Disse tilpasninger av den opprinnelige paradigmet har tillatt læring og hukommelse formasjon studier med mange metoder og flere modell arter. Dette er viktig for å identifisere virkningsmekanismer minne formasjon felles for insekter, men også for å identifisere ulikheter som gjenspeiler bestemt økologi av læring og hukommelse i forskjellige arter. Hovedmålet med protokollen som beskrives her er å gi en måte å utføre klassisk betinging eksperimenter med en visuell betinget stimulans med mye studert maur Art, Formica rufa. Det utstedes fra vår studie av visuelle læring i tre maur16, hvilke er likeledes en tilpasning av visuelle klassisk betinging paradigmer.
Protocol
1. vedlikeholde Formica rufa kolonier
Merk: Tre maur (Formica rufa, L.) koloniene brukes for denne studien ble samlet fra Ashdown Forest, Sussex, Storbritannia (N 51 4.680, E 0 1.800). I Storbritannia, skal F. rufa kolonier samles mellom juli og August. En god del av reiret må fjernes, inkludert flere hundre arbeidere og brood, sikre kolonien vedvarende og aktiv for lengre perioder tid (opptil ett år). Tillatelse må innhentes fra berørte myndigheter før reir fjerning.
- Huset tre maur kolonier i en bred (rundt 135 cm x 70 cm x 35 cm) med vegger dekket med polytetrafluoroethylene (PTFE) harpiks (f.eksFluon) hindre maur i å unnslippe, ved 21 ° C, en 12-h lys: 12-h mørke syklus.
- Gi sukrose (333 g/L) og vann til maur, med vann dispensere. For å sikre maurene diett er rik på proteiner, mate kolonien med frosne gresshopper 2 x en uke. Spray reiret med vann hver dag for å holde dem fuktige.
- For å forbedre de sanitære forholdene i kolonien, legge harpiks når det er nødvendig; harpiks reduserer parasitism i tre maur kolonier17. Plass en liten beholder (rundt 15 cm i diameter og 5 cm dyp) inne reiret slik at maur kan sette noen døde kolon medlemmer i den.
- Holde maur under tidligere nevnte forhold i minst 2 uker etter å samle før du prøver noen eksperimenter. Fjern sukrose fra redet å sulte maurene 2 d før eksperimenter, og dermed bedre motivasjon.
2. velge og utnytte maur
Merk: For å utnytte maur, skreddersydde innehaver er nødvendig. Dette kan bli bygget ved hjelp modellering leire og papp med en kuttet åpne knyttet til den horisontalt. Den øvre overflaten av papp bør være delvis belagt med voks, tillater insekt pinnene skal festes under fikse. Forsiktig håndtering er nødvendig for hvert trinn i denne protokollen (inkludert vedlikehold, transport- og eksperimentet), men spesielt når utnytte maur, for å unngå å utsette dem for høye nivåer av stress før.
- Du velger maur som er motivert til å spise ved Plasser et lysbilde med en dråpe sukrose (200 g/L) inne en liten boks (rundt 14 cm x 8 cm x 5 cm) med vegger dekket med PTF harpiks å hindre rømning. Ikke Velg maur som bærer tre eller døde maur fra redet. I stedet, ta maur som er crawling opp veggene i reiret, fordi disse er mer sannsynlig å være sankere prøver å forlate redet søke etter mat.
- Plasser hver maur i boksen og vente å se hvis det strømmer på sukrose miste. Hvis den does, overføre det umiddelbart til en annen tom for å hindre satiation.
- Overføre hver maur til en separat rør. Sted hver rør i fryseren for 1-2 min, eller knust is for opptil 5 min, for å nakkens maur.
- Plass immobilisert maur i holderen, gjennom kutt på papp, av felles mellom sitt hode og brystkasse. Sikre antenner forblir av hodet, bruker insekt pins innebygd i voks laget på papp.
- Bruk skreddersydd varmekabel til voks spissen av et insekt pin maur hodet, parallelt med papp. Under denne prosedyren, vil ikke touch antenner med varmeelementet wire for å unngå skade dem.
- Etter at voksen tørker, fjerne insekt pinnene holder antenner og fjern forsiktig maur fra abonnenten.
- Fastsette insekt pin holder maur i en modell leire sylinder og fikse en skreddersydde plast holderen nedenfor den sikre maur opprettholder en typisk står holdning og at hele kroppen er fri til å bevege bortsett fra hodet.
