

Summary
פרוטוקול להעריך את האבולוציה של הזמן. העצבי לגירויים חוזרים חיצוניים מוצג הקלטות מצב יציב של אותו מצב ניסיוני נרכשים וממוצעים בתחום הזמן. הדינמיקה מצב יציב מנותח על ידי התוויית משרעת התגובה כפונקציה של זמן.
Abstract
מתייחס לסנכרון של הפעילות העצבית. לתקופתיות של גירויים חושיים סנכרון זה מגדיר את הדור של תגובות מעורר המצב היציב (כלומר, תנודות בשלב האלקטרונצגרם-נעול לגירוי הנהיגה). הפרשנות הקלאסית של משרעת התגובה היציבה של המצב היציב מניח תגובת הזמן הסטריאוטיפית ותנודות ברקע אקראי, כגון בממוצע על מצגות חוזרות ונשנות של הגירוי משחזרת את ה התגובה סטריאוטיפית. גישה זו מתעלמת מדינאמיקה של המצב היציב, כמו במקרה של הסתגלות שמעוררת על ידי חשיפות ממושכות לגירוי. כדי לנתח את הדינמיקה של תגובות מצב יציב, ניתן להניח כי האבולוציה הזמן של משרעת התגובה היא זהה בגירוי שונים מופרדים על ידי הפסקות ארוכות מספיק. מבוסס על הנחה זו, שיטה לאפיון האבולוציה הזמן של תגובות מצב יציב מוצג. מספר גדול מספיק של הקלטות נרכשים בתגובה לאותו מצב ניסיוני. מסלולים ניסיוניים (הקלטות) הם הממוצע עמודה (כלומר, פועל ממוצעים אבל התקופה בתוך הקלטות אינם ממוצעים עם הקטעים הקודמים). ממוצע העמודות בממוצע מאפשר ניתוח של תגובות למצב יציב בהקלטות עם יחסי אות לרעש גבוהים במידה ניכרת. לכן, האות הממוצע מספק ייצוג מדויק של האבולוציה הזמן של התגובה מצב יציב, אשר ניתן לנתח הן בתחומי הזמן והתדר. במחקר זה, מסופק תיאור מפורט של השיטה, באמצעות המצב היציב הפוטנציאל מעורר חזותית כדוגמה לתגובה. היתרונות והאזהרות מוערכים על בסיס השוואה עם שיטות בודדות שנועדו לנתח את התנועה העצבית.
Introduction
כאשר הוקלט מתוך הקרקפת, פעילות המוח החשמלי הוא נצפתה שינויים רציפים וקבועים במתח לאורך זמן. פעילות חשמלית זו נקראת אלקטרונצאלוגרם (EEG) ותוארה לראשונה על ידי הנס ברגר בשנות העשרים המאוחרות שלהמאה הקודמת. מחקרים הזרע הבאים תיאר את EEG כסדרה בזמן מורכב, שבו דפוסים שונים קצבית או חוזרים ניתן להבחין2,3,4. בימים אלה, EEG מחולק בדרך כלל לחמש להקות תדר מבוססות היטב, דלתא, תטא, אלפא, בטא וגמא, המשויכים לתהליך החושי והקוגניטיבי השונים.
במשך שנים, המחקר של תנודות במוח באמצעות EEG היה מוגבל לניתוח של הספקטרום בפעילות מתמשכת או שינויים בפעילות מנדנוד שעורר על ידי אירועים סנסוריים שאינם תקופתיים. בעשורים האחרונים יושמו מתודולוגיות שונות ליצירת תנודות מתמשכות של EEG ולחקר ההשפעות של מודולונים כאלה על תהליכים תפיסתית וקוגניטיביים, כולל הצגת גירוי חושי קצבי ל גרימת הסתבכות עצבית. המונח "כניסה עצבית" מתייחס לסנכרון הפעילות העצבית עם המאפיינים התקופתיים של גירויים חושיים. תהליך זה מוביל לדור הפוטנציאל העורר של המצב היציב (כלומר, תנודות EEG שננעלו על המאפיינים התקופתיים של גירוי הנהיגה). המצב היציב של הפוטנציאל המתמשך מעורר בדרך כלל על ידי גירוי חזותי, שמיעתי ומישוש, תוך שימוש בגירויים ארעיים המוצגים בקצב קבוע או גירוי רציף מאופנן במישרעת בתדירות הריבית. הואיל והוא מוקלט בתגובה לגירוי מישוש חוזר של5,6, מצב יציב ומעורר באופן חזותי (ssseps) מוקלטים בדרך כלל על ידי התקופתיות הצגה של הבהוב בוהק, תמונות ופרצופים7,8. תגובות שמיעה המדינה יציבה (assrs) מופקים בדרך כלל על ידי רכבות של גירויים אקוסטיים חולף או על ידי הצגת רציפה של משרעת-הטונים מאופנן9,10.
החילוץ של המצב יציב הפוטנציאל מעורר מ EEG נמדד למעשה מסתמך על ממוצע כתוצאה מכך נרכש לאחר מכן הזמן הנעלה של הגירוי11. בשל התקופתיות של התגובות, ניתן לנתח אותם בתחומים הזמן והתדר. לאחר שינוי השכיחות בתחום, התגובה החושית נצפתה כפסגות של משרעת בקצב המצגת או תדירות האפנון של הגירויים החיצוניים, וההרמוניה המקבילה שלהם. הליכים אלה (תחום הזמן ממוצע ושינוי התדר העוקב אחר התחום) היו חיוניים לפיתוח מבחן שמיעה המבוסס על איתור שיטות ה-assr עם מטרות קליניות12,13,14 ,15,16.
יתרה מזאת, התחום הקלאסי של הזמן ממוצע בממוצע של מוצרי EEG היה שימושי ביותר לניתוח תהליכים פיזיולוגיים כגון הדור וההכחדה של ssvep17,18. הצגת רכבות רצופות של אורות הבהובים וממוצע התקופות הבאות בהקלטה, וואקר ואח '19 ציין כי מדד נעילת הפאזה של ה-SSVEP גדל במהירות במהלך 400 אלפיות הראשונה של גירוי ונשאר גבוה לאחר מכן . הם דיווחו גם כי חסון ויזואלי הוקמה בין 700-1 100 ms לאחר התפרצות גירוי. מידה מסויימת של האנריינט נשארה אפקטיבית לאחר היסט הרכבת הגירוי, שנמשכה כשלוש תקופות של התגובה הנדנוד17,19. התנהגויות אלה יפורשו כאפקט מרתק/מנתק של תנודות שנצפו, שהיא תוצאה של עיבוד מידע לא לינארית במערכת הראייה האנושית17. לחילופין, ידוע כי בתנאים ניסיוניים מסוימים, גירוי הבהוב יכול לעורר את התגובות בהתחלה, ומחוץ לתגובות בתום הרכבות הגירוי במקום במצב של הסתבכות עצבית18.
ההנחה העיקרית הממוצעת ברצף EEG שנרכשו ברציפות היא כי אות EEG מייצג שילוב ליניארי של התגובה החושית ואת רעש הרקע20. יתר על כן, משרעת, התדר, והשלב של התגובה מנדנוד מניחים להיות נייח, ואילו רעשי הרקע נחשבים כפעילות אקראית. עם זאת, במקרים שבהם לא מתקיימים הנחה זו, משרעת התגובה שחושבה לאחר שמספר התקופות אינן מתאימות בהכרח לשרעת היכולת הנקודתית של הפוטנציאל העורר.
