

Summary
外部反復刺激に対する神経の巻き上げた時間の進化を評価するプロトコルが提示される。同じ実験条件の定常状態記録が取得され、時間領域で平均化されます。定常状態ダイナミクスは、応答振幅を時間の関数としてプロットすることによって分析されます。
Abstract
神経の巻き上がりは、感覚刺激の周期性に対する神経活動の同期を指す。この同期は、定常状態の誘発応答(すなわち、駆動刺激に対する脳波位相ロックにおける振動)の生成を定義する。定常状態誘発応答の振幅の古典的な解釈は、刺激の繰り返しの提示を平均化するような、ステレオタイプ的な時間不変性神経応答とランダムな背景変動を仮定する。ステレオタイプな応答。このアプローチは、刺激への長期暴露によって引き起こされた適応の場合のように、定常状態のダイナミクスを無視します。定常応答のダイナミクスを分析するために、応答振幅の時間の進化は、十分に長い休憩によって分離された異なる刺激実行において同じであると仮定することができる。この仮定に基づいて、定常応答の時間進化を特徴付ける方法が提示される。同じ実験条件に応じて十分に多くの記録が得られる。実験実行 (録画) は列ごとの平均値です (つまり、実行は平均されますが、記録内のエポックは前のセグメントで平均化されません)。カラムワイズ平均化により、信号対雑音比が著しく高い録音における定常応答の分析が可能になります。したがって、平均信号は定常応答の時間進化を正確に表し、時間と周波数の両方の領域で分析することができる。本研究では、応答の一例として、定常状態の視覚的に誘発された電位を用いて、方法の詳細な説明が提供される。利点と注意事項は、神経の巻き込みを分析するように設計された単一試験方法との比較に基づいて評価されます。
Introduction
頭皮から記録されると、脳の電気的活動は、時間の経過とともに電圧の連続的かつ定期的な変化として観察されます。この電気活動は脳波計(EEG)と呼ばれ、前世紀1の20年代後半にハンス・ベルガーによって最初に説明されました。その後の精液研究は、異なるリズミカルまたは反復パターンが2、3、4を観察することができる複合時系列としてEEGを記述した。今日では、脳波は、通常、異なる感覚および認知プロセスに関連付けられている5つの確立された周波数帯域、デルタ、シータ、アルファ、ベータ、およびガンマに分けられます。
長年にわたり、脳波を用いた脳振動の研究は、進行中の活動におけるスペクトルの分析または非周期的感覚事象によって引き起こされた振動活性の変化のいずれかに制限されていた。過去数十年の間に、進行中の脳波振動を調節し、そのような変調が知覚および認知プロセスに及ぼす影響を探る方法論が、リズミカルな感覚刺激の提示を含めて実施されてきた。神経の巻き込みを誘発する。神経エントレインメントという用語は、感覚刺激の周期的特性との神経活動の同期を指す。このプロセスは、定常状態の誘発電位(すなわち、駆動刺激の周期的特性にロックされた脳波振動)の生成につながります。定常状態誘発電位は、一定の速度で提示される一過性刺激または目的の頻度で振幅で変調された連続刺激のいずれかを使用して、視覚、聴覚、およびバイブロタクタイル刺激によって最も一般的に引き起こされます。身体感覚安定状態誘発電位(SSセプ)は反復的な触覚刺激5、6、定常状態の視覚的に誘発された電位(SSVEP)に応答して記録されるのに対し、一般に周期的に引き起こされる輝きのちらつき、写真、および顔7、8のプレゼンテーション。聴覚定常応答(ASSR)は、通常、一過性音響刺激の列車または振幅変調トーン9、10の連続的な提示によって生成される。
測定された脳波からの定常状態誘発電位の抽出は、本質的に、刺激11に時間ロックされた後に獲得されたEEGエポックの平均化に依存する。応答の周期性により、時間および頻度の両方の領域で分析できます。周波数領域変換後、感覚応答は、外部刺激の提示速度または変調周波数、およびそれに対応する高調波の振幅のピークとして観察される。これらの手順(時間領域平均化およびその後の周波数領域変換)は、臨床目的12、13、14を用いたASSR法の検出に基づく聴力検査を開発するために不可欠であった。 、15、16.
さらに、EEGエポックの古典的な時間領域平均化は、SSVEP17,18の生成および絶滅などの生理学的プロセスを分析するのに非常に有用であった。ちらつきの連続した列車を提示し、記録内のその後のエポックを平均化し、Wacker et al.19は、SSVEPの位相ロック指数が刺激の最初の400ミリ秒の間に急速に増加し、その後も高いままであることを観察しました。.彼らはまた、刺激発症後700-1 100ミリ秒の間に堅牢な視覚の巻き上げが確立されたことを報告しました。ある程度の巻き込みは、振動応答17、19の約3期間続いた刺激列車のオフセット後に有効であった。これらの挙動は、観察された振動の係合/離脱効果として解釈されてきたが、これは、人間の視覚系17における非線形情報処理の結果である。あるいは、特定の実験条件下では、ちらつき刺激は、最初にオン応答を引き出すことができることが知られており、神経エントレインメント18の代わりに刺激列車の終わりにオフ応答が得られる。
平均連続的に獲得された脳波エポックへの主な仮定は、脳波信号が感覚応答とバックグラウンドノイズ20の線形組み合わせを表す。さらに、振動応答の振幅、周波数、位相は静止していると仮定し、バックグラウンドノイズはランダムな活動と見なされます。しかし、この仮定が満たされない場合、複数のエポックの後に計算される応答振幅は、必ずしも誘発電位の瞬間振幅に対応するとは限らない。
ラットの脳幹で生成されたASSRは、振幅変調トーンの連続的な発光に適応することが最近報告されている(すなわち、応答振幅は時間の経過とともに指数関数的に減少する)21,22.適応は、単調に反復的な感覚刺激の新規性の喪失を反映する神経機構として解釈され、音響環境23,24における関連する変動に対する感受性を高める。聴覚経路では、適応は騒々しい環境での音声理解を高めるかもしれない。さらに、このプロセスは、音声制作を制御するために、自分の声の聴覚フィードバックを監視するための既存のメカニズムの一部である可能性があります。
ヒトにおける40Hz ASSRの時間進化を分析したVan Eeckhoutte etal.25は、時間の経過に伴う応答振幅の有意な減少を観察した(時間の経過とともに線形減少を仮定した場合、グループ分析に基づく-0.0002 μV/s程度)。その結果、これらの著者らは、ヒトにおける40Hz ASSRは刺激に適応しないと結論付けた。ヒトでは、SSVEP26の安定性を分析する際に非静止挙動が観察されている。これらの著者は、SSVEPの基本周波数と第2高調波の振幅が、それぞれ試験した被験者のわずか30%と66.7%で静止していることを観察した。両方のSSVEP周波数成分の相は、時間の経過とともに比較的安定しているが、小さなドリフト26を示した。