- Holde harnessed maur i mørket for minst 2 timer før.
3. opplæring og Testing
-
Oppsett og betinget stimulans unconditioned stimulans
- Gjennomføre eksperimenter i en hvit akryl boks (50 x 50 x 50 cm), åpen i fronten tillater tilgang for eksperimentator. Registrere maur atferd med et kamera.
- Plass kameraet med en makroobjektiv rett over maur gjennom et hull i den øvre overflaten av boksen.
- For å redusere eventuelle unødvendige varselsignaler, holde rommet i mørket unntatt en lyskilde peker direkte til toppen av boksen akryl (to 26-W lys).
- Bruk som en visuell indikasjon (CS) en lys blå (spektrum i Figur S1) papp rektangel (60 x 45 mm) festet på midten til en nål, koblet til en sprøyte (2 mL) som USA (sukrose 200 g/L) leveres manuelt til maur (figur 1A og 1B ).
Merk: Sucrose løsning kan gjøres med hvitt sukker, forutsatt at den ikke har farge og lukt når oppløst i vann. - Utføre to typer trening i parallell, sammenkoblet og unpaired (se forklaring nedenfor), i en randomisert rekkefølge.
Merk: I denne studien, vi ikke betaler oppmerksomhet til hva slags visuelle signalet. Bare tilknytningen mellom stikkordet og belønningen tatt i betraktning. Farge og form er ikke testet, og det er sannsynlig at CS med andre funksjoner vil gi samme resultat. Likevel ble den blå fargen plukket fordi maur fra samme slekten har vist seg å være følsomme overfor bølgelengder som dekker blått18.
- Gjennomføre eksperimenter i en hvit akryl boks (50 x 50 x 50 cm), åpen i fronten tillater tilgang for eksperimentator. Registrere maur atferd med et kamera.
-
Sammenkoblede opplæring
Merk: I sammenkoblede trening, CS og amerikanske presenteres i hver prøve, forbundet med hverandre (figur 1 c).- Starte innspillingen av maur 10 s før presentere CS for å sikre at maur ikke spontant utfører MaLER før presentasjonen. Hvis maur utfører MaLER i denne perioden, utsette rettssaken i noen sekunder. Hvis noen maur viser dette kontinuerlig, utelukke det fra analysen.
- Flytte den sprøyten + CS foran mauren for ~ 10 s, med spissen av nålen holdt mellom maur ansiktet og maksimalt 5 mm ovenfor og på hver side. Samtidig flytte tuppen av nålen så nært som mulig til maur hodet, men uten å berøre antennene (figur 1B).
Merk: Bevegelse var inkludert når presentere CS fordi det har vist seg å spille en rolle i visuelle associative læring i honeybees6. - Bruk press på utstråler en eneste dråpe sukrose fra sprøyten spissen og plassere slipp av sukrose antenner og munndeler tillate maur å oppdage sukrose. Tillate maur til mat for ca 5 s.
Merk: Mengden av sukrose maur forbrukes i hvert forsøk var ikke kontrollert i disse eksperimentene. Slipp av Sukrose på som maur feed bør være store nok til å tillate dem å fritt mate for ca 5 s. - Gjenta denne fremgangsmåten 10 x, med en ITI 5 min.
Merk: I disse eksperimentene, maur var alltid slått til høyre, med sitt høyre øye nær den åpne siden av boksen akryl. Derfor nærmet CS alltid maurene fra sine høyre. Selv om dette ikke opphever læring, kan disse eksperimentene utføres ved å slå halvparten av maur til venstre og halvparten til høyre, for å unngå noen mulige lateralization effekt.
-
Kort opplæring
Merk: Denne opplæringen består i å presentere maurene CS eller USA separat, disse to stimuli som, dermed atskilt fra hverandre over tid (figur 1 d).- Starte innspillingen av maur 10 s før hvert forsøk.
- Presentere CS på samme måte som i sammenkoblede trening men ikke leverer sukrose for maur.
- Etter 2,5 min, levere USA direkte til munndelene (også berøre antenner) bruker en sprøyte uten CS knyttet.
- Start med en CS-rettssak og gjenta dette 10 x for hver prøve, intercalating dem med en ITI 2,5 min.