זה כבר דווח לאחרונה כי assr שנוצר במוח של חולדות מסתגל למצגת רציפה של משרעת-הטונים מאופנן (כלומר, משרעת התגובה ירידה אקספוננציאלית לאורך זמן)21,22. הסתגלות הייתה מתורגמת כמנגנון עצבי המשקף את אובדן החדשנות של גירוי חושי חדגוניות חוזר, ומגבירה את הרגישות לתנודות רלוונטיות בסביבה האקוסטית23,24. במסלול השמיעה, הסתגלות עשויה לשפר את הבנת הדיבור בסביבות רועשות. בנוסף, תהליך זה עשוי להיות חלק ממנגנון קיים כדי לפקח על המשוב השמיעתית של הקול עצמו כדי לשלוט בייצור הדיבור.
ניתוח האבולוציה של הזמן של 40 Hz AR בבני אדם, ואן אייאקלי ואח '25 נצפתה ירידה משמעותית אך קטנה במשרעת התגובה לאורך זמן (סביב-0.0002 μv/s מבוסס על ניתוח קבוצתי, כאשר בהנחה ירידה ליניארית לאורך זמן). כתוצאה מכך, מחברים אלה הסיקו כי 40 Hz ASSR בבני אדם אינו מסתגל לגירוי. בבני אדם, התנהגויות לא נייח נצפו בעת ניתוח היציבות של SSVEP26. מחברים אלה הבחינו שהמשרעת של התדר הבסיסי וההרמוניה השנייה של SSVEP היו נייחים רק ב -30% ו-66.7% מהנושאים שהם בדקו, בהתאמה. השלבים הן של רכיבי תדר SSVEP, למרות יציב יחסית לאורך הזמן,הציגו קטן של22.
לכן, למרות שתחום הזמן הקלאסי ממוצע בממוצע של כרכים שנרכשו לאחר מכן מאפשר לחקור מאפיינים נייחים של מתודולוגיה עצבית, המתודולוגיה הזו צריכה להיות מתוקנת כאשר דינמיקה ארוכת טווח של התנועה היא המוקד של ה מחקר, או כאשר הממוצע של דינמיקה לטווח קצר פגום על ידי התרחשות של דינמיקה ארוכת טווח. כדי לאפיין התנהגויות שאינן נייחות של תגובות המצב היציב, התגובה המחושבת שחושבה בחלון זמן נתון לא אמורה להיות מסוכצת על-ידי אלה שחושבו במקטעי ה-EEG הקודמים. במילים אחרות, יש להפיק את הפוטנציאל העורר מרעשי הרקע מבלי להיות בתחום הזמן הממוצע עם מקטעי ה-EEG הקודמים.
במחקר זה, שיטה להערכת הדינמיקה של האנטריה העצבית מוצגת. תגובות מצב יציב נרשמות באופן חוזר בתגובה לגירוי זהה, שבו הקלטות רצופות משולבים על ידי מרווח מנוחה של שלוש פעמים את אורך הריצה ניסיוני. בהתחשב בכך שאם האבולוציה בזמן של התגובה הפיסיולוגית זהה ברצפים שונים של ניסיוני עצמאיים (הקלטות עצמאיות), הקלטות הן מבחינת טור וממוצעים. במילים אחרות, ממוצע האפוקסי התואם לאותו מיקום בהקלטות השונות מקביל, ללא חישוב בממוצע בהקלטה. כתוצאה מכך, משרעת התגובה שחושבה בכל מרווח גירוי, יתאימו לשרעת הזמן הנקודתית של הפוטנציאל העורר. ניתן לנתח את התגובות התחושתיות בזמן התחום או להפוך לתחום התדרים, בהתאם למטרת הניסוי. בכל מקרה, ניתן להתוות את ההגברה כפונקציה של זמן לניתוח התפתחות הזמן של תגובת המצב היציב. הדור והכחדה של פוטנציאל המצב היציב ניתן להעריך על ידי הגבלת הניתוח לבין הראשון והאחרון של ההקלטות.
ניתן לנתח את הדינמיקה של המעטפת העצבית באמצעות גישות אחרות, כגון סינון אמצעות של מדידות חד-מתליות סביב תדירות הריבית והמחשוב המעטפה של אות החשמל באמצעות סינון מעבר נמוך25 וה ההמרה של הילברט27. בהשוואה למתודולוגיות אלו, ממוצע העמודים בממוצע הטורים מאפשר למיחשוב פרמטרים של מצב יציב ומבוסס על אותות עם יחס אותות הרעש הגבוה יותר (SNR). לאחרונה, מסנן קלמן התפתחה כטכניקה מבטיחה להערכת ההגברה של 40-הרץ המוני28,29,30. יישום הסינון של קלמן יכול לשפר את הזיהוי של התגובות למצב היציב הקרוב יותר להסף האלקטרו-פיזיולוגי ולצמצם את זמן מבחן השמיעה29. יתרה מזאת, אין צורך בתגובות נייחות להנחה כאשר משתמשים בגישה לסינון קלמן להערכת משרעת ה-ASSR30. למרות זאת, רק מחקר אחד ניתח את האבולוציה של ה-ASSRs באמצעות הסינון של קלמן25. סיום המחקר הוא כי משרעת ה-ASSR 40-Hz יציבה על מרווח הגירוי. לכן, סינון קלמן צריך להיבדק בתנאים שבהם ה-ASRR אינו נייח.
למרות שהשיטה צורכת זמן רב, השיטה הממוצע לעמודות היא ללא מודל ואינה זקוקה לערכי אתחול ו/או הגדרות פריורי של התנהגות הרעש. יתרה מזאת, כיוון שהיא אינה כרוכה במועדי התכנסות, ממוצע הטורים הממוצע עשוי לספק ייצוג אמין יותר של התפרצות התנועה העצבית. לפיכך, התוצאות המתקבלות באמצעות השיטה הממוצע לעמודות יכולות להיחשב כאמת הקרקעית לניתוח דינמיקה של השימוש העצבי בעזרת הסינון של קלמן.
תיאור זה של הפרוטוקול מבוסס על דוגמה של SSVEP. עם זאת, חשוב לציין כי השיטה המוצגת כאן היא ללא מודאליות, כגון זה יכול לשמש גם כדי לנתח את האבולוציה הזמן של SSSEP ו-ASSR.
Protocol
המחקר הנוכחי בוצע תחת אישור ועדת המחקר והאתיקה של האוניברסידה וואלפאראיסו, צ'ילה (קוד משפט הערכה CEC170-18), אישר את ההנחיות הלאומיות למחקר עם בני אדם.
1. הכנה
- . ברוך הבא לנושא
- הסבירו את המטרות והרלוונטיות של המחקר. לספק תיאור של פרטים טכניים רלוונטיים. ענה על כל השאלות ביסודיות.
- ציין במפורש כי היא/הוא יכול לקטוע את הפגישה ניסיוני בכל עת אם תרצה.
- בקשו ממתנדב לקרוא את הסכמת הנושא ולחתום על הטופס המתאים. הפסקת ההפעלה הניסיונית אם הסכמה מושכלת אינה מתקבלת.
2. הכנה לנושא
- בקשו מהנושא לשבת בכיסא מעבדה במצב נוח.
- נקה את הקרקפת עם אתנול (פתרון ב 95%) כדי להסיר את השכבה של תאי עור מתים וחלב המכסה אותו. שלב זה חשוב להפחית את העכבה בין אלקטרודות לקרקפת.
- למדוד את היקף הראש עם קלטת מדידה כדי להגדיר את גודל כובע האלקטרודה לשמש.
- בקשו מהנושא לענוד את כובע האלקטרודה. ספק את ההוראות למיקום נוח אך נכון של המכסה.
- למדוד את המרחק בין nasion (Nz, את הנקודה האמצעית של תפר הקדמית, אשר ניתן לזהות על ידי הדיכאון בין העיניים והחלק העליון של האף) ואת inion (איז, הגדולה של עצם העורף) באמצעות קלטת מדידה.
