

Summary
En protokol til vurdering af tids udviklingen af neurale medrivning til eksterne gentagne stimuli præsenteres. Steady-State optagelser af samme eksperimentelle tilstand erhverves og gennemsnit i tids domænet. Steady-State dynamik analyseres ved at afbilde respons amplitude som en funktion af tid.
Abstract
Neurale medrivning refererer til synkroniseringen af neurale aktivitet til hyppigheden af sensoriske stimuli. Denne synkronisering definerer genereringen af Steady-State fremkaldte svar (dvs. svingninger i elektro encephalogram fase-låst til drivende stimuli). Den klassiske fortolkning af amplituden af Steady-State fremkaldte svar antager en stereotyp tid-invariant neurale respons plus tilfældige baggrunds udsving, således at gennemsnit over gentagne præsentationer af stimulus genindvinder stereotype respons. Denne fremgangsmåde ignorerer dynamikken i Steady-State, som i tilfælde af tilpasningen fremkaldt af langvarige eksponeringer mod stimulus. For at analysere dynamikken i Steady-State svar, kan det antages, at tids udviklingen af respons amplitude er den samme i forskellige stimulerings kørsler adskilt af tilstrækkeligt lange pauser. Baseret på denne antagelse, en metode til at karakterisere den tid udviklingen af Steady-State svar er præsenteret. Et tilstrækkeligt stort antal optagelser erhverves som svar på samme eksperimentelle tilstand. Eksperimenterende kørsler (optagelser) er kolonne vise gennemsnit (dvs., at kørsler er gennemsnits, men Epoch inden for optagelser beregnes ikke i forhold til de foregående segmenter). Den søjle vise gennemsnit tillader analyse af Steady-State svar i optagelser med bemærkelsesværdigt høje signal-til-støj nøgletal. Derfor er det gennemsnitligt signal giver en nøjagtig gengivelse af tids udviklingen i Steady-State respons, som kan analyseres i både tid og frekvens domæner. I denne undersøgelse, en detaljeret beskrivelse af metoden er tilvejebragt, ved hjælp af Steady-State visuelt fremkaldte potentialer som et eksempel på et svar. Fordele og forbehold evalueres baseret på en sammenligning med enkelt-Trial metoder designet til at analysere neurale entrainment.
Introduction
Når den optages fra hovedbunden, observeres hjernens elektriske aktivitet som kontinuerlige og regelmæssige ændringer i spændinger over tid. Denne elektriske aktivitet kaldes elektro encephalogram (EEG) og blev først beskrevet af hans Berger i slutningen af tyverne af sidste århundrede1. Efterfølgende skelsættende undersøgelser beskrev EEG som en sammensat tidsserie, hvor forskellige rytmiske eller gentagne mønstre kan observeres2,3,4. I dag er EEG typisk opdelt i fem veletablerede frekvensbånd, Delta, theta, alpha, beta, og gamma, som er forbundet med de forskellige sensoriske og kognitive proces.
I årevis, studiet af hjernen svingninger ved hjælp af EEG var begrænset til enten analyse af spektret i den igangværende aktivitet eller ændringer i oscillerende aktivitet fremkaldt af ikke-periodiske sensoriske hændelser. I de seneste årtier er forskellige metoder blevet implementeret for at moduere igangværende EEG-svingninger og undersøge virkningerne af sådanne modulationer på perceptuelle og kognitive processer, herunder præsentationen af rytmisk sensorisk stimulation for inducerende neurale entrainment. Udtrykket neurale medrivning refererer til synkroniseringen af neurale aktivitet med de periodiske egenskaber af sensoriske stimuli. Denne proces fører til generering af Steady-State fremkaldte potentialer (dvs. EEG svingninger låst til de periodiske egenskaber af drivende stimuli). Steady-State fremkaldte potentialer er mest udbredt ved visuel, auditiv og vibrotaktile stimulation, ved hjælp af enten forbigående stimuli præsenteret med en konstant hastighed eller kontinuerlig stimulation moduleret i amplitude på hyppigheden af interesse. Der henviser til, at somatosensoriske Steady State fremkaldte potentialer (sssep'er) registreres som reaktion på gentagen taktile stimulation5,6, Steady-State visuelt fremkaldte potentialer (ssveps) er generelt fremkaldt af den periodiske præsentation af luminans flimrer, billeder og ansigter7,8. Auditive Steady-State respons (assrs) genereres normalt af tog af forbigående akustiske stimuli eller ved kontinuerlig præsentation af amplitude-modulerede toner9,10.
Udvinding af Steady-State fremkaldte potentialer fra den målte EEG hovedsagelig er afhængig af gennemsnit efterfølgende erhvervet EEG epoker tid-låst til stimulus11. På grund af hyppigheden af svarene, kan de analyseres i både tid og frekvens domæner. Efter frekvens-domæne transformation, den sensoriske respons er observeret som toppe af amplitude ved præsentationen sats eller modulation frekvens af de eksterne stimuli, og deres tilsvarende harmoniske. Disse procedurer (tid-domæne gennemsnit og den efterfølgende frekvens-domæne omdannelse) har været afgørende for at udvikle en høretest baseret på påvisning af ASSR-metoder med kliniske formål12,13,14 ,15,16.