-
Testing
Merk: En test bør være gjennomført bare 1 x, enten 10 min eller 1 time etter siste trening rettssaken; en test som maur er presentert med CS, men ikke med USA føre til utryddelse av assosiative minnet dannet tidligere.- Starte innspillingen av maur 10 s før testing.
- Presentere CS maur for ca 10 s.
- Kontroller at maur er fortsatt motivert til å mate ved å levere sukrose etter testen.
4. datainnsamling, analyse
- Registrere maur atferd, ovenfra, 10 s før hvert forsøk og under CS og amerikanske presentasjonene. Kontroller at alle prøvelser og tester registreres for bakre analyse.
- Bruke opptak gjort under trening og testing, score maurene svar under 10 s CS presentasjonen.
- Skille maurene svar under CS presentasjonen i tre typer atferd, avhengig av filtypen og bevegelse av munndelene: Full forlengelse med bevegelse (FEM) som om fôring, Full forlengelse uten bevegelse (FE) eller delvis forlengelsen (PE) av den maxilla-labium eller maxillary palps (figur 2A - 2D). For analyse, kan du gruppere alle MaLER typer som et enkelt svar (figur 2E).
- Ekskludere noen maur som gjorde ikke feed i hver eneste prøve og test.
Merk: For analyse, en statistisk test som tar hensyn til individuelle maur anbefales, og dermed regnskap for variasjon i individer. Ved klassifisering maur svar under trening på en binær måte (1 positiv respons og 0 for nei svaret), er logistisk regresjon med blandet effekter tilrådelig16,19. For å sammenligne andelen av maur svarer i hver trening prøve eller test, en G- eller Fishers pressepenger testen anbefales, avhengig av antall maur analysert16,20.
Representative Results
Under klassisk betinging eksperimenter, må CS ikke få en spontan reaksjon i dyr. I eksperimentene vi gjennomførte, utføres bare 3-4% av maur MaLER som svar på den visuelle indikatoren på første rettssaken, før. Maur som gjennomgikk en sammenkoblet trening utført stadig mer MaLER svar til CS (figur 3A; logistisk regresjon, N = 51, frihetsgrader (df) = 507, z = 5.949, p < 0,01). Prosentandelen av sammenkoblede maur som svarer til CS platå rundt 50%, fra tredje rettssaken og utover. Tvert imot, hender maur viste ingen betydelig økning i MaLER under trening (figur 3B; logistisk regresjon, N = 29, df = 287 z = 0.758, p = 0.45). Forekomsten av MaLER som svar på den visuelle pekepinnen var signifikant høyere under sammen enn unpaired trening (logistisk regresjon, N = 80, df = 796, z =-5.306, p < 0,01), som var sanne for hver prøveversjon unntatt først (tabell 1).
Maurene ble testet for å undersøke deres kort- og en mellomlang sikt minne15, 10 min eller 1 time etter siste trening rettssaken. For begge tester var andelen av maur utfører MaLER svar til CS signifikant høyere når de hadde gjennomgått sammen i stedet for kort opplæring (Figur 3 c og 3D; Tabell 1).
Under trening, personlige maur viste betydelige variasjoner i antall og type av MaLER som de vises (figur 4A og 4B). Bare 14% av sammenkoblede maur reagerte på hver prøveversjon fra andre eller tredje rettssaken videre, mens de fleste maur vekslet mellom forsøk der de svarte og som de gjorde ikke. På disse opplæring prøvelser som svarte maur, grad de utvidet og flyttet sine munndeler variert. Derfor vi delt MaLER i tre forskjellige typer: FEM, FE eller PE. Vanligvis utført maur FEM eller PE oftere enn FE. Men utført bare noen maur konsekvent samme type svar; i de fleste tilfeller viste maur litt konsistens på typen MaLER de utført (Figur 4).

Figur 1 : Eksperimentelle set-up og opplæringsprogram. (A) fastsette maur med voks til et insekt pin festet til en modellering leire sylinder (oransje) og sted en skreddersydd holder under det å tillate en naturalistisk holdning. Plass den inne en hvit akryl boks opplyst av to lyskilder, rett under et kamera. Bruk som betinget stimulans (CS) papp (blå firkant) koblet til sprøyten som leverer unconditioned stimulans (oss), en sukker belønning. Den innfelt viser et nærbilde av en maur i abonnenten. (B) spissen av nålen flyttes så nært som mulig til maur hodet, men uten å berøre antenner som vist i dette skjematisk. Tog maur gjennom enten en (C) sammen eller (D) unpaired trening. Dette tallet har blitt endret fra Fernandes et al. 16. Klikk her for å se en større versjon av dette tallet.