- למדוד את המרחק בין הנקודות השמאלי והימני מראש (מזוהה כדיכאון ממש לפני האוריקל של האוזניים) באמצעות קלטת מדידה.
- תקן את המיקום של כובע האלקטרודה, כך הצטלבות בין הקווים הדמיוניים שהוגדרו בשלבים הקודמים מתאימים לקודקוד הראש. ודא שהנושא נוח לאחר ההתאמות.
- הניחו ג'ל מוליך במחזיקי האלקטרודות, בהתאם למיקומים הנחשבים לניסוי.
- השתמש 64 מיקומים הקרקפת בעקבות הבינלאומי 10-20 מערכת31 להשתמש בתוצאה של הפרוטוקול לבצע ניתוח לוקליזציה מקור. מספר גבוה יותר של מיקומי האלקטרודה (128) על הקרקפת ניתן להשתמש אם יש צורך.
- יישום הגדרות קליניות או אמבולטוריות (עם מספר אלקטרודות בלבד) אם ניתוח לוקליזציה של מקור אינו מתוכנן. השתמש במיקומים העורף כדי להקליט SSVEP, מיקומים הזמני לרכוש ASSR, ו הקודקוד מיקומים להקליט SSSEP.
- תדחוף את האלקטרודות. למחזיקי האלקטרודות ודא שהתווית של האלקטרודה תואמת לתווית המיקום בכיפה.
- ללוות את המתנדבים לחדר הניסיוני (רצוי, חדר מסוכך, מחליש קול). בקשו מהנושא לשבת על כיסא בתוך החדר, במצב נוח.
- הניחו אלקטרודות חיצוניות על האף ותנוכי האוזן אם התייחסות פיזית (שונה מאלקטרודות הקרקפת) ישמשו להפניית מחדש של הקלטת EEG (בשלב 3.8.1).
- הציבו אלקטרודות חיצוניות במיקומים הפריקולקולריות.
- מניחים אלקטרודות על הלחי ועל האזור הקדמי של הראש, כ 1 ס מ מעל הגבה, כדי להקליט מהבהב (בשלב 2.6.1).
- מניחים אלקטרודות על canthus החיצונית של העיניים, כ 1 ס מ מעל/מתחת לקו האמצע, כדי להקליט תנועות העין (בשלב 2.6.1).
הערה: electrooculogram (EOG) ישמש בשלב 3.8.5 להסרת חפצי ביצה הנגרמת על ידי תנועות מהבהב ועיניים.
- להפעיל את מערכת הרכישה EEG ולבדוק את העכבה האלקטרודה אם מערכת העכבה נמוכה משמש להקלטת EEG. תקן את העכבה, לפי הצורך, לפי כיווני היצרן. עכבה יש לשמור מתחת 10 kΩ32.
- בקש מהנושא למצמץ ולהזיז את העיניים בכיוונים שונים כדי לוודא ש-EOG מוקלט כראוי.
- כדי לנתח את הדינמיקה של SSVEP, התאם את מיקום המסך בכיוון האנכי, כדי להתאים לזווית התצוגה של הנושא. עמעם את האורות של החדר עד להשגת רמה נוחה. התאימו את רמת הזוהר של המסך לגבול העליון של רמת הנוחות ה́s של המשתתף.
- כדי לנתח את הדינמיקה של ASSR, הכנס את האוזניות בעזרת הקצף הנכון, כך שלאוזניות יתאים לתעלת האוזן. בדוק שצלילים מועברים בעוצמה הרצויה (למשל, רמה מפסיכופיסית33).
3. רכישת EEG ועיבוד מקדים
- קבעו את פרמטרי הגירוי שהוגדרו בתכנון הניסיוני. עיין במדריך למשתמש שסיפק היצרן של מערכת הגירוי לקבלת פרטים על התוכנה.
הערה: להסברים המקיפים את הגירוי המשמש ליצירת מדור SSVEP ו-ASSR, ראו Norcia et al.8 ו-ראנס34, בהתאמה. - הנחה את הנושא לשים לב לגירוי, במקרה שהמלכוד החזותי הוא נושא הניסוי.
- הציגו סרט כתוביות באמצעות צליל כבוי כאשר השמיעה מבוטלת היא הנושא של הניסוי.
הערה: הצגת סרט שקט מאפשרת הטיה של תשומת לב מגירוי אקוסטי תוך שמירה על רמת התעוררות25.
- הציגו סרט כתוביות באמצעות צליל כבוי כאשר השמיעה מבוטלת היא הנושא של הניסוי.
- ההווה גירוי ארוך יותר מ 90 s, כפי שנעשה כדי לחקור את האבולוציה בזמן של ssveps ו-assr בשני בני אדם מודלים בעלי חיים21,22,25,26.
הערה: הנוכחות הנוכחית קצרה יותר במשך הזמן אם מבוצע מחקר פיילוט. - השהה את הגירוי עבור 2 דקות אם רק תנאי ניסיוני אחד נבדק. התקשר עם הנושא כדי לבדוק את המודעות.
הערה: משך ההשהיה תלוי במשך הגירוי. השהיית 3 x יותר ממרווחי הזמן הגירוי תבטיח שתגובה העולה על ידי גירוי אחד לא תשפיע על הגירוי הקודם. השהיות ארוכות יותר מותרות אם הנושא מתבקש.- להשהות את הגירוי לפחות 10 s כאשר מצבים ניסיוניים שונים נבדקים מאז גירוי לסירוגין עם הפסקות של 10 s הוצע כדי להקטין את ההשפעות הסתגלות נוספות ולהקטין את אורך הניסוי25.
- חזור על שלבי המצגת (שלבים 3.3-3.4) לפחות 30x כדי להבטיח את SNR הגבוהה של המדידות לאחר בממוצע האפוקסי (שלב 4.4).
- הקלט את EEG באמצעות הליכים סטנדרטיים35. צור קובץ EEG נפרד עבור כל הפעלה ניסיונית.
הערה: עיין במדריך למשתמש של מערכת הרכישה לקבלת פרטים על התוכנה. - נטר את הקלטת ה-EEG כדי לזהות תקופות שינה המבוססות על רמת פעילות האלפא והתדירות שבה מופיעים פריטים מהבהבים. להשהות את הניסוי כאשר מוגברת רמות אלפא מלווה בתדרים מהבהב ירידה מזוהים, אשר מעיד על ישנוניות. דחה את ההפעלה הניסיונית מניתוח נוסף כאשר מאותרים תקופות שינה.
- חשב את השרעת של תגובת המצב היציב בתום כל הפעלה ניסויית, בעקבות ההוראה המסופקת במדריך למשתמש של תוכנת הרכישה המשמשת בניסוי.
- נטר את הרמה האטניפית של הנושא על ידי השוואת משרעת התגובות של מצב יציב שהתקבלו בסוף כל הפעלה ניסויית. להגדיר את המצב היציב משרעת שהושג ניסיוני הראשון פועל כשרעת התייחסות.
- הגדר סף דחייה (ירידה בשרעת התגובה של 5% לגבי משרעת ההתייחסות). דחה את הריצות הנסיוניות שבהן משרעת התגובה של המצב היציב עומדת בcriterium הדחייה.
- סיים את המפגש הניסיוני לאחר שרכשת את מספר הריצות המוגדרות בתכנון הניסיוני.
- טרום תהליך את נתוני EEG באופן מקוון באמצעות הליכי EEG סטנדרטיים35 מתוארים בשלבים הבאים לכיוונים של היצרן.
- הפניה מחדש להקלטה באמצעות הפניה ממוצעת (ממוצע של כל אלקטרודות ההקלטה) או ממוצע של תת-ערכה של אלקטרודות. לחלופין, השתמש בהפניה פיזית (למשל, אלקטרודות חיצוניות הנמצאות על האף והתנוכי שתוארו בשלב 2.4).