Desuden har den klassiske tid-domæne gennemsnit af EEG epoker har været yderst nyttig til at analysere fysiologiske processer såsom generering og udryddelse af ssvep17,18. Ved at præsentere fortløbende tog med flimmer og gennemsnitligt efterfølgende epoker i en optagelse bemærkede Wacker et al.19 , at det fase låsende indeks for ssvep hurtigt steg i løbet af de første 400 MS stimulation og forblev højt bagefter . De rapporterede også, at robust visuel medrivning blev etableret mellem 700-1 100 ms efter stimulus debut. En vis grad af indtrædelse forblev effektiv efter offset af stimulerings toget, som varede ca. tre perioder af den oscillatoriske respons17,19. Disse adfærd er blevet fortolket som engagerende/frakobling effekten af de observerede svingninger, som er en konsekvens af den ikke-lineære informationsbehandling i det menneskelige visuelle system17. Alternativt, det er kendt, at under visse eksperimentelle betingelser, flimmer stimulation kan fremkalde on-svar i begyndelsen, og off-svar i slutningen af stimulation tog i stedet for neurale medrivning18.
Den vigtigste antagelse til gennemsnitlige fortløbende erhvervede EEG-epoker er, at EEG-signalet repræsenterer en lineær kombination af sensorisk respons og baggrundsstøjen20. Desuden antages amplituden, hyppigheden og fase af det oscillatoriske respons at være stationært, hvorimod baggrundsstøjen betragtes som en tilfældig aktivitet. Men i tilfælde, hvor denne antagelse ikke er opfyldt, svarer den respons amplitude, der er beregnet efter flere epoker, ikke nødvendigvis til den øjeblikkelige amplitude af det fremkaldte potentiale.
Det er for nylig blevet rapporteret, at ASSR genereret i hjernestammen af rotter tilpasser sig den kontinuerlige præsentation af amplitude-modulerede toner (dvs. respons amplitude falde eksponentielt over tid)21,22. Tilpasning er blevet fortolket som en neurale mekanisme, der afspejler tabet af nyhed af en monotont gentagne sensoriske stimulus, øge følsomheden over for relevante udsving i det akustiske miljø23,24. I auditiv pathway kan tilpasning forbedre taleforståelsen i støjende omgivelser. Desuden kan denne proces være en del af de eksisterende mekanismer til at overvåge den auditive feedback af ens egen stemme til at styre tale produktionen.
Analyse af tiden udvikling af 40 Hz ASSR hos mennesker, Van Eeckhoutte et al.25 observeret en signifikant, men lille fald i respons amplitude over tid (omkring-0,0002 μV/s baseret på gruppen analyse, når de antager en lineær nedgang over tid). Disse forfattere konkluderede derfor, at 40 Hz ASSR hos mennesker ikke tilpasser sig stimuleringen. I mennesker, ikke-stationær adfærd er blevet observeret ved analyse af stabiliteten af SSVEP26. Disse forfattere bemærkede, at amplituden af den grundlæggende frekvens og den anden harmoniske af SSVEP var stationært i kun 30% og 66,7% af de, de testede, hhv. Faserne i begge SSVEP-frekvens komponenter, selv om relativt stabile over tid, udstillet små driver26.
Derfor, selv om den klassiske tid-domæne gennemsnit af efterfølgende erhvervede epoker tillader udforskning af stationære egenskaber af neurale medrivning, denne metode skal revideres, når langsigtede dynamik i indbegrebet er i fokus i den forskning, eller når gennemsnittet af kortsigtede dynamik er ødelagt af forekomsten af langsigtede dynamik. For at karakterisere ikke-stationær adfærd af Steady-State svar, bør den fremkaldte respons beregnet på et givet tidsvindue ikke kompromitteres af dem, der er beregnet i de foregående EEG segmenter. Med andre ord bør det fremkaldte potentiale ekstraheres fra baggrundsstøjen, uden at epoker er tids domæne gennemsnitligt med de foregående EEG-segmenter.
I denne undersøgelse, en metode til vurdering af dynamikken i neurale medrivning er præsenteret. Steady-State-respons registreres gentagne gange som reaktion på den samme stimulering, hvor fortløbende optagelser er Interleaved med et hvile interval på tre gang længden af den eksperimentelle løbetur. I betragtning af, at hvis tids udviklingen i den fysiologiske respons er den samme i forskellige uafhængige eksperimentelle kørsler (uafhængige optagelser), er optagelser kolonne vise gennemsnit. Med andre ord er epoker, der svarer til den samme placering i de forskellige optagelser, gennemsnitligt uden gennemsnitlige epoker i en optagelse. Som følge heraf vil respons amplitude beregnet ved ethvert stimulerings interval svare til den øjeblikkelige amplitude af det fremkaldte potentiale. De sensoriske reaktioner kan enten analyseres i tids domænet eller omdannes til frekvens domænet, afhængigt af formålet med eksperimentet. Under alle omstændigheder, amplituder kan plottet som en funktion af tid til at analysere tids udviklingen af Steady-State respons. Generering og udryddelse af Steady-State fremkaldte potentialer kan vurderes ved at begrænse analysen til den første og sidste epoker af optagelserne.