Figur 2 : The Maxilla-Labium forlengelsen svar av tre maur. Enkeltbilder fra videoopptak viser maur munn del bevegelser under trening. (A) dette panelet viser ingen respons. (B) dette panelet viser en full forlengelse (FE) av maxilla-labium som ender i glossa. (C) denne siden viser filtypen delvis (PE) med bare den maxillary palpus synlig. (D) dette panelet viser en delvis utvidelse (PE) av maxilla-labium strukturer. (E) maur gjennomgår sammenkoblede trening (N = 51) utfører en full forlengelse med bevegelse (FEM, mørk brun), en FE (midten av brun) eller PE (lys brun). Dette tallet har blitt endret fra Fernandes et al. 16. Klikk her for å se en større versjon av dette tallet.

Figur 3 : Tre maur skjemaet assosiativ minner om en visuell pekepinn sammen med sukker belønning. (A) prosentandelen av sammenkoblede maur (N = 51) utfører MaLER svar på CS presentasjonen betydelig økt gjennom trening. (B) prosentandelen av maur utfører MaLER ikke økte betydelig gjennom kortet trening (N = 29). Maur var testet (C) 10 minutter (sammenkoblede: N = 15; unpaired: N = 15) eller (D) 1 time (sammenkoblede: N = 15; unpaired: N = 14) etter siste trening rettssaken. Prosentandelen av maur svarer til CS under og etter sammenkoblede eller kortet trening er representert i brune eller grå, henholdsvis. De tre typene MaLER er representert i mørket (FEM), medium (FE) og lys (PE) toner. Dette tallet har blitt endret fra Fernandes et al. 16. Klikk her for å se en større versjon av dette tallet.

Figur 4 : Individuelle resultater av maur under trening. Disse skjermbildene viser individuelle forestillinger av (A) sammenkoblede maur og (B) unpaired maur. De tre typene MaLER er representert i mørket (FEM), medium (FE), og lysbrun (PE) eller grå. Dette tallet har blitt endret fra Fernandes et al. 16. Klikk her for å se en større versjon av dette tallet.
| Rettssaken | N | DF | G (justert) | P |
| 1 | 80 | na | na | > 0,1 |
| 2 | 80 | 1 | 3.86 | < 0,05 |
| 3 | 80 | 1 | 8.41 | < 0,01 |
| 4 | 80 | 1 | 6.63 | < 0,01 |
| 5 | 80 | 1 | 8.41 | < 0,01 |
| 6 | 80 | 1 | 7.5 | < 0,01 |
| 7 | 80 | 1 | 10.69 | < 0,01 |
| 8 | 80 | 1 | 11.76 | < 0,01 |
| 9 | 80 | 1 | 17.13 | < 0,01 |
| 10 | 80 | 1 | 17.13 | < 0,01 |
| 10 min | 59 | 1 | 5.5 | < 0,05 |
| 1 h | 59 | 1 | 4,42 | < 0,05 |
Tabell 1: sammenligning av MaLER svar til CS mellom maur som hadde gjennomgått sammen og unpaired trening, for hver prøve og test Antall maur (N), frihetsgrader (df), G-test av uavhengighet (G) og p-verdien vises. Den første rettssaken ble analysert med et Fisher eksakt test. Denne tabellen er endret fra Fernandes et al. 16.
Discussion
Klassisk betinging er en av de mest godt etablerte paradigmene å studere læring og hukommelse. Protokollen vi presentert her er en tilpasning av paradigmet for honey bee arbeidere2,3 og senere brukes med flere andre arter, som Humler, frukt fluer, som også bruker PER som en presentasjon for å lære10 ,11, og gresshopper og maur, som bruker POR og MaLER,12,13henholdsvis. Bruker denne protokollen, er det mulig å trene brukt tre maur å lære tilknytningen mellom en visuell pekepinn og sukker belønning og analysere oppbevaring av denne kort - (10 min) og midt-term (1 time) minne16.