- המירו את קואורדינטות האלקטרודה למערכת 10/20 הבינלאומית אם מערכת הקואורדינטות הרדיאלי שימש במהלך רכישות EEG. עיין במדריך ליצרן לקבלת פרטים על ההמרה.
- מעבר הרצועה מסנן את אות ה-EEG בין 0.5-300 Hz. הגדר מסנן חריץ (ממורכז ב-50 Hz או 60 Hz) במידת הצורך.
- למטה-לדגום את האות EEG כדי להקטין את זמן ההוצאה להורג של האלגוריתם שנבחר להסרת חפצים בעינית (שלב 3.8.5).
הערה: תדירות דגימה של 512 Hz מספיקה לניתוח תנודות במוח בתדר מתחת 40 Hz35. - להסיר את החפצים העינית.
הערה: למטרה זו, ניתן להשתמש בטכניקות שונות (ראה אוריגיאן וגרסיה-זאפיפ35 לסקירה מקיפה על האלגוריתמים להסרת החפץ). ביניהם, ניתוח המרכיב העצמאי הוא אחד המתודולוגיות המורחבת ביותר והוא מיושם הן ניתוח מסחרי וחופשי תוכנות37,38,39. - פלח את נתוני EEG בזמן אפוקסי-נעול לגירוי. בחר את אורך האפוקסי בהתאם למטרת הניסוי.
הערה: האפוקסי צריך להיות מספיק זמן כדי לאפשר ניתוח של תגובת המצב היציב בתחום התדרים עם החלטה ספקטרלית נאותה. - אין להפעיל את האלגוריתמים הדחייה החפץ בשלב זה כדי לזהות ולהסיר את האפוקסי המכיל פריטים.
הערה: הסרת התקופה בשלב זה תגרום לשגיאות כאשר ערכת הנתונים מאורגנת כדי להפעיל את הממוצע החכם של העמודות (שלבים 4.2 ו-4.4). אלגוריתמי דחייה מיושמים בשלב עיבוד מאוחר יותר (שלב 4.1.4). - הפעל את הפונקציה DC-detrend כדי לחשב DC-מגמות באפוקסי בודדים ולתקן אותם.
- הפעל את פונקציית התיקון הבסיסית כדי לתקן את קו הבסיס של ההקלטה. בחר מרווחי זמן טרום גירוי יותר מ 200 ms.
הערה: תיקון בסיסי מורכב מחישוב ממוצע של הנתונים במרווח הזמן שנבחר. הממוצע מחושב עבור כל ערוץ ומופחתים מכל נקודת נתונים בכל תקופה.
4. חישוב המוני התגובות
- הזן את הפרמטרים הנחוצים לחישוב התגובות למצב הקבוע (איור 1A).
הערה: קוד הבית המשמש לעיבוד הנתונים זמין באופן חופשי ב-< https:/state_visually_evoked_potentials_SSVEP_elicited_in_humans_by_continuos_light_modulated_in_amplitude_at_10Hz/62573 >. עיין בטקסט העזרה בתוך הקוד לקבלת הוראות נוספות. באופן דומה, קבוצת משנה של הנתונים המשמשים במחקר זה זמין.- הזן את מספר ההקלטות (הריצות הנסיוניות) של הניסוי.
- הזן את אורך האפוקסי כדי לפלח את ההקלטות הבודדות.
- הזן את תדירות הדגימה של הניסוי.
- בחר באלגוריתם דחיית החפץ כדי לזהות ולהסיר את האפוקסי המכיל פריטים. קריטריוני הבחירה הזמינים הם 1) הדרגתי (הבדל מוחלט בין שתי דגימות רצופות), 2) מקסימום-min (ההפרש בין משרעת המקסימום והמינימלי בתקופה) ו-3) משרעת (הגברה מוחלטת ומינימלית של מקסימום).
- . תריץ את קוד העיבוד
הערה: שלבים 4.2-4.7 מתבצעים באופן אוטומטי כאשר אפשרות זו נבחרת. הפעל את השלבים באופן ידני אם הדבר מתאים.
- סדר מחדש את האפוקסי לתוך מטריצת נתונים של n שורות ו- m עמודות, שבו n מייצג את מספר ההקלטות (מסלולים ניסיוניים) ו- m מספר האפוקסי (איור 1b).
- משקל התקופה להחליש את ההשפעה של התנועה חפצים שרירים.
הערה: בעלי משקל משוקלל מתקבלים על ידי חלוקת כל דגימת מתח על-ידי שונות השרעת של התקופה שאליה הם שייכים, כך ששונות משמשת כאמצעי לשינויי משרעת וגורם שקלול40. - ממוצע עמודות בממוצע הנתונים. לשם כך, תחום הזמן ממוצע האפוקסי המתאים לאותו חלון זמן בהקלטות השונות.
הערה: שלב זה מאפשר למחשוב של משרעת המצב היציב בהקלטות עם יחס מאוד גבוה של אות לרעש (SNR). - לייצא את סדרת הזמן הנובע בממוצע לניתוח נוסף של האבולוציה הזמן של היישום בתוכנה חיצונית.
- חשב את משרעת התגובה של המצב היציב בכל תקופה הנובעת מממוצע העמודות, באמצעות התמרת פורייה מהירה (FFT).
הערה: אורך FFT אמור להתאים לאורך תקופה אחת. היישום של טכניקת הרוח אינו חובה. משרעת התגובה של מצב יציב מוגדרת כשרעת ספקטרלית המתקבלת בתדירות של אפנון השרעת של הגירויים הסנסוריים.
- חשב את משרעת התגובה של המצב היציב בכל תקופה הנובעת מממוצע העמודות, באמצעות התמרת פורייה מהירה (FFT).
- וקטור הממוצע משרעת של מספר אד הוק של FFT סלים בכל צד של תדירות התגובה כדי לחשב את רמת הרעש שיורית (RNL). מספר פחי FFT צריך להתאים לרצועת תדר של כ-3 Hz, בכל צד של תדירות התגובה.
הערה: התדר הגבוה-ספציפיות של תגובות המצב היציב הופך את משרעת התגובה ללא תלות בתנודות הרקע הללו עם תדרים דומים, שבתורו מפיצים בצורה אחידה ברצועת תדר צרה יחסית41 , 42 , 43. - התווה את השרעת של תגובת המצב היציב ושל RNL כפונקציה של אינדקס עמודות (כלומר, מספר התקופה הנרכשת) כדי לחקור את האבולוציה של תגובת המצב היציב במהלך מרווח הגירוי.
Representative Results
SSVEP היה הרוויח על ידי גירויים חזותיים רציפה של 40 s באורך, שבו עוצמת האור היה מאופנן על ידי גל sinusoidal של 10 הרץ (עומק אפנון של 90%). גירויים נמסרו על ידי ארבעה דיודות פולטות אור (נוריות) ממוקם במרכז של 50 ס מ x 50 ס מ שחור מסך, כמו קודקודים של 5 ס מ x 5 כיכר ס"מ. כאשר המשתתף ישב 70 ס מ מן המסך, האזור של ריבוע של נוריות משנה זווית חזותית של כ 4 °. מסך ה-LED תוכנן באמצעות מערכת פיתוח מיקרובקר מבוססי USB וארבע נוריות Led לבנות בהירות של 10 מ"מ קוטר. מודולציה של רוחב הפולס (PWM) שימש לשליטה באספקת החשמל המסופקת לנוריות. טכניקה זו נשלטת על עוצמות נוריות בתדר נתון וליצור את המעטפה sinusoidal הסופי. תדירות PWM של 40 kHz שימש כדי למנוע אפקט הבהוב נתנים.