Dynamikken i neurale medrivning kan analyseres ved hjælp af andre tilgange, såsom smalbånds filtrering enkelt-Trial målinger omkring hyppigheden af interesse og beregning af konvolutten af strøm signalet ved hjælp af low-pass filtrering25 og Hilbert transformation27. Sammenlignet med disse metoder giver den kolonne vise gennemsnit af epoker mulighed for at beregne Steady-State-parametre baseret på signaler med det højere signal-støj-forhold (SNR). For nylig, Kalman filtrering er dukket op som en lovende teknik til vurdering af 40-Hz ASSR amplituder28,29,30. Gennemførelse af Kalman filtrering kan forbedre påvisning af Steady-State svar tættere på den elektrofysiologiske tærskel og reducere tidspunktet for høringen test29. Desuden er stationære reaktioner ikke nødvendige for at blive antaget, når en Kalman filtreringsmetode anvendes til at anslå ASSR amplitude30. Ikke desto mindre, kun én undersøgelse har analyseret tids udviklingen af ASSRs ved hjælp Kalman filtrering25. Konklusionen af undersøgelsen er, at 40-Hz ASSR amplitude er stabil over stimulations intervallet. Derfor skal Kalman-filtrering testes under forhold, hvorunder ASRR ikke er stationært.
Selvom det er tidskrævende, er den kolonne vise gennemsnitsmetode model-fri og behøver ikke initialiseringsværdier og/eller a priori definitioner af støj adfærden. Da det ikke indebærer konvergens tider, kan den kolonne vise gennemsnit give en mere pålidelig gengivelse af neurale indtrækninger. Derfor, de opnåede resultater med kolonne-Wise gennemsnit metode kan betragtes som jorden sandhed for at analysere dynamikken i neurale medrivning ved hjælp af Kalman filtrering.
Denne beskrivelse af protokollen er baseret på et eksempel på SSVEP. Men, det er vigtigt at bemærke, at den metode, der præsenteres her er modalitet-uafhængig, således at det også kan bruges til at analysere tids udviklingen af SSSEP og ASSR.
Protocol
Denne undersøgelse blev udført under godkendelse af forsknings-og etik komitéen under Universidad de Valparaíso, Chile (vurderings erklæring Code CEC170-18), som blev bekræftet til de nationale retningslinjer for forskning med mennesker.
1. forberedelse
- Velkommen emnet.
- Forklar undersøgelsens mål og relevans. Giv en beskrivelse af relevante tekniske detaljer. Besvar alle spørgsmål grundigt.
- Eksplicit nævner, at hun/han kan afbryde den eksperimentelle session til enhver tid, hvis det ønskes.
- Bed volontøren om at læse emnet informeret samtykke og underskrive den tilsvarende formular. Afbryde forsøgs sessionen, hvis der ikke indhentes informeret samtykke.
2. forberedelse af emnet
- Bed motivet om at sidde i en laboratorie stol i en behagelig stilling.
- Rengør hovedbunden med ethanol (en opløsning på 95%) for at fjerne laget af døde hudceller og sebum, der dækker det. Dette trin er vigtigt at reducere impedans mellem elektroderne og hovedbunden.
- Mål hovedomkredsen med et målebånd for at definere størrelsen af den elektrode hætte, der skal anvendes.
- Bed motivet om at bære elektrode hætten. Giv instruktionerne til komfortabel, men korrekt positionering af hætten.
- Mål afstanden mellem nasion (NZ, midten punkt af nasofrontal sutur, som kan identificeres ved depression mellem øjne og toppen af næsen) og fra (iz, fremtrædende af occipital knogle) ved hjælp af et målebånd.
- Mål afstanden mellem venstre og højre præ-aukulære punkter (identificeret som depression lige før øremuslingen af ørerne) ved hjælp af et målebånd.
- Korriger elektrode hætten, så skæringspunktet mellem de imaginære linjer, der er defineret i de foregående trin, svarer til toppens knudepunkt. Sørg for, at motivet er behageligt efter justeringerne.
- Anbring ledende gel i elektrode holderne i henhold til de steder, som er taget i betragtning ved forsøget.
- Brug 64 Hovedbunds placeringer efter det internationale 10-20 system31 til at bruge resultatet af protokollen til at udføre analyse af kilde lokalisering. Det højeste antal elektrode placeringer (128) på hovedbunden kan anvendes, hvis det er nødvendigt.