I noen atferdsdata eksperimentet er det nødvendig å ta hensyn avgjørende skritt som kan minimere variasjon i dyr svar. I protokollen presenteres her, er flere tiltak tatt å minimere variasjon før og under trening. Før starten av eksperimenter, kolonien må være sultet i minst to dager og maur bør velges basert på deres vilje til å spise fra en sukker nedgang i boksen holder. Hvis du velger maur på denne måten er ment å maksimere sjansen for trening maur som er motivert til å mate. Forsiktig håndtering er også en viktig faktor fordi det kan hjelpe for å redusere stressnivået, som forstyrrer læring hvis det er for intens21. Dette bør maur være anesthetized med kaldt å bo urørlig mens blir utnyttet, fordi enhver bevegelse (for å unnslippe) under denne prosedyren kan være en kilde til stress. Videre bør kontakten mellom mauren og voksen være minimal, unngå kontakt mellom antenner og varm voks eller ledning, kan skade. Selv om disse observasjonene ikke har analysert formelt, syntes antennene å gå med et bestemt mønster i læring.
Under eksperimenter er forsiktig levering av sukker også viktig å holde maur motivert. Igjen, mens dette ikke er risikoanalysert formelt, brå mat levering syntes å forårsake ekstra stress til maur, som i sin tur førte til mangel på motivasjon og læring. Videre sukrose levert under trening bør være en redusert konsentrasjon (200 g/L) å unngå satiation før slutten av trening og testing. Dette gjør MaLER å være en god kandidat ubetinget svaret fordi, sammen med en lav spontan ytelse av dette svaret til den visuelle indikatoren, det også ikke mette over prøvelser. Til slutt, i motsetning til de fleste klassisk betinging studier2,3,5,6,7,8,9,10, 11 , 12 , 13, vi trente en maur samtidig til slutten av eksperimentet, forlate det på plass mellom forsøk i stedet for å fjerne den for å teste en maur. Trene flere maur sammen syntes å produsere mer variabel resultater som kan skyldes en økning i stress og/eller konflikt mellom visuell informasjon forårsaket av den totale endringen av landskapet. For å redusere varigheten av hvert eksperiment, brukte vi en 5-minutters ITI i stedet for 10 minutters ITI brukes i de fleste klassisk betinging studier16. Selv om alle disse hensyn bør hjelpe sikre at maur er motivert til å mate og lære under trening, unngås ikke noe variasjon. Vi anbefaler å bruke maur som synes å ha sosial og appetitive bevegelse normalt og eksklusive maur fra analysen øyeblikket de ikke klarer å spise en trening rettssak eller en test.
Hva slags CS ble ikke testet i denne studien. Selv om vi har brukt en blå visuelle stimulans fordi maur av samme slekten er følsomme for disse bølgelengder18, kan også andre farger læres i samarbeid med en belønning. Ytterligere eksperimenter vil være nødvendig for fullt karakterisere fargene blir sett og lært i dette oppsettet. Dette gjelder også for forskjellige former og størrelser av den visuelle indikatoren. Vi har ikke testet om maur romlig oppløsning ville være tilstrekkelig for å skille visuelle stimulans presentert her på avstand fra maurene øyne det ble presentert på. Selv om tre maur øyne har blitt beskrevet størrelse og antall fasetter22, til vår kunnskap, har deres romlig oppløsning ikke blitt fullt beskrevet ennå. Men er dette beregnet for Melophorus magoti23. Lignende karakteristikk av tre maurene eller andre testet insekter øyne ville bidra til en klar undersøkelse av funksjonene av visuell indikasjon blir observert og lært av dyrene. Videre, vi inkludert bevegelse presentasjonen visuelle stimulans for maur fordi det har vist seg å spille en rolle i honeybee associative læring i klassisk betinging6. Men kan dette ikke ble testet i denne studien, og på grunn av flygende insekter i forhold til gangavstand insekter forskjellige bevegelse, forskjellene mellom honeybee og maur visuelle klassisk betinging observeres.
På et endelig notat kunne vi ikke undersøke langsiktige minne bevaring fordi maur ikke overlevde blir utnyttet for slike lange perioder etter trening. Men i senere sett av eksperimenter, vi har holdt maur i live og motivert til å spise og lære når utnyttet og forlot dem over natten i et mørkt og fuktig miljø (plassere en boks over dem). Dette paradigmet kan derfor brukes til å løse langsiktige minne bevaring av tre maur, i tillegg til kort og midten sikt minne.