שלושים הקלטות התקבלו, אשר היו מחולקים באפוקסי של 4 s. לכן, ערכת נתונים המורכבת 10 עמודות (מספר כמות EEG בתוך הקלטות) ו 30 שורות (מספר הקלטות, מספר ריצות ניסיוני) הושג.
התנודות העצביות של זמן הגירוי העצבי הפכו להיות ברור ככל שהממוצע של הטורים בוצע (איור 2). באופן משמעותי, המרווח בו נוצר SSVEP ניתן לתצפית בעקבות המתאימות לעמודה 1. בעמודה זו, 0.2 s של הראשונית הגירוי מראש מותווים בנוסף 0.8 s הראשון של entrainment העצבית. לכן, ההליך המתואר כאן מאפשר אפיון של 1) הדינמיקה של התגובה הנדנוד לאחר הקמת הסביבה העצבית כבר מוקמת ו -2) ההתחייבות של תנודות עצביות. אחד או יותר באפוקסי הוקלט לאחר סיום הגירוי יכול להיכלל גם במטריצת הנתונים כדי ללמוד הכחדה של תגובת מצב יציב לאחר היסט גירוי.
במהלך ממוצע העמודות בממוצע העמודים, משרעת המשמעות של ה-SSVEP (משרעת ספקטרלית ב-10 Hz, שחושבה על-ידי החלת הFFT) ירדה במהלך ממוצע התקופות הראשונות של העמודים ונטו לייצב לאחר מכן (איור 3A). תוצאה זו מסכימה עם מחקרים קודמים לנתח את האבולוציה של assr במהלך ממוצע של כרכים שנרכשו ברציפות21,22,40,43,44. ההתנהגות של משרעת התגובה בממוצע מוסבר בדרך כלל על ידי התרומה הגבוהה יחסית של רעש בלתי ממוצע לשרעת התגובה שחושבה האפוקסי הראשון, אשר החליש כממוצע מבוצע13, 44 , 45 , 46 , 47. לציון, השונות משרעת SSVEP ירדה באופן משמעותי ככל שהתקדמות בממוצע.
ניתחנו גם את RNL של המידות במהלך ממוצע העמודות בממוצע של אפוקסי (איור 3B). RNL החושב ברצועת תדר צרה (3 Hz) בשני צידי תדירות ה-SSVEP. למרות שהליך זה אינו נפוץ כאשר ssvep מנותח, וקטור-ממוצע מספר נתון של סלי תדירות סביב זה של ה-entrainment עצביים הוא תקן להערכת rnl מדידות assr41,42, 43. כמצופה, הירידה בהדרגה RNL כמספר האפוקסי הממוצע עלה והגיע לרמה האסימפטוטית לאחר כ-20 כמות שעובדו. שלא כמו זה נצפתה כאשר משרעת SSVEP נותחו, סטיית התקן של RNL נשאר קבוע יחסית כמו מספר התקופות בממוצע גדל, אשר מציע כי תנאי ההקלטה היו יציבים לאורך המפגש ניסיוני.
התוצאות שהוצגו לעיל קבעו את השינויים ביחס השיא לרעש (pSNR) של המידות במהלך הממוצע בעמודה של תקופה (איור 3ג). מונח זה מוגדר כאן כיחס (ב-dB) בין משרעת הכיכר של התגובה (SSVEP) ומשרעת מרובע של RNL. כאשר התקדמה בממוצע, pSNR גדל כמספר התקופות הממוצע גדל עד 18, בערך. בהפרשים נוספים במספר התקופות הממוצע לא השפיעו באופן משמעותי על איכות האות. השונות של pSNR ירדו כאפוקסי יותר הממוצע.
לבסוף, הדינמיקה של משרעת SSVEP ו-RNL מיוצגים באיור 4. הפעם התקבלו האבולוצעות על ידי התוויית פרמטרי התגובה שחושבו בסוף ממוצע העמודות בממוצע הטורים כפונקציה של מספר העמודות (כפונקציה של זמן). כפי שמתואר על ידי Labecki et al.26, הדינמיקה של SSVEP יכול להשתנות באופן משמעותי בין הנושאים. מאחר שהתוצאות המוצגות באיור 4 מתאימות לאדם אחד, לא ניתן לעשות הכללות. בנושא זה, משרעת ה-SSVEP הציגו התנהגות מורכבת יחסית (איור 4A). משרעת התגובה גדלה בהדרגה במהלך 12 השניות הראשונות לאחר התפרצות גירוי (הזמן המתאים לאורך של 3 האפוקסי). כאשר הגירוי המשיך, ה-SSVEP ירד בעקביות במהלך 12 השניות הבאות, ונשאר קבוע יחסית לאחר מכן. אין אפשרות להסביר תוצאות אלה על-ידי אופן הפעולה של RNL, מאחר שפרמטר זה היה קבוע יחסית בזמן הגירוי (איור 4B). העלייה בשרעת ה-SSVEP בעקבות התפרצות הגירוי ניכרת בעקבות המוצגים באיור 2 וניתן להסביר אותם באמצעות תהליכי אינטגרציה, הנובעים מייצוב היווצרות העצבים העצבי. הירידה הבאה בשרעת מציעה את ההסתגלות של SSVEP לגירוי מתמשך. עם זאת, השערות אלה צריכות להיבדק בניסויים מבוקרים עם מידות מדגם מופקעים.
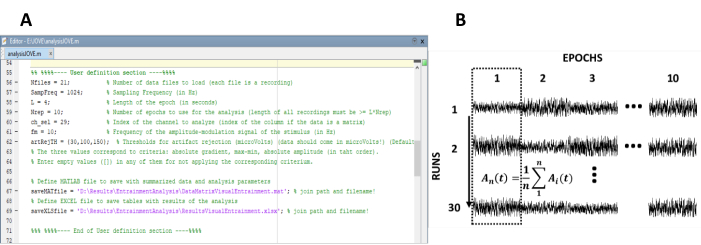
איור 1 : צעדים קריטיים לחילוץ האבולוציה זמן של משרעת של תגובות מצב יציב. (A) צילום מסך של קוד העיבוד, שבו מוגדרים פרמטרי ניתוח. (ב) מייצג את הארגון של ערכת הנתונים. מטריצת נתונים המורכבת מ-30 הקלטות של 10 כרכים מיוצגת. ממוצע העמודות של האפוקסי מודגש בעמודה הראשונה. הקו האנכי מייצג את כיוון הממוצע. אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.

איור 2 : שינויים בצורת גל של המצב היציב הפוטנציאל מעורר חזותית (SSVEP) במהלך ממוצע העמודה בממוצע של האפוקסי. תגובות הודלו על ידי מצגת רציפה של אור מאופנן משרעת ב 10 הרץ. השורות מציגות את טפסי הגל שהושגו לאחר חישוב ממוצע כל ההקלטות הקודמות (כלומר, שורה 1 היא ההקלטה הראשונה, שורה 5 היא צורת הגל המתקבלת לאחר ממוצע של חמש ההקלטות הראשונות, והשורה האחרונה היא הממוצע של כל ההקלטות). צורות גל אמינות יותר של SSVEP נצפו בכל עמודה כמו מספר בממוצע הריצות גדל. כדי לספק בהירות (להפוך את התנודות של SSVEP גלוי), רק את השנייה הראשונה של האפוקסי מיוצג. החריגים הם עקבות בעמודה הראשונה של ערכת הנתונים, שעבורן מוצגים 0.2 שניות של בסיס קדם-גירוי. אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.

איור 3 : שינויים בפרמטרי התגובה וההקלטה במהלך הממוצע החכם של העמודות. (א) האבולוציה של משרעת SSVEP. (ב) התנהגות של rnl. (ג) שינויים בpSNR. קווים שחורים מייצגים את הערכים המורעים שהושגו עבור כל עמודה (n = 10) והצל האפור מייצג את האזור המכוסה ב-± סטיית תקן אחת. אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.