したがって、その後獲得されたエポックの古典的な時間領域の平均化は、神経のエントレインメントの静止特性の探索を可能にするが、この方法論は、エントレインメントの長期的なダイナミクスが焦点である場合に改訂する必要がある。長期ダイナミクスの発生によって短期ダイナミクスの平均化が破損した場合。定常状態応答の非定常動作を特徴付けるには、特定の時間枠で計算された呼び出された応答は、前述の EEG セグメントで計算された応答によって危険にさらされるべきではありません。言い換えれば、呼び出された電位は、前のEEGセグメントで時間領域平均化されることなく、バックグラウンドノイズから抽出する必要があります。
本研究では,神経の巻き込みのダイナミクスを評価する方法を紹介する.定常状態応答は、同じ刺激に応じて繰り返し記録され、連続した記録は実験実行の3倍の休止間隔によってインターリーブされる。生理学的応答の時間の進化が異なる独立した実験実行(独立した記録)で同じである場合、録音は列方向に平均化されます。つまり、異なるレコーディング内の同じ場所に対応するエポックは、録音内のエポックを平均化せずに平均化されます。その結果、任意の刺激間隔で計算される応答振幅は、誘発された電位の瞬間的な振幅に対応します。感覚応答は、実験の目的に応じて、時間領域で分析するか、周波数ドメインに変換することができます。いずれの場合も、振幅は定常応答の時間進化を分析する時間の関数としてプロットできます。定常状態の誘発電位の生成と絶滅は、分析を録音の最初と最後のエポックに制限することによって評価することができる。
ニューラルエントレインメントのダイナミクスは、狭帯域フィルタリング単一試験測定を対象範囲に絞り込み、ローパスフィルタリング25を使用して電力信号のエンベロープを計算するなど、他のアプローチを使用して分析することができます。ヒルバート変換27.これらの方法論と比較して、エポックの列ごとの平均化により、信号対雑音比(SNR)が高い信号に基づいて定常状態パラメータを計算できます。最近、カルマンフィルタリングは、40Hz ASSR振幅28、29、30の推定のための有望な技術として出現している。カルマンフィルタリングの実装は、電気生理学的閾値に近い定常状態応答の検出を改善し、聴力検査29の時間を短縮することができる。さらに、カルマン フィルタリングアプローチを使用して ASSR 振幅30を推定する場合は、静止応答を想定する必要はありません。それにもかかわらず、カルマンフィルタリング25を使用してASSRの時間進化を分析した研究は1つしかありません。研究の結論は、40Hz ASSR振幅が刺激間隔にわたって安定している。したがって、カルマンフィルタリングは、ASRRが静止していない条件でテストする必要があります。
時間はかかりますが、列方向の平均化方法はモデルフリーであり、ノイズ動作の初期化値や事前定義は必要ありません。さらに、収束時間を伴わないため、列方向の平均化は、神経エントレインメントの発症のより信頼性の高い表現を提供してもよい。したがって、カラムワイズ平均法で得られた結果は、カルマンフィルタリングを用いて神経エントレインメントのダイナミクスを解析するための根拠となる真理と考えることができる。
このプロトコルの説明は、SSVEP の例に基づいています。ただし、ここで示す方法はモダリティに依存しないものであり、SSSEP および ASSR の時間進化の分析にも使用できることに注意してください。
Protocol
本研究は、チリ・バルパライソ大学の研究倫理委員会(評価声明コードCEC170-18)の承認を得て実施し、ヒト被験者との研究に関する国家ガイドラインに確認された。
1. 準備
- 件名を歓迎します。
- 研究の目的と関連性を説明する。関連する技術的な詳細の説明を提供します。すべての質問に徹底的に答えます。
- 必要に応じて、いつでも実験セッションを中断できることを明示的に言及します。
- ボランティアに対して、件名に関する同意書を読み、対応するフォームに署名してもらいます。インフォームド コンセントが取得されない場合は、実験セッションを中断します。
2. 被験者の準備
- 被験者に快適な位置で実験室の椅子に座るように頼む。
- エタノールで頭皮をきれいにする(95%の溶液)死んだ皮膚細胞の層とそれを覆う皮脂を除去する。このステップは、電極と頭皮の間のインピーダンスを低減するために重要です。
- 測定テープでヘッド円周を測定し、使用する電極キャップのサイズを定義します。
- 電極キャップを着用するように被験者に依頼します。キャップの快適で正しい位置決めの指示を提供します。
- 鼻(Nz、鼻の上の目と鼻の上の間のくぼみによって識別することができる鼻前部縫合糸の中間点)とイオン(Iz、後頭骨の顕著)との間の距離を測定テープを使用して測定します。
- 測定テープを使用して、左右の前耳のポイント(耳の耳の前のくぼみとして識別される)間の距離を測定します。
- 電極キャップの位置を補正し、前の手順で定義した架空の線の交点がヘッドの頂点に対応するようにします。調整後、被写体が快適であることを確認します。
- 実験のために考慮される場所に応じて、電極ホルダーに導電性ゲルを入れてください。
- インターナショナル 10-20 システム31に続く 64 の頭皮の位置を使用して、プロトコルの結果を使用してソースローカリゼーション分析を実行します。必要に応じて、頭皮上の電極位置(128)の数が多いほど使用できます。
- ソースローカリゼーション解析が計画されていない場合は、臨床または歩行設定(少数の電極のみ)を実装します。後頭部の場所を使用して、SSVEP、ASSR を取得する時間的な場所、および SSSEP を記録する頭頂部の場所を記録します。
- 電極ホルダーの電極を押します。電極のラベルがキャップ内の位置のラベルと一致していることを確認します。
- ボランティアを実験室に同行する(好ましくは、遮蔽された、音減衰室)。快適な位置で、部屋の中の椅子に座るように被験者に尋ねます。
- 物理的な参照(頭皮電極とは異なる)がEEG記録を再参照するために使用される場合は、鼻と耳たぶに外部電極を置きます(ステップ3.8.1)。
- 外電極を円形の場所に配置します。
- 頬と頭部の前頭部の前頭部に電極を置き、眉の上に約1cm上に、点滅を記録します(ステップ2.6.1)。
- 目の外側のカンサスに電極を置き、中線の上下約1cmで目の動きを記録します(ステップ2.6.1)。