Med dette enkel prosedyre tilpasset fra generelle klassisk betinging paradigmer, er det mulig å studere erverv og oppbevaring av visuelle minner i brukt tre maur, som har vært undersøkt mye i paradigmer bruke fritt bevegelige dyr. Dette paradigmet har potensial til å brukes for å analysere nevrale grunnlaget for visuell læring i en svært godt etablerte modell for insekt navigasjon.
Disclosures
Forfatterne ikke avsløre.
Acknowledgments
Forfatterne takker Tom Collett og Cornelia Buehlmann for å dele verdifull informasjon om innsamling og vedlikehold av tre maur kolonier. Forfatterne takker også Justine Crevel for kommenterer på tidligere versjoner av denne artikkelen.
Materials
| Name | Company | Catalog Number | Comments |
| Fluon | Blades Biological Ltd, Edenbridge, UK | ACS 109; ACS 112; ACS 114 | For preventing insects from scaping |
| Crickets | Blades Biological Ltd, Edenbridge, UK | LZJ 217 | Given to the ant colonies as protein source |
| Natural Pine Rosin/Resin | Minerals-water Ltd, Rainham, UK | 500g | Given to the ant colonies for sanitation |
| Austerlitz Insect Pin | Fine Science Tools GmbH, Heidelberg, Germany | 26000-40 | For harnessing ants |
| High speed camera | Edmund Optics Inc., Barrington, USA | eo-13122M | MaLER recordings during training and testing |
| Macrolens | Cannon, Surrey, UK | EF 100 mm f/2.8 L Macro IS USM | MaLER recordings during training and testing |
| Software | IDS Imaging Development Systems GmbH | uEye64 | MaLER recordings during training and testing |
| Blue Cardboard | john smith's at Union Store, University of Sussex | JACK-PJM41358 | Constitutes de conditional stimulus |
| Syringe | Fisher Scientific LTD, Loughborough, UK | BD Plastipak 300185case; Product Code.12369289 | US and CS (attached) presentation |
| Needle (0.5 x 16 mm) | Fisher Scientific LTD, Loughborough, UK | BD Microlance 300600; Product Code:10442204 | US and CS (attached) presentation |
References
- Chittka, L., Niven, J.
- Takeda, K. Classical conditioned response in the honey bee. Journal of Insect Physiology. 6, 168-179 (1961).
- Bitterman, M. E., Menzel, R., Fietz, A., Schäfer, S. Classical conditioning of proboscis extension in honeybees (Apis mellifera). Journal of Comparative Psychology. 97 (2), 107-119 (1983).
- Menzel, R., Muller, U. Learning and memory in honeybees: from behavior to neural substrates. Annual Review of Neuroscience. 19 (1), 379-404 (1996).
- Hori, S., Takeuchi, H., Kubo, T. Associative learning and discrimination of motion cues in the harnessed honeybee Apis mellifera L. Journal of Comparative Physiology A. 193 (8), 825-833 (2007).
- Balamurali, G. S., Somanathan, H., De Ibarra, N. H. Motion cues improve the performance of harnessed bees in a colour learning task. Journal of Comparative Physiology A. 201 (5), 505-511 (2015).
- Mujagić, S., Würth, S. M., Hellbach, S., Dürr, V. Tactile conditioning and movement analysis of antennal sampling strategies in honey bees (Apis mellifera L). Journal of Visualized Experiments. (70), e50179 (2012).
- Wright, G., et al. Parallel reinforcement pathways for conditioned food aversions in the honeybee. Current Biology. 20 (24), 2234-2240 (2010).
- Vergoz, V., Roussel, E., Sandoz, J. C., Giurfa, M. Aversive learning in honeybees revealed by the olfactory conditioning of the sting extension reflex. PloS One. 2 (3), e288 (2007).
- Laloi, D., et al. Olfactory conditioning of the proboscis extension in bumble bees. Entomologia Experimentalis et Applicata. 90 (2), 123-129 (1999).
- Chabaud, M. A., Devaud, J. M., Pham-Delègue, M. H., Preat, T., Kaiser, L. Olfactory conditioning of proboscis activity in Drosophila melanogaster. Journal of Comparative Physiology A. 192 (12), 1335-1348 (2006).
- Simões, P., Ott, S. R., Niven, J. E. Associative olfactory learning in the desert locust, Schistocerca gregaria. Journal of Experimental Biology. 214 (15), 2495-2503 (2011).