איור 4 : האבולוציה הזמן של SSVEP הרוויח על ידי הצגת גירוי חזותי רציפה, מאופנן משרעת ב 10 הרץ. (A) מסלול הזמן של משרעת SSVEP. (ב) מסלול הזמן של ה-rnl. אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
Discussion
עבודה זו מתארת נוהל ניסיוני לניתוח הדינמיקה של תגובות מוחי מנדנוד. מתודולוגיה כזאת מורכבת מרכישת מספר מספיק של מריצות נסיוניות עצמאיות של אותו מצב ניסיוני, ותחום הזמן ממוצע הגוונים המתאים לאותו חלון זמן בהקלטות השונות (בממוצע עמודות ממוצע ב איור 1B). משרעת שחושבה בנתוני הממוצע מייצגים את משרעת מיידית של תגובת הנדנוד. התוויית המוני אלה כפונקציה של זמן (או מספר העמודות בערכת הנתונים) מאפשר לנתח את האבולוציה זמן של זמן תגובה מנדנוד-נעול על הגירוי. מתודולוגיה זו מהווה שינוי של ההצעה המוצעת על-ידי ריטר ואח '23 לניתוח ההסתגלות של הפוטנציאלים הבקורטיקלית הארעית. השיטה שימש כדי לנתח את הדינמיקה של פוטנציאל מעורר שמיעתי בשני בני אדם24 ודגמי בעלי חיים20,21.
מבחינה מתודולוגיים, השילוב של הפרמטרים המשמשים לעורר את התגובה מצב יציב ואלה מיושם כדי לחלץ את התגובה העצבית מרעשי הרקע הוא קריטי כדי לנתח את האבולוציה הזמן של המצב היציב הפוטנציאל המעורר 22. אורך הגירוי המשמש בניסוי המוצג כאן (40 s) נבחר בהתבסס על תוצאות שהתקבלו במחקר פיילוט. אורך זה גירוי היה מספיק כדי לנתח את ההסתגלות של assr שנוצר במוח החולדה גזע21,22. יתר על כן, אורך הגירוי צריך לחרוג מהזמן שבו הכוח הארוך אסימפטוטית של SSVEPs הוא הגיע (איור 1 ב labecki ואח '26). עם זאת, ניתן להגיע אל מעבר לשישים במקרים מסוימים (איור 2 ב לאבצקי ואח '26). לכן, הפעלת מחקר מדגם קטן מומלץ להגדיר את אורך הגירוי של הגירוי. אחרת, אורך הגירוי יותר מ 90 s מומלץ להשיג ייצוג מלאה של האבולוציה הזמן של התגובה. שימוש בהפסקות ארוכות כראוי בין הקלטות עוקבות מרמז על מסלולים ניסיוניים רציפים להיות עצמאיים מבחינה סטטיסטית (כלומר, אמצעים שונים, עצמאיים של אותו משתנה). למיטב ידיעתנו, לא נערכו ניסויים כדי לנתח את ההשהיה האופטימלית בין הרצפים (הפסקה מינימלית הנדרשת כדי לבצע רצפים עצמאיים זה מזה). הcriterium של שימוש בהפסקות לפחות 3 x יותר מאשר אורך הגירוי הוא שמרני מספיק כדי להבטיח כי התגובה מדינה יציבה שנרשמה בכל הפעלה נתונה לא מושפע הגירוי הקודם.
לאחרונה, גירויים לסירוגין (תנאים ניסיוניים) הוצע כבחירה כדי להפחית את ההשהיה בין הפעלת ניסיוני, הימנעות אפקט הסתגלות נוספת25. כמו כן, מספר הרצפים הניסיוניים (30) מיושם בפרוטוקול ניסיוני זה הוא שמרני, מאז RNL אסימפטוטיק ו pSNR מגיעים בדרך כלל לאחר בממוצע 20 ריצות ניסיוני, בערך. כאשר גירויים נופלים בתוך האזור הבינוני-עליון של הטווח הדינמי של התגובה (רמות התחושה הגבוהה), סביר להניח שמספר הרצפים הנמוכים ביותר צריך לנתח את הדינמיקה של התגובה המעוברת. אף על פי כן, במקרים שבהם נבדקים מצבים ניסיוניים שונים, מספר הריצות הניסיוני הוא חיוני לביצוע השוואות בין התנאים (כלומר, רמות תחושה שונות).
בנוסף לממוצע העמודות של האפוקסי, הדינמיקה של פוטנציאל מעורר מסובב הייתה מנותח על ידי סינון המידות החד-משפט ברצועת תדר צרה סביב תדירות הריבית והמחשוב המעטפת של הכוח אות באמצעות סינון מעבר נמוך26. כמו כן, ניתוח ניסיון יחיד יושם כדי לאפיין את תקופת המעבר הקודמת לאזור היציב של SSVEP48, ואת השינויים משרעת והשלב של ssvep במהלך האזור היציב של התגובה49. בעוד ניתוח משפטי יחיד לאפשר אפליה של תנודות מהירות יחסית בשרעת תגובה, עיצובים ניסיוניים לנתח את התגובה הממוצעת בלוקים מופרדים עם מרווח נתון בין לחסום רק לבין וריאציות לטווח ארוך ב משרעת של הפוטנציאל מעורר50,51. ממוצע העמודים של האפוקסי עומד בין שתי אפשרויות אלה. המרת האות הממוצע לתדר-תחום באמצעות FFT מרמז על ניתוח הדינמיקה של התגובה עם רזולוציה שווה לאורך התקופה. בדוגמה שהוצגה כאן, ה-SSVEP דווח על כל 4 ס'. למרות שהרזולוציה של 4 s מספיקה לתאר את הדינמיקה המתרחשת במרווחי זמן של עשרות שניות, כגון זה של ssvep26, האפוקסי החופף חלקית בהקלטות המקוריות מאפשר לתאר את התפתחות הזמן של ה תגובה מדינה יציבה באופן מעודן יותר25.
הדינמיקה של התגובות מצב יציב המתקבל לאחר ממוצע העמודה בממוצע של האפוקסי מייצגים בעיקר האבולוציה של פעילות מנדנוד המסונכרן בין פלחי EEG הממוצע (אלה אשר לשרוד את הממוצע). לפיכך, בעיה מרכזית בנוגע לכדאיות של המתודולוגיה היא הנחת התגובה האפשרית עקב וריאציות בשלב התנודות העצביות של מרוץ ניסיוני עצמאי אחד למשנהו (כלומר, בין ההקלטות). נושא זה צריך להיות ממוען לניסויים. עם זאת, ראיות מעידות על כך ששלב התגובות במוח הוא פחות משתנה מהצפוי. למעשה, מספר מחקרים דיווחו על סדירות בשלב הצפוי של האדם 80 Hz assr47,48,49. כאשר השהיות מוערכות בהתאם לשלב הפעילות הנדנוד, ההשפעה הצפויה של העוצמה ותדר המנשא של הגירויים האקוסטיים על השהיית תגובות השמיעה נצפתה (כלומר, ירידה בהשהיה כ עוצמה ועלייה בתדר המוביל)52,53,54. יתר על כן, שינויים מורזיים טיפוסיים משרעת והשמאל-לימין סימטריה ברמות השמיעה נצפו גם כאשר השהיות מוערך משלב של assr47,55,56 , 57 , 58. כאשר מתארים את האבולוציה הזמן של ssvep באמצעות ניתוח חד פעמי, labecki ואח '26 ציין כי למרות שינויים בין משפט של הגברה התגובה בתוך אותו נושא היה גבוה במידה ניכרת, השונות של ה שלב היה פחות מבוטא באופן משמעותי.