注:電極図(EOG)は、点滅と目の動きによって誘発されるEGGアーティファクトを除去するためのステップ3.8.5で使用されます。
- EEG取得システムをオンにし、低インピーダンスシステムを使用してEEGを記録する場合は、電極インピーダンスを確認します。製造元の指示に応じて、必要に応じてインピーダンスを修正します。インピーダンスは10kΩ32以下にしてください。
- 被写体にまばたきをし、異なる方向に目を動かして、EOGが正しく記録されていることを確認します。
- SSVEP のダイナミックを解析するには、被写体の視野角に合わせて画面の位置を垂直方向に調整します。快適なレベルが達成されるまで、部屋のライトを暗くします。画面の輝度レベルを参加者の快適度の上限に調整します。
- ASSRのダイナミクスを分析するには、正しいフォームインサートを使用してイヤホンを挿入し、イヤホンが外耳道にフィットするようにします。音が所望の強度(例えば、心理物理学の快適なレベル33)で送達されることを確認してください。
3. 脳波の取得と前処理
- 実験計画で定義された刺激パラメータを設定します。ソフトウェアの詳細については、刺激システムの製造元が提供する取扱説明書を参照してください。
注:SSVEPおよびASSRの生成に使用される刺激の包括的な説明については、それぞれNorcia et al.8およびRance34を参照してください。 - 刺激に注意を払うように被験者に指示し、視覚のエントレインが実験の話題である場合に。
- 聴覚の巻き上げが実験の話題である場合は、音をオフにして字幕付きムービーを提示します。
注:サイレントムービーのプレゼンテーションは、覚醒レベル25を維持しながら、音響刺激からの注意の偏向を可能にします。
- 聴覚の巻き上げが実験の話題である場合は、音をオフにして字幕付きムービーを提示します。
- 現在の刺激は90sを超え、ヒトおよび動物モデル21、22、25、26の両方におけるSSVEPおよびASSRの時間進化を調査するために行われている。
注:パイロット調査が行われた場合、持続時間が短い刺激を提示します。 - 1 つの実験条件のみがテストされている場合は、刺激を 2 分間一時停止します。被験者と対話して意識を確認します。
注: 一時停止の継続時間は、刺激の継続時間によって異なります。刺激間隔より3倍長く一時停止すると、1つの刺激によって引き起こされた応答が前の刺激の影響を受けないようにします。サブジェクトが要求する場合は、より長い一時停止が許可されます。- 10sの一時停止と交互刺激が提案されているので、異なる実験条件が試験されたときに少なくとも10sの刺激を一時停止し、余分な適応効果を減少させ、実験25の長さを減らすことが提案されている。
- エポックの平均化後の測定値の高い SNR を確認するために、プレゼンテーション手順 (手順 3.3~3.4) を少なくとも 30 倍繰り返します(ステップ 4.4)。
- 標準的な手順35を使用して脳波を記録する。実験実行ごとに個別の EEG ファイルを作成します。
注: ソフトウェアの詳細については、取得システムの取扱説明書を参照してください。 - 脳波記録を監視して、アルファアクティビティのレベルと点滅するアーティファクトが表示される頻度に基づいて睡眠期間を検出します。点滅周波数の低下を伴うアルファレベルの増加が検出された場合、実験を一時停止します。睡眠期間が検出されたときに、さらなる分析から実験実行を拒否します。
- 実験実行の終了時に定常状態応答の振幅を計算し、実験で使用される取得ソフトウェアの取扱説明書に記載されている指示に従って計算する。
- 各実験実行の最後に得られた定常応答の振幅を比較することにより、被験者の注意レベルを監視します。最初の実験実行で得られた定常状態振幅を基準振幅として設定します。
- 拒絶しきい値を設定します (基準振幅に関する応答振幅の 5% の減少)。定常状態応答の振幅が拒絶基準を満たす実験実行を拒否します。
- 実験計画で定義された実行数を取得した後、実験セッションを終了します。
- 製造元の指示に従って次の手順で説明されている標準的な脳波手順35を使用して、EEG データをオフラインで事前処理します。
- 平均参照(すべての記録電極の平均)または電極のサブセットの平均を使用して記録を再参照します。あるいは、物理的参照を使用する(例えば、ステップ2.4に記載の鼻および耳たぶに置かれた外部電極)。
- 脳波集録中に放射状座標系を使用した場合は、電極座標を国際10/20系に変換します。変換の詳細については、製造元のマニュアルを参照してください。
- バンドパスフィルタ 0.5~300 Hz の間のEEG信号をフィルタリングする 必要に応じてノッチフィルタ(50 Hzまたは60Hzを中心に)を設定します。
- EEG 信号をダウンサンプリングして、眼のアーティファクトを除去するために選択したアルゴリズムの実行時間を短縮します(ステップ 3.8.5)。
注:512 Hzのサンプリング周波数は、40 Hz35以下の周波数の脳振動を分析するのに十分です。 - 眼のアーティファクトを取り除く。
注:この目的のために、異なる技術を使用することができます(アーティファクト除去アルゴリズムの広範なレビューについては、ウリギュエンとガルシアザピレイン35を参照してください)。その中で、独立した成分分析は、最も拡張された方法論の1つであり、商用および自由分析ソフトウェア37、38、39の両方で実装されている。 - 刺激にタイムロックされたエポックでEEGデータをセグメント化します。実験の目的に応じてエポックの長さを選択します。
注: エポックは、十分なスペクトル分解能を持つ周波数領域における定常状態応答の分析を可能にするのに十分な長さである必要があります。 - アーティファクトを含むエポックを検出して削除するには、この段階でアーティファクト拒否アルゴリズムを実行しないでください。
注: この段階でエポックを削除すると、データセットがエポックの列ごとの平均化を実行するように編成されたときにエラーが発生します (手順 4.2 および 4.4)。拒否アルゴリズムは、後の処理ステップ (ステップ 4.1.4) で実装されます。 - DC-デトレンド関数を実行して、個々のEEGエポックのDCトレンドを計算し、それらを修正します。
- ベースライン補正機能を実行して、記録のベースラインを修正します。200 ミリ秒より長い事前刺激時間間隔を選択します。
注: ベースライン修正は、選択した時間間隔内のデータの平均化で構成されます。平均は各チャネルについて計算され、各エポックの各データポイントから減算されます。
4. 応答振幅の計算