- Implementer kliniske eller ambulante indstillinger (med kun få elektroder), hvis der ikke er planlagt en analyse af kilde lokalisering. Brug occipital lokationer til at registrere SSVEP, temporale lokationer for at erhverve ASSR og parietal lokationer til registrering af SSSEP.
- Tryk elektroderne i elektrode holderne. Sørg for, at etiketten på elektroden matcher etiketten på placeringen i hætten.
- Ledsage volontøren til forsøgs lokalet (fortrinsvis et afskærmet, lyddæmpet kammer). Bed motivet om at sidde i en stol inde i lokalet i en behagelig position.
- Placer eksterne elektroder på næsen og ørelapper, hvis en fysisk reference (forskellig fra hovedbunden elektroder) vil blive anvendt til re-reference EEG optagelsen (i trin 3.8.1).
- Placer eksterne elektroderne i periokulære områder.
- Placer elektroderne på kinden og frontal området af hovedet, ca. 1 cm over øjenbrynene, for at optage blinkende (i trin 2.6.1).
- Placer elektroderne på den udvendige canthus af øjnene, ca. 1 cm over/under midterlinjen, for at registrere øjenbevægelser (i trin 2.6.1).
Bemærk: electrooculogram (EOG) vil blive brugt i Step 3.8.5 for at fjerne ÆGGE artefakter induceret af blinkende og øjenbevægelser.
- Drej EEG-anskaffelses systemet på, og kontrollér elektrode impedansen, hvis der anvendes et lavimpedans system til optagelse af EEG. Korriger impedansen efter behov i overensstemmelse med producentens anvisninger. Impedans skal holdes under 10 kΩ32.
- Bed motivet om at blinke og flytte øjnene i forskellige retninger for at sikre, at EOG registreres korrekt.
- Hvis du vil analysere dynamikken i SSVEP, skal du justere skærmens placering i den lodrette retning, så den passer til motivet. Dæmp lysene i rummet, indtil der opnås et komfortabelt niveau. Juster skærmens luminans niveau til den øvre grænse for deltagerens komfort niveau.
- For at analysere dynamikken i ASSR, Indsæt øretelefonerne ved hjælp af de korrekte skum skær, så øretelefonerne passer til øregangen. Kontrollér, at lydene leveres med den ønskede intensitet (f. eks. et psykofysisk behageligt niveau33).
3. EEG-erhvervelse og forbehandling
- Indstil de stimulus parametre, der er defineret i det eksperimentelle design. Se brugervejledningen fra producenten af stimulerings systemet for detaljer om softwaren.
Bemærk: for omfattende forklaringer af den stimulus, der anvendes til generering af SSVEP og ASSR, se henholdsvis Norcia et al.8 og Rance34. - Instruere emnet til at være opmærksom på stimulering, i tilfælde af, at visuelle indfald er emnet for eksperimentet.
- Præsenter en tekstet film med lyden slukket, når auditive medrivning er emnet for eksperimentet.
Bemærk: præsentation af en stumfilm giver mulighed for afbøjning af opmærksomhed fra den akustiske stimulation, samtidig med at ophidselse niveau25.
- Præsenter en tekstet film med lyden slukket, når auditive medrivning er emnet for eksperimentet.
- Nuværende stimuli længere end 90 s, som er blevet gjort for at undersøge den tid udviklingen af ssveps og ASSR i både mennesker og dyr modeller21,22,25,26.
Bemærk: nuværende stimuli kortere varighed, hvis der er udført en pilotundersøgelse. - Sæt stimuleringen på pause i 2 minutter, hvis der kun testes én eksperimentel tilstand. Interagere med emnet for at kontrollere bevidstheden.
Bemærk: varigheden af pausen afhænger af varigheden af stimuleringen. Pauser 3x længere end stimulation intervaller vil sikre, at et svar fremkaldt af en stimulus ikke påvirkes af den tidligere stimulation. Længere pauser er tilladt, hvis motivet anmoder herom.- Sæt stimuleringen på pause i mindst 10 s, når forskellige eksperimentelle betingelser er afprøvet, da vekslende stimulation med pauser på 10 s er blevet foreslået for at mindske ekstra tilpasnings effekter og reducere længden af eksperimentet25.
- Gentag præsentations trinnene (trin 3.3-3.4) mindst 30x for at sikre det høje SNR af målingerne efter gennemsnittet af epoker (trin 4,4).
- Optag EEG ved hjælp af standardprocedurer35. Opret en separat EEG-fil for hver eksperimentel kørsel.
Bemærk: Se brugermanualen til anskaffelses systemet for detaljer om softwaren. - Overvåg EEG-optagelsen for at registrere søvn perioder baseret på niveauet af alfaaktivitet og den hyppighed, hvormed blinkende artefakter vises. Pause eksperimentet, når forhøjede alfa niveauer ledsaget af nedsat blinkende frekvenser opdages, hvilket indikerer søvnighed. Afvis forsøgs kørslen fra yderligere analyse, når der registreres søvn perioder.