בהתבסס על התצפיות שלהם, Labecki ואח '26 הציע כי מינימום של 50 מבחנים צריך להיות ממוצע כדי לקבל שערוך אמין של מעטפת כוח ממוצע של התגובה. תוצאות אלה מצביעות על כך, שגם כאשר משרעת התגובה מחושבת בניסויים בודדים, יש צורך בחישוב ממוצע (של מעטפות במקרה זה) כדי לדווח על תוצאות אמינות. יתרה מזאת, השונות הבין-משפט בשרעת ה-SSVEP שדווחה על-ידי לאבצקי ואח '26 מצביעה על כך שהמיחשוב של פרמטר זה בניסויים בודדים יכול להיות מושפע מאוד מרעשי הרקע. בהתחשב באבולוציה של יחס אות לרעש המוצג באיור 2, מיחשוב התגובה באות בממוצע במקום ניסויים בודדים מפחית באופן משמעותי את מספר פלחי EEG הדרושים לעיבוד כדי לקבל אמין דידות. בנוסף, השונות הנמוכה בשלב שהושג על-ידי לאבצקי ואח '26 תומכת ברעיון שממוצע העמודים של האפוקסי המוצג כאן הוא הליך חוקי למחשוב את הדינמיקה של הפוטנציאל המעורר.
חישוב ממוצע הנתונים ברמות שונות מוביל לפרשנות שונה של התוצאות. בנוגע לפוטנציאלים מסובבים, מחשב את משרעת התגובה לאחר שהתחום הזמני של הריצות העצמאיות מרמז על ניתוח תנודות בזמן בלבד (קרי, אלה ששרדו את הממוצע). הליך זה עשוי לסנן מידע רלבנטי לגבי הדינמיקה של התגובה בניסויים בודדים. עם זאת, הוא מבטיח יחס מספיק גבוה לרעש של המידות. היבט זה עשוי להיות משמעותי כאשר התגובות קרובות לסף האלקטרולוגי, מצב שבו ניתן להתפשר על זיהוי של התגובה בגלל היחס של אות לרעש נמוך של המדידה.
Disclosures
. למחברים אין מה לגלות
Acknowledgments
המחברים מכירים בהכרת תודה לוסיה Zepeda, גרייס א. ויטאקר וניקולא ניוטו על תרומתם להפקת וידאו. עבודה זו נתמכת בחלק על ידי תוכניות CONICYT בסיס FB0008, MEC 80170124 ו PhD מלגה 21171741, כמו גם המכון הלאומי על חירשות ובעיות תקשורת אחרות של המוסדות הלאומיים לבריאות תחת מספר הפרס P50DC015446. התוכן הינו באחריות המחברים בלבד ואינו מייצג בהכרח את ההשקפות הרשמיות של המכון הלאומי לבריאות.
Materials
| Name | Company | Catalog Number | Comments |
| Active electrodes | Biosemi | P32-1020-32ACMS (ABC) | for channels 1-32 |
| Active electrodes | Biosemi | P32-1020-32A (ABC) | for channels 33-64 |
| Active electrodes | Biosemi | 8 x TP FLAT | external electrodes |
| Active-Two adquisition system | Biosemi | version 7.0 | EEG adquisition system |
| alcohol | Salcobrand | Code: 3309011 | for cleaning the scalp |
| Electrode cap 64 channels | Biosemi | CAP MS xx yy | cap |
| Electrode cap 64 channels | Biosemi | CAP ML xx yy | cap |
| gel | Biosemi | SIGNA BOX12 | conductive gel |
| Laptop | Asus | Core i7 1TB DD + 128GB SSD 8GB RAM | computer for stimulation |
| Laptop | Asus | Core i7 1TB DD + 128GB SSD 8GB RAM | computer for recording |
| LED screen | in-house production | - | The screen consists of four light-emitting diodes (LEDs) situated on the center of a 50x50 cm black screen, as vertexes of a square of 5x5 cm |
| sterile gauze | Salcobrand | Code: 8730277 | for cleaning the scalp |
References
- Berger, H. Über das elektrenkephalogramm des menschen. European Archives of Psychiatry and Clinical Neuroscience. 87 (1), 527-570 (1929).
- Berger, H. Electroencephalogram of humans. Journal fur Psychologie und Neurologie. 40, 160-179 (1930).
- Walter, W. G. The location of cerebral tumours by electro-encephalography. The Lancet. 228 (5893), 305-308 (1936).
- Jasper, H. H., Andrews, H. L. Electro-encephalography: III. Normal differentiation of occipital and precentral regions in man. Archives of Neurology & Psychiatry. 39 (1), 96-115 (1938).
- Snyder, A. Z. Steady-state vibration evoked potentials: descriptions of technique and characterization of responses. Electroencephalography and Clinical Neurophysiology. 84 (3), 257-268 (1992).
- Giabbiconi, C. M., Trujillo-Barreto, N. J., Gruber, T., Muller, M. M. Sustained spatial attention to vibration is mediated in primary somatosensory cortex. Neuroimage. 35, 255-262 (2007).
- Rossion, B., Boremanse, A. Robust sensitivity to facial identity in the right human occipito-temporal cortex as revealed by steady-state visual-evoked potentials. Journal of Vision. 11 (2), (2011).
- Norcia, A. M., Appelbaum, L. G., Ales, J. M., Cottereau, B. R., Rossion, B. The steady-state visual evoked potential in vision research: a review. Journal of Vision. 15 (6), 4 (2015).
- Galambos, R., Makeig, S., Talmachoff, P. J. A 40-Hz auditory potential recorded from the human scalp. Proceedings of the National Academy of Sciences of the United States of America. 78 (4), 2643-2647 (1981).
- Picton, T. W., Skinner, C. R., Champagne, S. C., Kellett, A. J., Maiste, A. C. Potentials evoked by the sinusoidal modulation of the amplitude or frequency of a tone. The Journal of the Acoustical Society of America. 82 (1), 165-178 (1987).
- Dawson, G. D. A summation technique for the detection of small evoked potentials. Electroencephalography and Clinical Neurophysiology. 6 (1), 65-84 (1954).
- Savio, G., Cardenas, J., Abalo, M. P., Gonzalez, A., Valdes, J. The low and high frequency auditory steady state responses mature at different rates. Audiology and Neurotology. 6 (5), 279-287 (2001).
- Luts, H., Desloovere, C., Kumar, A., Vandermeersch, E., Wouters, J. Objective assessment of frequencyspecific hearing thresholds in babies. International Journal of Pediatric Otorhinolaryngology. 68, 915-926 (2004).
- Valdes, J. L., et al. Comparison of statistical indicators for the automatic detection of 80 Hz auditory steady state responses. Ear and Hearing. 18 (5), 420-429 (1997).
- Wilding, T., McKay, C., Baker, R., Kluk, K. Auditory steady state responses in normal-hearing and hearing-impaired adults: an analysis of between-session amplitude and latency repeatability, test time, and F ratio detection paradigms. Ear and Hearing. 33 (2), 267 (2012).
- de Resende, L. M., et al. Auditory steady-state responses in school-aged children: a pilot study. Journal of NeuroEngineering and Rehabilitation. 12 (1), 13 (2015).
- Halbleib, A., et al. Topographic analysis of engagement and disengagement of neural oscillators in photic driving: a combined electroencephalogram/magnetoencephalogram study. Journal of Clinical Neurophysiology. 29 (1), 33-41 (2012).
- Salchow, C., et al. Rod Driven Frequency Entrainment and Resonance Phenomena. Frontiers in Human Neuroscience. 10, 413 (2016).
- Wacker, M., et al. A time-variant processing approach for the analysis of alpha and gamma MEG oscillations during flicker stimulus generated entrainment. IEEE Transactions on Biomedical Engineering. 58 (11), 3069-3077 (2011).