- 定常状態応答の計算に必要なパラメータを入力します (図 1A)。
注: データの処理に使用される社内コードは、- 実験の録画数 (実験実行) を入力します。
- 個々の録音をセグメント化するエポックの長さを入力します。
- 実験のサンプリング頻度を入力します。
- アーティファクトを含むエポックを検出して削除するには、アーティファクト拒否アルゴリズムを選択します。使用可能な選択基準は、1)グラデーション(2つの連続したサンプル間の絶対差)、2)最大最小(エポックの最大振幅と最小振幅の差)、および3)振幅(絶対最大および最小振幅)です。
- 処理コードを実行します。
注: このオプションを選択すると、手順 4.2-4.7 が自動的に実行されます。必要に応じて、手順を手動で実行します。
- n 行とm列のデータ 行列にエポックを再配置し、n は記録の数 (実験実行) とmのエポック数を表します (図 1B)。
- エポックを重み付けして、モーションと筋肉のアーティファクトの効果を減衰させます。
注:加重EEGエポックは、各電圧サンプルをそれらが属するエポックの振幅分散で割ることによって得られ、分散は振幅変動および重み係数40の尺度として使用される。 - 列ごとのデータセットの平均値。この目的のために、時間ドメインは、異なる録音の同じタイムウィンドウに対応するエポックを平均化します。
注: このステップでは、信号対雑音比(SNR)が著しく高い記録における定常振幅の計算が可能になります。 - 外部ソフトウェアのエントレインメントの時間進化をさらに分析するために、平均化に起因する時系列をエクスポートします。
- 高速フーリエ変換(FFT)を使用して、列ごとの平均化に起因する各エポックの定常状態応答の振幅を計算します。
注: FFT の長さは、1 つのエポックの長さに対応する必要があります。ウィンドウ化手法の実装は必須ではありません。定常応答の振幅は、感覚刺激の振幅変調の周波数で得られるスペクトル振幅として定義される。
- 高速フーリエ変換(FFT)を使用して、列ごとの平均化に起因する各エポックの定常状態応答の振幅を計算します。
- ベクトルは、応答の周波数の両側にあるFFTビンのアドホック数の振幅を平均して、残留ノイズレベル(RNL)を計算します。FFTビンの数は、応答の周波数の両側で、約3Hzの周波数帯域に対応する必要があります。
注:定常応答の高周波特異性は、応答振幅を同様の周波数でバックグラウンド振動とは無関係にし、比較的狭い周波数帯域41に均一に分配する,42歳,43. - 定常状態応答とRNLの振幅を列インデックスの関数としてプロットし(すなわち、後天的なエポックの数)、刺激間隔中の定常状態応答の進化を探索する。
Representative Results
SSVEPは、長さ40sの連続的な視覚刺激によって引き起こされ、光強度は10Hz(変調深さ90%)の前弦波によって変調された。刺激は、50 cm x 50 cm の黒い画面の中央に位置する 4 つの発光ダイオード (LED) によって、5 cm x 5 cm の正方形の頂点として送達されました。参加者が画面から70cmに座ると、LEDの正方形の面積は約4°の視覚角度を下塗りする。LEDスクリーンは、USBベースのマイクロコントローラ開発システムと直径10mmの超明るい白色LED4種を使用して設計されました。パルス幅変調(PWM)技術を使用して、LEDに供給される電力を制御しました。この手法は、特定の周波数で LED 強度を制御し、最終的な洞帯エンベロープを生成します。40 kHzのPWM周波数を使用して、知覚可能なちらつき効果を回避しました。
30の録音が得られ、これは4sのエポックでセグメント化された。そこで、10列(記録内のEEGエポック数)と30行(記録数、実験実行回数)で構成されるデータセットを取得した。
刺激に対する神経振動時間ロックは、列ワイズ平均化が行われたので明らかになった(図2)。重要なことに、SSVEPが生成される間隔は、列1に対応するトレースで観察することができる。その列では、神経エントレインメントの最初の 0.8 s に加えて、0.2 s の事前刺激ベースラインがプロットされます。したがって、ここで説明する手順は、1)神経のエントレインメントが既に確立された後の振動応答のダイナミクスを特徴付け、2)神経振動の関与を可能にする。刺激の終了後に記録された1つ以上のエポックは、刺激オフセット後の定常状態応答の絶滅を研究するためにデータマトリックスに含めることもできる。
エポックのカラムワイズ平均化の間、SSVEPの平均振幅(10Hzでスペクトル振幅、FFTを適用して計算)は、カラムの最初のエポックの平均化中に減少し、その後安定する傾向があった(図3A)。この結果は、順次獲得されたエポック21、22、40、43、44の平均化中のASSRの進化を分析する以前の研究と一致する。平均化中の応答振幅の挙動は、通常、平均化が行われるように減衰される最初のエポックで計算された応答振幅に対する平均化されていないノイズの比較的高い寄与によって説明される。44歳,45歳,46歳,47.注目すべきは、平均化が進むにつれてSSVEPの振幅変動が著しく減少した点である。
また、エポックのカラムワイズ平均化時の測定値のRNLを分析しました(図3B)。RNLは、SSVEPの周波数の両側で狭い周波数帯域(3Hz)で計算された。この手順は、SSVEPを分析する際には一般的ではありませんが、神経エントレインメントの周りの所定の周波数ビンの数をベクトル平均化することは、ASSR測定値41,42でRNLを推定するための標準であり、 43. 予想通り、平均エポックの数が増加するにつれてRNLは徐々に減少し、約20のエポックが処理された後に無症床レベルに達した。SSVEP振幅を解析した際に観測されたものとは異なり、平均エポック数が増加するにつれてRNLの標準偏差は比較的一定であり、実験セッションに沿って記録条件が安定していることを示唆している。
上で提示された結果は、エポックのカラムワイズ平均化中の測定値のピーク信号対雑音比(pSNR)の変化を決定した(図3C)。この用語は、応答の平方振幅(SSVEP)とRNLの正方形振幅との比率(dB)としてここで定義される。平均化が進むにつれて、pSNRは平均エポックの数が18まで増加するにつれて増加しました。平均エポックの数のさらなる増分は、信号の品質に大きな影響を与えなかった。pSNRの変動性は、より多くのエポックが平均されるにつれて減少した。