- Beregn amplitude af Steady-State respons ved afslutningen af hver eksperimenterende løb, efter instruktion i brugermanualen for den erhvervelse software, der anvendes i forsøget.
- Overvåg den opmærksomme niveau af emnet ved at sammenligne amplituden af Steady-State svar opnået ved afslutningen af hver eksperimenterende løb. Indstil Steady-State amplitude opnået i den første eksperimentelle kørsler som en reference amplitude.
- Indstil en afvisnings tærskel (et fald i respons amplitude på 5% vedrørende reference amplituden). Afviser de eksperimentelle kørsler, hvor amplituden af Steady-State respons opfylder afvisnings kriterierne.
- Afslut den eksperimentelle session efter erhvervelse af antallet af kørsler, der er defineret i det eksperimentelle design.
- Forbehandl EEG-data offline ved hjælp af standard EEG-procedurer35 beskrevet i de næste trin pr. producentens anvisninger.
- Genreference optagelsen ved hjælp af en gennemsnitlig reference (gennemsnit af alle optagelses elektroder) eller gennemsnittet af en delmængde af elektroder. Alternativt kan du bruge en fysisk reference (f. eks. eksterne elektroder placeret på næsen og ørelapper beskrevet i trin 2,4).
- Konverter elektrode koordinaterne til det internationale 10/20-system, hvis det radiale koordinatsystem blev anvendt i forbindelse med EEG-anskaffelserne. Se producentens manual for detaljer om konvertering.
- Band-pass Filtrer EEG-signalet mellem 0,5-300 Hz. Indstil et hak-filter (centreret ved 50 Hz eller 60 Hz), hvis det er nødvendigt.
- Ned-Sample EEG-signalet for at reducere udførelsestiden for den algoritme, som er valgt til fjernelse af okulære artefakter (trin 3.8.5).
Bemærk: en prøvetagningsfrekvens på 512 Hz er tilstrækkelig til at analysere hyppigheden af hjerne svingningerne under 40 Hz35. - Fjern de okulære artefakter.
Bemærk: til dette formål kan forskellige teknikker anvendes (Se Urigüen og Garcia-Zapirain35 for en omfattende gennemgang af artefakt fjernelse algoritmer). Blandt dem, uafhængig komponent analyse er en af de mest udvidede metoder og er implementeret i både kommerciel og fri analyse software37,38,39. - Segmentér EEG-dataene i epoker time-Locked til stimulation. Vælg epoker længde i henhold til formålet med eksperimentet.
Bemærk: Epochs bør være tilstrækkelig lang til, at der kan foretages en analyse af Steady-State-reaktionen i frekvensområdet med en passende spektral opløsning. - Kør ikke algoritmer til afvisning af artefakt på dette stadie for at finde og fjerne epoker, der indeholder artefakter.
Bemærk: Hvis du fjerner epoke på dette stadie, vil det medføre fejl, når datasættet er organiseret til at køre den kolonne vise gennemsnit af epoker (trin 4,2 og 4,4). Afvisnings algoritmer implementeres på et senere behandlingstrin (trin 4.1.4). - Kør funktionen DC-detrend for at beregne DC-trends i individuelle EEG-epoker og korrigere dem.
- Kør den oprindelige korrektionsfunktion for at korrigere grundlinjen for optagelsen. Vælg pre-stimulus tidsintervaller længere end 200 MS.
Bemærk: oprindelig korrektion består af gennemsnit af dataene i det valgte tidsinterval. Gennemsnittet beregnes for hver kanal og trækkes fra hvert datapunkt i hver epoke.
4. beregning af respons amplituder
- Angiv de parametre, der er nødvendige for beregningen af Steady-State-responserne (figur 1a).
Bemærk: den in-House kode, der bruges til behandling af data, er frit tilgængelig på < https://state_visually_evoked_potentials_SSVEP_elicited_in_humans_by_continuos_light_modulated_in_amplitude_at_10Hz/62573 >. Se hjælp-teksten i koden for at få yderligere oplysninger. Tilsvarende er en delmængde af de data, der anvendes i dette studie, tilgængelig.- Angiv antallet af optagelser (eksperimentelle kørsler) af eksperimentet.
- Indtast epoker længde for at segmentere de enkelte optagelser.
- Angiv prøveudtagningsfrekvensen for eksperimentet.
- Vælg algoritmer til afvisning af artefakt for at registrere og fjerne epoker, der indeholder artefakter. De tilgængelige udvælgelseskriterier er 1) gradient (absolut forskel mellem to på hinanden følgende prøver), 2) max-min (forskellen mellem den maksimale og mindste amplitude i epoken), og 3) amplitude (absolutte maksimum og minimum amplituder).
- Kør behandlings koden.