- Glaser, E. M., Ruchkin, D. S. Principles of Neurobiological Signal Analysis. Journal of Clinical Engineering. 2 (4), 382-383 (1977).
- Prado-Gutierrez, P., et al. Habituation of auditory steady state responses evoked by amplitude-modulated acoustic signals in rats. Audiology Research. 5 (1), (2015).
- Prado-Gutierrez, P., Martínez-Montes, E., Weinstein, A., Zañartu, M. Estimation of auditory steady-state responses based on the averaging of independent EEG epochs. PLoS ONE. 14 (1), (2019).
- Ritter, W., Vaughan, H. G. Jr, Costa, L. D. Orienting and habituation to auditory stimuli: a study of short terms changes in average evoked responses. Electroencephalography and Clinical Neurophysiology. 25 (6), 550-556 (1968).
- Malmierca, M. S., Cristaudo, S., Pérez-González, D., Covey, E. Stimulus-specific adaptation in the inferior colliculus of the anesthetized rat. Journal of Neuroscience. 29 (17), 5483-5493 (2009).
- Van Eeckhoutte, M., Luke, R., Wouters, J., Francart, T. Stability of auditory steady state responses over time. Ear and Hearing. 39 (2), 260-268 (2018).
- Labecki, M., Nowicka, M. M., Suffczynski, P. Temporal Modulation of Steady-State Visual Evoked Potentials. International Journal of Neural Systems. 29 (3), 1850050 (2019).
- Zhang, S., et al. A study on dynamic model of steady-state visual evoked potentials. Journal of Neural Engineering. 15 (4), 046010 (2018).
- Wilson, U. S., Kaf, W. A., Danesh, A. A., Lichtenhan, J. T. Assessment of low-frequency hearing with narrowband chirp evoked 40-Hz sinusoidal auditory steady state response. International Journal of Audiology. 55 (4), 239-247 (2016).
- Kaf, W. A., Mohamed, E. S., Elshafiey, H. 40-Hz Sinusoidal Auditory Steady-State Response and Tone Burst Auditory Brainstem Response Using a Kalman Filter to Determine Thresholds Pre- and Post-Myringotomy with Grommet Tube in Children With Mild, Low-Frequency Conductive Hearing Loss. American Journal of Audiology. 25 (1), 41-53 (2016).
- Luke, R., Wouters, J. Kalman filter based estimation of auditory steady state response parameters. IEEE Transactions on Neural Systems and Rehabilitation Engineering. 25, 196-204 (2017).
- Homan, R. W. The 10-20 Electrode System and Cerebral Location. American Journal of EEG Technology. 28 (4), 269-279 (1988).
- Kappenman, E. S., Steven, J. L. The Effects of Electrode Impedance on Data Quality and Statistical Significance in ERP Recordings. Psychophysiology. 47 (5), 888-904 (2010).
- Moore, R., Gordon-Hickey, S., Jones, A. Most comfortable listening levels, background noise levels, and acceptable noise levels for children and adults with normal hearing. Journal of the American Academy of Audiology. 22 (5), 286-293 (2011).
- Rance, G. The Auditory Steady-state Response: Generation, Recording, and Clinical Application. Plural Publishing. 335, (2008).
- Luck, S. J. An introduction to the event-related potential technique. Second Edition. , MIT Press. 406 (2014).
- Urigüen, J. A., Garcia-Zapirain, B. EEG artifact removal-state-of-the-art and guidelines. Journal of Neural Engineering. 12 (3), (2015).
- Jung, T. P., et al. Removing electroencephalographic artifacts by blind source separation. Psychophysiology. 37 (2), 163-178 (2000).
- Li, Y., Ma, Z., Lu, W., Li, Y. Automatic removal of the eye blink artifact from EEG using an ICA-based template matching approach. Physiological Measurement. 27 (4), 425 (2006).
- Mannan, M. M. N., Jeong, M. Y., Kamran, M. A. Hybrid ICA-Regression: automatic identification and removal of ocular artifacts from electroencephalographic signals. Frontiers in Human Neuroscience. 10, 193 (2016).
- John, M. S., Dimitrijevic, A., Picton, T. W. Weighted averaging of steady-state responses. Clinical Neurophysiology. 112 (3), 555-562 (2001).
- Zurek, P. M. Detectability of transient and sinusoidal otoacoustic emissions. Ear and Hearing. 13 (5), 307-310 (1992).
- Lins, O. G., Picton, P. E., Picton, T. W., Champagne, S. C., Durieux-Smith, A. Auditory steady-state responses to tones amplitude-modulated at 80-110 Hz. Journal of the Acoustical Society of America. 97 (5), 3051-3063 (1995).
- John, M. S., Picton, T. W. Human auditory steady-state responses to amplitude-modulated tones: phase and latency measurements. Hearing Research. 141 (1-2), 57-79 (2000).
- Fortuny, A. T., et al. Criterio de parada de la promediación de la respuesta auditiva de estado estable. Acta Otorrinolaringológica Española. 62 (3), 173-254 (2011).
- John, M. S., Purcell, D. W., Dimitrijevic, A., Picton, T. W. Advantages and caveats when recording steady-state responses to multiple simultaneous stimuli. Journal of the American Academy of Audiology. 13 (5), 246-259 (2002).
- Luts, H., Van Dun, B., Alaerts, J., Wouters, J. The influence of the detection paradigm in recording auditory steady-state responses. Ear and Hearing. 29 (4), 638-650 (2008).
- Choi, J. M., Purcell, D. W., John, M. S. Phase stability of auditory steady state responses in newborn infants. Ear and Hearing. 32 (5), 593-604 (2011).
- Xu, M., et al. Use of a steady-state baseline to address evoked vs. oscillation models of visual evoked potential origin. Neuroimage. 134, 204-212 (2016).
- Peachey, N. S., Demarco, P. J. Jr, Ubilluz, R., Yee, W. Short-term changes in the response characteristics of the human visual evoked potential. Vision Research. 34 (21), 2823-2831 (1994).
- Woods, D. L., Elmasian, R. The habituation of event-related potentials to speech soundsand tones. Electroencephalography and Clinical Neurophysiology. 65, 447-459 (1986).
- Ravden, D., Polich, J. Habituation of P300 from visual stimuli. International Journal of Psychophysiology. 30 (3), 359-365 (1998).
- Picton, T. W., Dimitrijevic, A., John, M. S., Van Roon, P. The use of phase in the detection of auditory steady-state responses. Clinical Neurophysiology. 112 (9), 1698-1711 (2001).
- Alaerts, J., Luts, H., Van Dun, B., Desloovere, C., Wouters, J. Latencies of auditory steady-state responses recorded in early infancy. Audiology and Neurotology. 15 (2), 116-127 (2010).
- John, M. S., Brown, D. K., Muir, P. J., Picton, T. W. Recording auditory steady-state responses in young infants. Ear and Hearing. 25 (6), 539-553 (2004).
- Purcell, D. W., John, S. M., Schneider, B. A., Picton, T. W. Human temporal auditory acuity as assessed by envelope following responses. Journal of the Acoustical Society of America. 116 (6), 3581-3593 (2004).
- Kuwada, S., et al. Sources of the scalp-recorded amplitude-modulation following response. Journal of the American Academy of Audiology. 13 (4), 188-204 (2002).
- Pauli-Magnus, D., et al. Detection and differentiation of sensorineural hearing loss in mice using auditory steady-state responses and transient auditory brainstem responses. Neuroscience. 149 (3), 673-684 (2007).
- Prado-Gutierrez, P., et al. Maturational time course of the Envelope Following Response to amplitude-modulated acoustic signals in rats. International Journal of Audiology. 51 (4), 309-316 (2012).