最後に、SSVEP 振幅と RNL のダイナミクスを図4に表します。これらの時間の進化は、列数の関数として(時間の関数として)エポックのカラムワイズ平均の最後に計算された応答パラメータをプロットすることによって得られました。Labecki et al.26によって示されるように、SSVEPのダイナミクスは被験者間で有意に異なる可能性がある。図 4に示す結果は単一の個人に対応するため、一般化はできません。この主題では、SSVEPの振幅は比較的複雑な挙動を示した(図4A)。応答振幅は、刺激発症後の最初の12秒間に徐々に増加した(3エポックの長さに相当する時間)。刺激が持続するにつれて、SSVEPは次の12秒間一貫して減少し、その後は比較的一定のままでした。このパラメータは刺激間隔中に比較的一定であったため、これらの結果はRNLの挙動では説明できません(図4B)。刺激発症後のSSVEP振幅の増加は、図2に示された痕跡に明らかであり、統合プロセスによって説明することができ、神経の巻き込みの安定化をもたらす。振幅のその後の減少は、持続的な刺激へのSSVEPの適応を示唆する。それにもかかわらず、これらの仮説は、適切なサンプルサイズを持つ制御された実験でテストする必要があります。
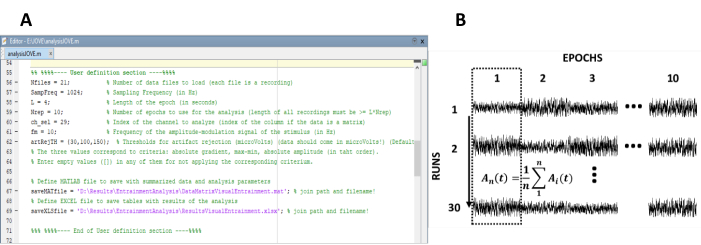
図 1: 定常応答の振幅の時間進化を抽出するための重要なステップ。(A) 分析パラメータが定義されている処理コードのスクリーンショット。(B) データセットの編成を示す代表的な図。10エポックの30の記録から成るデータマトリックスが表される。エポックの列ごとの平均化は、最初の列で強調表示されます。垂直線は、平均化の方向を表します。この図のより大きなバージョンを表示するには、ここをクリックしてください。

図 2: エポックのカラムワイズ平均化における定常状態の波形の変化を視覚的に誘発する電位(SSVEP)。応答は、10 Hzで振幅で変調された光の連続的な提示によって引き起こされた。行は、すべての以前の記録を平均した後に得られた波形を示します(つまり、行 1 は最初の記録、行 5 は最初の 5 つの記録を平均した後に取得された波形であり、最後の行はすべての記録の平均です)。平均実行回数が増加するにつれて、各列で SSVEP のより信頼性の高い波形が観察されました。(SSVEP の振動を表示するために)わかりやすくするために、エポックの最初の 1 秒だけが表されます。例外は、データ・セットの最初の列のトレースで、0.2 秒の事前刺激ベースラインが表示されます。この図のより大きなバージョンを表示するには、ここをクリックしてください。

図 3: エポックの列ごとの平均化中の応答パラメータと記録パラメータの変更。(A) SSVEP振幅の進化。(B) RNL の動作。(C) pSNR の変更。黒い線は各列(n = 10)で得られた平均値を表し、灰色の影は±1標準偏差でカバーされる領域を表します。この図のより大きなバージョンを表示するには、ここをクリックしてください。

図 4:連続的な視覚刺激の提示によって引き起こされたSSVEPの時間進化は、10 Hzで振幅で変調した。(A) SSVEP振幅のタイムコース。(B) RNLのタイムコース。この図のより大きなバージョンを表示するには、ここをクリックしてください。
Discussion
本研究では、振動脳応答のダイナミクスを解析するための実験的手順について述べている。このような方法論は、同じ実験条件の十分な数の独立した実験実行を獲得し、異なる記録における同じ時間枠に対応する時間ドメイン平均エポック(列ワイズ平均化) 図1B)。平均データで計算される振幅は、振動応答の瞬間的な振幅を表します。これらの振幅を時間の関数(またはデータセット内の列数)としてプロットすると、刺激に対する時間ロックされた振動応答の時間の進化を分析できます。この方法論は、一過性皮質誘発電位の適応を分析するためにRitter et al.23によって提案されたものの修正である。この方法は、ヒト24および動物モデル20、21の両方における聴覚誘発電位の動的性を分析するために使用されてきた。
方法論的観点から、定常状態応答を引き出すために使用されるパラメータと、バックグラウンドノイズから神経応答を抽出するために実装されたパラメータの組み合わせは、定常状態誘発電位の時間進化を分析するために重要です。22.ここで提示した実験で用いられる刺激長(40s)を、パイロット試験で得られた結果に基づいて選択した。この刺激長は、ラット脳幹21,22で生成されたASSRの適応を分析するのに十分であった。さらに、刺激の長さは、SSVEPの無症性瞬間帯域パワーに達した時間を超えるべきである(図1 labecki et al.26)。それにもかかわらず、SSVEPの無症の瞬間的なバンドパワーは、場合によっては60を超えて到達することができます(図2 labecki et al.26)。したがって、刺激の刺激長を定義するために、小サンプルパイロットスタディを実行することをお勧めします。それ以外の場合は、応答の時間進化の完全な表現を達成するために、90sより長い刺激長を推奨する。連続した記録間で十分に長い休止を使用すると、連続した実験実行を統計的に独立している(すなわち、同じ変数の異なる独立した尺量)と考える必要があります。私たちの知るうまで、実行間の最適な一時停止(実行を互いに独立させるために必要な最小休止)を分析するための実験は行われていません。刺激の長さよりも少なくとも3倍長い休止を使用する基準は、任意の実行で記録された定常状態応答が前の刺激の影響を受けないことを保証するのに十分保守的である。
近年、実験実行間の休止を低減する選択肢として交互刺激(実験条件)が提案されており、余分な適応効果25を回避する。同様に、この実験プロトコルで実装された実験実行数(30)は、通常、20回の実験実行を平均した後に無症間RNLおよびpSNRに達するため、保守的である。刺激が応答の動的範囲の中間上の領域内に収まるとき(高感覚レベル)、誘発された応答のダイナミクスを分析するために実行の数が少なくなる可能性が高い。それにもかかわらず、異なる実験条件がテストされる場合には、同じ数の実験実行を持つことは、条件間の比較(すなわち、異なる感覚レベル)を行う上で重要です。
エポックのカラムワイズ平均化に加えて、振動誘発電位のダイナミクスは、関心のある周波数の周りの狭い周波数帯域で単一試算測定をフィルタリングし、電力のエンベロープを計算することによって分析されています。ローパスフィルタリング26を使用した信号。同様に、単一試験分析は、SSVEP48の安定領域に先行する遷移期間、および応答49の安定領域におけるSSVEPの振幅および位相の変化を特徴付けるために実施された。単一の試験分析は、応答振幅の比較的速い変動の判別を可能にするが、実験的な計画は、特定のブロック間間隔で区切られたブロック内の平均応答を分析する唯一の長期変動を考慮する誘発された電位50、51の振幅。エポックの列ごとの平均化は、これら 2 つのオプションの間にあります。FFT を使用して平均信号を周波数ドメインに変換すると、応答のダイナミクスがエポックの長さと等しい解像度で解析されます。ここで示す例では、SSVEP は 4 s ごとに報告されています。4秒の解像度は、SSVEP26のような数十秒を超える時間間隔で起こるダイナミクスを記述するのに十分であるが、元の録音のエポックの一部が重複し、時間の進化を記述することができます。より洗練された方法で定常状態応答25.