Bemærk: trin 4.2-4.7 udføres automatisk, når denne indstilling er valgt. Kør trinnene manuelt, hvis det er relevant.
- Arranger epoker igen i en data matrix med n rækker og m kolonner, hvor n repræsenterer antallet af optagelser (eksperimentelle kørsler) og m antallet af epoker (figur 1b).
- Vægt epoken til at dæmpe effekten af motion og muskuløse artefakter.
Bemærk: vægtede EEG-epoker opnås ved at dividere hver spændings prøve med amplitude variansen af den epoke, de tilhører, således at variansen anvendes som et mål for amplitude variabilitet og vægtningsfaktor40. - Kolonne vise gennemsnit af datasættet. Til dette formål gennemsnit tids domæne epoker svarer til samme tidsvindue i de forskellige optagelser.
Bemærk: dette trin gør det muligt at beregne Steady State-amplituden i optagelser med et bemærkelsesværdigt højt signal-støj-forhold (SNR). - Eksporter tidsserierne som følge af gennemsnittet for yderligere analyse af tids udviklingen for indstigning i ekstern software.
- Beregn amplitude af Steady-State respons i hver epoke som følge af kolonne-Wise gennemsnit, ved hjælp af fast Fourier transformation (FFT).
Bemærk: FFT-længden skal svare til længden af en epoke. Implementeringen af en vindues teknik er ikke obligatorisk. Amplituden af Steady-State respons er defineret som den spektral amplitude opnået ved hyppigheden af amplitude modulation af de sensoriske stimuli.
- Beregn amplitude af Steady-State respons i hver epoke som følge af kolonne-Wise gennemsnit, ved hjælp af fast Fourier transformation (FFT).
- Vektor gennemsnitligt amplitude af et ad-hoc antal FFT siloer på hver side af hyppigheden af respons til at beregne rest støjniveau (RNL). Antallet af FFT-siloer skal svare til et frekvensbånd på ca. 3 Hz på hver side af reaktions frekvensen.
Bemærk: den høje frekvens-specificitet af Steady-State respons gør respons amplitude uafhængig af disse baggrunds svingninger med tilsvarende frekvenser, som igen fordeler ensartet i et relativt snævert frekvensbånd41 , 42 , 43. - Plot amplituden af Steady-State respons og RNL som en funktion af kolonneindeks (dvs. antallet af den erhvervede epoke) for at udforske udviklingen af Steady-State respons under stimulerings intervallet.
Representative Results
SSVEP blev fremkaldt af kontinuerlige visuelle stimuli af 40 s i længden, hvor lysintensiteten blev moduleret af en sinusformet bølge på 10 Hz (modulations dybde på 90%). Stimuli blev leveret af fire lysemitterende dioder (lysdioder) beliggende i midten af en 50 cm x 50 cm sort skærm, som toppunkter af en 5 cm x 5 cm firkant. Når deltageren sad 70 cm fra skærmen, arealet af kvadratet af lysdioder subtendens en visuel vinkel på omkring 4 °. LED-skærmen blev designet ved hjælp af et USB-baseret microcontroller udviklingssystem og fire super lyse hvide lysdioder med en diameter på 10 mm. Den Pulse width modulation (PWM) teknik blev brugt til at styre den strøm, der leveres til lysdioderne. Denne teknik kontrollerede lysdioderne intensiteter ved en given frekvens og generere den endelige sinusformet konvolut. En PWM-frekvens på 40 kHz blev brugt til at undgå en opfatte flimren effekt.
Tredive optagelser blev opnået, som blev segmenteret i epoker af 4 s. Derfor blev der opnået et datasæt bestående af 10 kolonner (antal EEG-epoker i optagelser) og 30 rækker (antal optagelser, antal eksperimentelle kørsler).
Den neurale svingnings tid-låst til stimulation blev tydelig som den kolonne-Wise gennemsnit blev udført (figur 2). Signifikant, kan det interval, hvor SSVEP genereres, observeres i Traces, der svarer til kolonne 1. I denne kolonne er 0,2 s af præ-stimulus baseline afbildet ud over de første 0,8 s af neurale entrainment. Derfor er den procedure, der er beskrevet her, tillader karakterisering af 1) dynamikken i den oscillatoriske respons, når neurale medrivning er allerede etableret, og 2) inddragelse af neurale svingninger. En eller flere epoker registreret efter afslutningen af stimulation kan også inkluderes i data matrixen for at studere udslettelse af Steady-State respons efter stimulus offset.
Under den kolonne vise gennemsnit af epoker, faldt middelamplitude af ssvep (spektral amplitude ved 10 Hz, beregnet ved anvendelse af FFT) under gennemsnittet af de første epoker af kolonnerne og tendens til at stabilisere bagefter (figur 3a). Dette resultat er enigt i tidligere undersøgelser, som analyserede udviklingen af ASSR under gennemsnittet af sekventielt erhvervede epoker21,22,40,43,44. Opførsel af respons amplitude under gennemsnittet er normalt forklares ved den relativt høje bidrag af ikke gennemsnitlige støj til respons amplitude beregnet i den første epochs, som er svækket som gennemsnit udføres13, 44 , 45 , 46 , 47. bemærkelsesværdigt, den SSVEP amplitude variabilitet signifikant faldt som gennemsnit skred frem.