- Guerrieri, F. J., d'Ettorre, P. Associative learning in ants: conditioning of the maxilla-labium extension response in Camponotus aethiops. Journal of Insect Physiology. 56 (1), 88-92 (2010).
- Simões, P. M., Niven, J. E., Ott, S. R. Phenotypic transformation affects associative learning in the desert locust. Current Biology. 23 (23), 2407-2412 (2013).
- Guerrieri, F. J., d'Ettorre, P., Devaud, J. M., Giurfa, M. Long-term olfactory memories are stabilised via protein synthesis in Camponotus fellah ants. Journal of Experimental Biology. 214 (19), 3300-3304 (2011).
- Fernandes, A. S. D., Buckley, C. L., Niven, J. E. Visual associative learning in wood ants. Journal of Experimental Biology. 221 (3), 173260 (2018).
- Christe, P., Oppliger, A., Bancala, F., Castella, G., Chapuisat, M. Evidence for collective medication in ants. Ecology Letters. 6 (1), 19-22 (2003).
- Aksoy, V., Camlitepe, Y. Behavioural analysis of chromatic and achromatic vision in the ant Formica cunicularia (Hymenoptera: Formicidae). Vision research. 67, 28-36 (2012).
- Bates, D. M. lme4: Mixed-Effects Modeling with R. , Springer. New York, NY. (2010).
- Sokal, R. R., Rohlf, F. J. Biometry (3rd edn). , W.H. Freman and Company. New York, NY. (1995).
- Bateson, M., Desire, S., Gartside, S. E., Wright, G. A. Agitated honeybees exhibit pessimistic cognitive biases. Current Biology. 21 (12), 1070-1073 (2011).
- Perl, C. D., Niven, J. E. Differential scaling within an insect compound eye. Biology letters. 12 (3), 20160042 (2016).
- Schwarz, S., Narendra, A., Zeil, J. The properties of the visual system in the Australian desert ant Melophorus bagoti. Arthropod Structure & Development. 40 (2), 128-134 (2011).
Tags
Atferd problemet 140 varselsignaler appetitive condition læring minne MaLER Formica rufaErratum
Formal Correction: Erratum: Visual Classical Conditioning in Wood Ants
Posted by JoVE Editors on 12/31/1969.
Citeable Link.
An erratum was issued for: Visual Classical Conditioning in Wood Ants. The Acknowledgments section was updated, and a supplemental figure was added.
The Acknowledgments sections was updated from:
The authors thank Tom Collett and Cornelia Buehlmann for sharing valuable information regarding collection and maintenance of wood ant colonies. The authors also thank Justine Crevel for commenting on previous versions of this article.
to:
The authors thank Tom Collett and Cornelia Buehlmann for sharing information regarding collection and maintenance of wood ant colonies. The authors also thank Justine Crevel for commenting on previous versions of this article, and Nora Nevala for measuring the spectrum intensity of the visual stimulus. This work was supported by a BBSRC grant to JEN (grant number BB/R005036/1). All the data pertaining to this manuscript are published in the University of Sussex Research Data Repository online database (10.25377/sussex.5794386).
The following supplemental figure was added to the end of the Representative Results section:
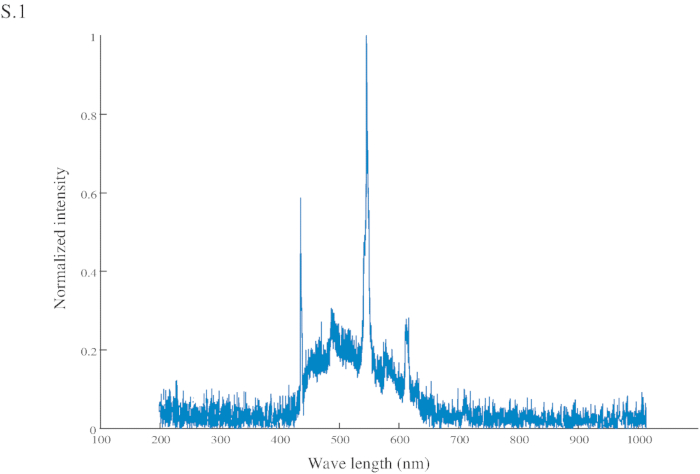
Figure S1: Normalized intensity of the conditional stimuli (CS). The CS has a peak intensity at 545 nm (in the green range) and another at 435 nm (in the blue range).