エポックのカラムワイズ平均化後に得られた定常応答のダイナミクスは、主に平均的な脳波セグメント間で同期される振動活性の進化を表します(平均化を生き残るもの)。したがって、方法論の実現可能性に関する主要な問題は、ある独立した実験実行から別の(すなわち、記録の間)に神経振動の相の変動による応答振幅の減衰の可能性である。このトピックは実験的に対処する必要があります。しかし、証拠は、脳振動応答の相が予想よりも変動が少ないことを示しています。実際には、いくつかの研究は、ヒト80 Hz ASSR 47、48、49の予想される段階で規則性を報告している。振動活動の相に基づいて待ち時間を推定すると、聴覚応答の待ち時間に対する音響刺激の強度およびキャリア周波数の予測可能な影響が観察されている(すなわち、遅延が減少する)強度とキャリア周波数増加)52,53,54.さらに、ASSR47、55、56の段階から遅延を推定した場合、聴力レベルの振幅と左右の非対称性の典型的な合理変化も観察されている。,57歳,58.単一試験分析を用いてSSVEPの時間進化を記述する場合、Labecki et al.26は、同じ被験者内の応答振幅の試算間変動がかなり高いが、フェーズの顕著なほど顕著ではなかった。
彼らの観察に基づいて、Labeckiらは、応答の平均パワーエンベロープの信頼性の高い推定を得るために、最低50の試験を平均化する必要があることを示唆しました。これらの結果は、応答の振幅が単一の試行で計算された場合でも、信頼できる結果を報告するために(その場合のエンベロープの)平均化が必要であることを示しています。さらに、Labeckiら26によって報告されたSSVEPの振幅における試験間変動は、単一試験におけるこのパラメータの計算がバックグラウンドノイズの影響を強く受けることができることを示唆している。図2に示す信号対雑音比の進化を考慮すると、単一の試験ではなく平均信号における応答の計算は、信頼性を得るために処理に必要なEEGセグメントの数を大幅に減少させます。測定。さらに、Labecki et al.26によって得られた相の低変動は、ここで提示されるエポックのカラムワイズ平均化が、振動誘発電位のダイナミクスを計算するための有効な手順であるという考えを支持する。
異なるレベルでデータを平均すると、結果の解釈が異なります。振動誘発電位に関しては、独立した実行の時間ドメイン平均化後の応答振幅の計算は、時間ロックされた振動(すなわち、平均化を生き残るもの)のみを分析することを意味します。この手順は、個々の試験における応答のダイナミクスに関する関連情報をフィルタリングする場合があります。しかし、それは測定の十分に高い信号対雑音比を保証する。この態様は、応答が電気生理学的閾値に近い場合に重要であり得る。
Disclosures
著者は何も開示していない。
Acknowledgments
著者たちは、ルシア・ゼペダ、グレース・A・ウィテカー、ニコラス・ニートがビデオ制作に貢献したことを感謝しています。この研究は、CONICYTプログラムBASAL FB0008、MEC 80170124および博士課程奨学金21171741、および国立衛生研究所の難聴およびその他のコミュニケーション障害に関する国立研究所によって一部支援されました P50DC015446.内容は著者の責任のみであり、必ずしも国立衛生研究所の公式見解を表すものではありません。
Materials
| Name | Company | Catalog Number | Comments |
| Active electrodes | Biosemi | P32-1020-32ACMS (ABC) | for channels 1-32 |
| Active electrodes | Biosemi | P32-1020-32A (ABC) | for channels 33-64 |
| Active electrodes | Biosemi | 8 x TP FLAT | external electrodes |
| Active-Two adquisition system | Biosemi | version 7.0 | EEG adquisition system |
| alcohol | Salcobrand | Code: 3309011 | for cleaning the scalp |
| Electrode cap 64 channels | Biosemi | CAP MS xx yy | cap |
| Electrode cap 64 channels | Biosemi | CAP ML xx yy | cap |
| gel | Biosemi | SIGNA BOX12 | conductive gel |
| Laptop | Asus | Core i7 1TB DD + 128GB SSD 8GB RAM | computer for stimulation |
| Laptop | Asus | Core i7 1TB DD + 128GB SSD 8GB RAM | computer for recording |
| LED screen | in-house production | - | The screen consists of four light-emitting diodes (LEDs) situated on the center of a 50x50 cm black screen, as vertexes of a square of 5x5 cm |
| sterile gauze | Salcobrand | Code: 8730277 | for cleaning the scalp |
References
- Berger, H. Über das elektrenkephalogramm des menschen. European Archives of Psychiatry and Clinical Neuroscience. 87 (1), 527-570 (1929).