Vi analyserede også rnl af målingerne under den kolonne vise gennemsnit af epoker (figur 3b). RNL blev beregnet i et snævert frekvensbånd (3 Hz) på begge sider af frekvensen af SSVEP. Selv om denne procedure ikke er almindelig, når ssvep analyseres, er vektor-gennemsnit af et givet antal frekvens beholdere omkring den neurale medrivning standarden for estimering af rnl i ASSR-målinger41,42, 43. som forventet faldt rnl gradvist, da antallet af gennemsnitlige epoker steg og nåede det asymptotiske niveau efter ca. 20 epoker blev behandlet. I modsætning til det, der blev observeret, da SSVEP-amplituden blev analyseret, forblev standardafvigelsen i RNL forholdsvis konstant, da antallet af gennemsnitlige epoker steg, hvilket tyder på, at optagelsesforholdene var stabile langs forsøgs sessionen.
De resultater, der blev præsenteret ovenfor, fastsatte ændringerne i det maksimale signal-støj-forhold (pSNR) for målingerne under den spalte vise gennemsnit af Epoch (figur 3C). Dette udtryk defineres her som forholdet (i dB) mellem den firkantede amplitude af responset (SSVEP) og kvadrat amplitude af RNL. Som et gennemsnit steg psnr, da antallet af gennemsnit for epoker steg til 18, ca. Yderligere stigninger i antallet af gennemsnit for epoker påvirkede ikke kvaliteten af signalet væsentligt. Variabiliteten af pSNR faldt, da flere epoker blev gennemsnitligt.
Endelig er dynamikken i SSVEP amplitude og RNL repræsenteret i figur 4. Disse tids udviklinger blev opnået ved at afbilde respons parametrene beregnet i slutningen af kolonne-Wise gennemsnit af epoker som en funktion af antallet af kolonner (som en funktion af tid). Som påvist af Labecki et al.26kan dynamikken i SSVEP variere betydeligt mellem. Da de resultater, der præsenteres i figur 4 , svarer til en enkelt person, kan der ikke foretages generaliseringer. I dette emne, amplituden af SSVEP viste en relativt kompleks adfærd (figur 4a). Respons amplitude steg gradvist i løbet af de første 12 sekunder efter stimulus debut (tid, der svarer til længden af 3 epochs). Da stimuleringen varede, faldt SSVEP konsekvent i løbet af de følgende 12 sekunder, og forblev relativt konstant bagefter. Disse resultater kan ikke forklares ved RNL-funktionsmåden, da denne parameter var relativt konstant under stimulerings intervallet (figur 4b). Stigningen i SSVEP amplitude efter stimulus debut er tydelig i de spor, der præsenteres i figur 2 og kan forklares ved integrations processer, som resulterer i stabilisering af neurale indtrækninger. Den efterfølgende nedgang i amplitude tyder på tilpasningen af SSVEP til den vedvarende stimulation. Ikke desto mindre skal disse hypoteser testes i kontrollerede eksperimenter med tilegnede stikprøvestørrelser.
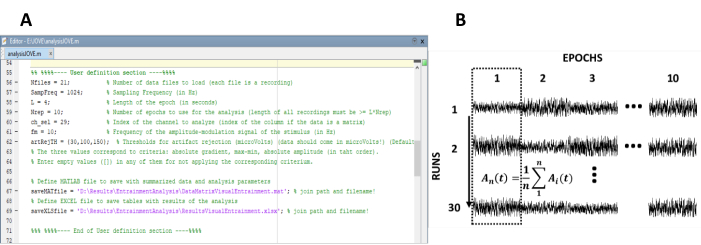
Figur 1 : Kritiske trin til at udtrække den tids udvikling af amplituden af Steady-State svar. (A) skærmbillede af behandlings koden, hvor der er defineret analyseparametre. B) repræsentativt diagram, som illustrerer organiseringen af datasættet. En data matrix bestående af 30 optagelser af 10 epoker er repræsenteret. Den kolonne vise gennemsnit af epoker er fremhævet i den første kolonne. Den lodrette linje repræsenterer retningen af gennemsnits snittet. Venligst klik her for at se en større version af dette tal.