- Berger, H. Electroencephalogram of humans. Journal fur Psychologie und Neurologie. 40, 160-179 (1930).
- Walter, W. G. The location of cerebral tumours by electro-encephalography. The Lancet. 228 (5893), 305-308 (1936).
- Jasper, H. H., Andrews, H. L. Electro-encephalography: III. Normal differentiation of occipital and precentral regions in man. Archives of Neurology & Psychiatry. 39 (1), 96-115 (1938).
- Snyder, A. Z. Steady-state vibration evoked potentials: descriptions of technique and characterization of responses. Electroencephalography and Clinical Neurophysiology. 84 (3), 257-268 (1992).
- Giabbiconi, C. M., Trujillo-Barreto, N. J., Gruber, T., Muller, M. M. Sustained spatial attention to vibration is mediated in primary somatosensory cortex. Neuroimage. 35, 255-262 (2007).
- Rossion, B., Boremanse, A. Robust sensitivity to facial identity in the right human occipito-temporal cortex as revealed by steady-state visual-evoked potentials. Journal of Vision. 11 (2), (2011).
- Norcia, A. M., Appelbaum, L. G., Ales, J. M., Cottereau, B. R., Rossion, B. The steady-state visual evoked potential in vision research: a review. Journal of Vision. 15 (6), 4 (2015).
- Galambos, R., Makeig, S., Talmachoff, P. J. A 40-Hz auditory potential recorded from the human scalp. Proceedings of the National Academy of Sciences of the United States of America. 78 (4), 2643-2647 (1981).
- Picton, T. W., Skinner, C. R., Champagne, S. C., Kellett, A. J., Maiste, A. C. Potentials evoked by the sinusoidal modulation of the amplitude or frequency of a tone. The Journal of the Acoustical Society of America. 82 (1), 165-178 (1987).
- Dawson, G. D. A summation technique for the detection of small evoked potentials. Electroencephalography and Clinical Neurophysiology. 6 (1), 65-84 (1954).
- Savio, G., Cardenas, J., Abalo, M. P., Gonzalez, A., Valdes, J. The low and high frequency auditory steady state responses mature at different rates. Audiology and Neurotology. 6 (5), 279-287 (2001).
- Luts, H., Desloovere, C., Kumar, A., Vandermeersch, E., Wouters, J. Objective assessment of frequencyspecific hearing thresholds in babies. International Journal of Pediatric Otorhinolaryngology. 68, 915-926 (2004).
- Valdes, J. L., et al. Comparison of statistical indicators for the automatic detection of 80 Hz auditory steady state responses. Ear and Hearing. 18 (5), 420-429 (1997).
- Wilding, T., McKay, C., Baker, R., Kluk, K. Auditory steady state responses in normal-hearing and hearing-impaired adults: an analysis of between-session amplitude and latency repeatability, test time, and F ratio detection paradigms. Ear and Hearing. 33 (2), 267 (2012).
- de Resende, L. M., et al. Auditory steady-state responses in school-aged children: a pilot study. Journal of NeuroEngineering and Rehabilitation. 12 (1), 13 (2015).
- Halbleib, A., et al. Topographic analysis of engagement and disengagement of neural oscillators in photic driving: a combined electroencephalogram/magnetoencephalogram study. Journal of Clinical Neurophysiology. 29 (1), 33-41 (2012).
- Salchow, C., et al. Rod Driven Frequency Entrainment and Resonance Phenomena. Frontiers in Human Neuroscience. 10, 413 (2016).
- Wacker, M., et al. A time-variant processing approach for the analysis of alpha and gamma MEG oscillations during flicker stimulus generated entrainment. IEEE Transactions on Biomedical Engineering. 58 (11), 3069-3077 (2011).
- Glaser, E. M., Ruchkin, D. S. Principles of Neurobiological Signal Analysis. Journal of Clinical Engineering. 2 (4), 382-383 (1977).
- Prado-Gutierrez, P., et al. Habituation of auditory steady state responses evoked by amplitude-modulated acoustic signals in rats. Audiology Research. 5 (1), (2015).
- Prado-Gutierrez, P., Martínez-Montes, E., Weinstein, A., Zañartu, M. Estimation of auditory steady-state responses based on the averaging of independent EEG epochs. PLoS ONE. 14 (1), (2019).
- Ritter, W., Vaughan, H. G. Jr, Costa, L. D. Orienting and habituation to auditory stimuli: a study of short terms changes in average evoked responses. Electroencephalography and Clinical Neurophysiology. 25 (6), 550-556 (1968).
- Malmierca, M. S., Cristaudo, S., Pérez-González, D., Covey, E. Stimulus-specific adaptation in the inferior colliculus of the anesthetized rat. Journal of Neuroscience. 29 (17), 5483-5493 (2009).
- Van Eeckhoutte, M., Luke, R., Wouters, J., Francart, T. Stability of auditory steady state responses over time. Ear and Hearing. 39 (2), 260-268 (2018).
- Labecki, M., Nowicka, M. M., Suffczynski, P. Temporal Modulation of Steady-State Visual Evoked Potentials. International Journal of Neural Systems. 29 (3), 1850050 (2019).
- Zhang, S., et al. A study on dynamic model of steady-state visual evoked potentials. Journal of Neural Engineering. 15 (4), 046010 (2018).
- Wilson, U. S., Kaf, W. A., Danesh, A. A., Lichtenhan, J. T. Assessment of low-frequency hearing with narrowband chirp evoked 40-Hz sinusoidal auditory steady state response. International Journal of Audiology. 55 (4), 239-247 (2016).
- Kaf, W. A., Mohamed, E. S., Elshafiey, H. 40-Hz Sinusoidal Auditory Steady-State Response and Tone Burst Auditory Brainstem Response Using a Kalman Filter to Determine Thresholds Pre- and Post-Myringotomy with Grommet Tube in Children With Mild, Low-Frequency Conductive Hearing Loss. American Journal of Audiology. 25 (1), 41-53 (2016).
- Luke, R., Wouters, J. Kalman filter based estimation of auditory steady state response parameters. IEEE Transactions on Neural Systems and Rehabilitation Engineering. 25, 196-204 (2017).
- Homan, R. W. The 10-20 Electrode System and Cerebral Location. American Journal of EEG Technology. 28 (4), 269-279 (1988).
- Kappenman, E. S., Steven, J. L. The Effects of Electrode Impedance on Data Quality and Statistical Significance in ERP Recordings. Psychophysiology. 47 (5), 888-904 (2010).
- Moore, R., Gordon-Hickey, S., Jones, A. Most comfortable listening levels, background noise levels, and acceptable noise levels for children and adults with normal hearing. Journal of the American Academy of Audiology. 22 (5), 286-293 (2011).
- Rance, G. The Auditory Steady-state Response: Generation, Recording, and Clinical Application. Plural Publishing. 335, (2008).
- Luck, S. J. An introduction to the event-related potential technique. Second Edition. , MIT Press. 406 (2014).
- Urigüen, J. A., Garcia-Zapirain, B. EEG artifact removal-state-of-the-art and guidelines. Journal of Neural Engineering. 12 (3), (2015).
- Jung, T. P., et al. Removing electroencephalographic artifacts by blind source separation. Psychophysiology. 37 (2), 163-178 (2000).
- Li, Y., Ma, Z., Lu, W., Li, Y. Automatic removal of the eye blink artifact from EEG using an ICA-based template matching approach. Physiological Measurement. 27 (4), 425 (2006).
- Mannan, M. M. N., Jeong, M. Y., Kamran, M. A. Hybrid ICA-Regression: automatic identification and removal of ocular artifacts from electroencephalographic signals. Frontiers in Human Neuroscience. 10, 193 (2016).
- John, M. S., Dimitrijevic, A., Picton, T. W. Weighted averaging of steady-state responses. Clinical Neurophysiology. 112 (3), 555-562 (2001).
- Zurek, P. M. Detectability of transient and sinusoidal otoacoustic emissions. Ear and Hearing. 13 (5), 307-310 (1992).
- Lins, O. G., Picton, P. E., Picton, T. W., Champagne, S. C., Durieux-Smith, A. Auditory steady-state responses to tones amplitude-modulated at 80-110 Hz. Journal of the Acoustical Society of America. 97 (5), 3051-3063 (1995).
- John, M. S., Picton, T. W. Human auditory steady-state responses to amplitude-modulated tones: phase and latency measurements. Hearing Research. 141 (1-2), 57-79 (2000).
- Fortuny, A. T., et al. Criterio de parada de la promediación de la respuesta auditiva de estado estable. Acta Otorrinolaringológica Española. 62 (3), 173-254 (2011).
- John, M. S., Purcell, D. W., Dimitrijevic, A., Picton, T. W. Advantages and caveats when recording steady-state responses to multiple simultaneous stimuli. Journal of the American Academy of Audiology. 13 (5), 246-259 (2002).
- Luts, H., Van Dun, B., Alaerts, J., Wouters, J. The influence of the detection paradigm in recording auditory steady-state responses. Ear and Hearing. 29 (4), 638-650 (2008).
- Choi, J. M., Purcell, D. W., John, M. S. Phase stability of auditory steady state responses in newborn infants. Ear and Hearing. 32 (5), 593-604 (2011).
- Xu, M., et al. Use of a steady-state baseline to address evoked vs. oscillation models of visual evoked potential origin. Neuroimage. 134, 204-212 (2016).
- Peachey, N. S., Demarco, P. J. Jr, Ubilluz, R., Yee, W. Short-term changes in the response characteristics of the human visual evoked potential. Vision Research. 34 (21), 2823-2831 (1994).
- Woods, D. L., Elmasian, R. The habituation of event-related potentials to speech soundsand tones. Electroencephalography and Clinical Neurophysiology. 65, 447-459 (1986).
- Ravden, D., Polich, J. Habituation of P300 from visual stimuli. International Journal of Psychophysiology. 30 (3), 359-365 (1998).
- Picton, T. W., Dimitrijevic, A., John, M. S., Van Roon, P. The use of phase in the detection of auditory steady-state responses. Clinical Neurophysiology. 112 (9), 1698-1711 (2001).
- Alaerts, J., Luts, H., Van Dun, B., Desloovere, C., Wouters, J. Latencies of auditory steady-state responses recorded in early infancy. Audiology and Neurotology. 15 (2), 116-127 (2010).
- John, M. S., Brown, D. K., Muir, P. J., Picton, T. W. Recording auditory steady-state responses in young infants. Ear and Hearing. 25 (6), 539-553 (2004).
- Purcell, D. W., John, S. M., Schneider, B. A., Picton, T. W. Human temporal auditory acuity as assessed by envelope following responses. Journal of the Acoustical Society of America. 116 (6), 3581-3593 (2004).
- Kuwada, S., et al. Sources of the scalp-recorded amplitude-modulation following response. Journal of the American Academy of Audiology. 13 (4), 188-204 (2002).
- Pauli-Magnus, D., et al. Detection and differentiation of sensorineural hearing loss in mice using auditory steady-state responses and transient auditory brainstem responses. Neuroscience. 149 (3), 673-684 (2007).
- Prado-Gutierrez, P., et al. Maturational time course of the Envelope Following Response to amplitude-modulated acoustic signals in rats. International Journal of Audiology. 51 (4), 309-316 (2012).