Figur 2 : Ændringer i bølgeform af Steady-State visuelt fremkaldt potentialer (SSVEP) under kolonne-Wise gennemsnit af epochs. Svarene blev fremkaldt af den kontinuerlige præsentation af lys moduleret i amplitude ved 10 Hz. Rækkerne viser bølge formerne opnået efter gennemsnittet af alle tidligere optagelser (dvs. række 1 er den første optagelse, række 5 er den bølgeform, der opnås efter gennemsnittet af de første fem optagelser, og den sidste række er gennemsnitsværdien af alle optagelser). Mere pålidelige bølgeformer af SSVEP blev observeret i hver kolonne, da antallet af gennemsnitlige kørsler steg. For at skabe klarhed (for at gøre svingningerne i SSVEP synlige), er kun det første sekund af epoker repræsenteret. Undtagelserne er spor i den første kolonne i datasættet, for hvilke der vises 0,2 sekunders præ-stimulus-baseline. Venligst klik her for at se en større version af dette tal.

Figur 3 : Ændringer i respons-og optagelses parametrene under den kolonne vise gennemsnit af epochs. A) udviklingen i SSVEP-amplituden. (B) opførsel af rnl. C) ændringer i psnr. Sorte streger repræsenterer middelværdierne for hver kolonne (n = 10), og den grå skygge repræsenterer det område, som er omfattet af ± en standardafvigelse. Venligst klik her for at se en større version af dette tal.

Figur 4 : Tid udvikling af SSVEP fremkaldt af præsentationen af kontinuerlig visuel stimulation, moduleret i amplitude ved 10 Hz. A) tidsforløbet for SSVEP-amplituden. B) tidsforløbet for rnl. Venligst klik her for at se en større version af dette tal.
Discussion
Dette arbejde beskriver en eksperimentel procedure for at analysere dynamikken i oscillatoriske hjernen svar. En sådan metodologi består i at erhverve et tilstrækkeligt antal uafhængige eksperimentelle kørsler af samme eksperimentelle tilstand og tids domæne gennemsnitlige epoker svarende til samme tidsvindue i de forskellige optagelser (kolonner-Wise gennemsnit i Figur 1B). Den amplitude, som beregnes i de gennemsnit data, repræsenterer den øjeblikkelige amplitude af det oscillatoriske respons. Plotning disse amplituder som en funktion af tid (eller antallet af kolonner i datasættet) gør det muligt at analysere tids udviklingen af den oscillatoriske responstid-låst til stimulation. Denne metode er en ændring af den, der foreslås af Ritter et al.23 til analyse af tilpasningen af forbigående kortikale fremkaldte potentialer. Metoden er blevet brugt til at analysere dynamikken i auditive fremkaldte potentialer i både mennesker24 og dyremodeller20,21.
Fra et metodologisk synspunkt, kombinationen af parametre, der anvendes til at fremkalde Steady-State respons og dem, der gennemføres for at udtrække den neurale respons fra baggrundsstøjen er afgørende for at analysere tids udviklingen af Steady-State fremkaldte potentialer 22. den stimulus længde, der blev anvendt i forsøget præsenteret her (40 s), blev udvalgt på grundlag af resultater opnået i en pilotundersøgelse. Denne stimulus længde var tilstrækkelig til at analysere tilpasningen af ASSR genereret i rotte hjernestammen21,22. Desuden bør stimulus længde overskride det tidspunkt, hvor den asymptotiske øjeblikkelige bånd effekt af SSVEPs er nået (figur 1 i labecki et al.26). Ikke desto mindre kan den asymptotiske øjeblikkelige bånd effekt af SSVEPs nås ud over 60 ' erne i nogle tilfælde (figur 2 i labecki et al.26). Derfor, at køre en lille stikprøve pilotundersøgelse anbefales at definere stimulus længde af stimulation. Ellers anbefales en stimulus længde længere end 90 s for at opnå fuldstændig repræsentation af tids udviklingen af responset. Brug af tilstrækkeligt lange pauser mellem på hinanden følgende optagelser indebærer, at man overvejer fortløbende eksperimentelle kørsler som statistisk uafhængige (dvs. forskellige uafhængige målinger af samme variabel). Efter vores bedste overbevisning er der ikke udført eksperimenter for at analysere den optimale pause mellem kørsler (minimum pause kræves for at gøre kørsler uafhængige af hinanden). Kriterier for at bruge pauser mindst 3x længere end stimulus længde er konservativ nok til at sikre, at Steady-State respons registreret i en given kørsel ikke påvirkes af den tidligere stimulation.
For nylig er vekslende stimuli (eksperimentelle betingelser) blevet foreslået som et valg til at reducere pause mellem eksperimentelle kørsler, undgå ekstra tilpasnings effekt25. Ligeledes er antallet af eksperimentelle kørsler (30) implementeret i denne forsøgsprotokol konservativ, da det asymptotiske RNL og pSNR typisk nås efter gennemsnitligt 20 eksperimentelle kørsler, omtrent. Når stimuli falder inden for den midterste øvre region af det dynamiske område af reaktionen (høj fornemmelse niveauer), lavere antal kørsler er sandsynligvis nødvendige for at analysere dynamikken i den fremkaldte respons. I tilfælde, hvor der testes forskellige forsøgsbetingelser, er det imidlertid afgørende at have det samme antal forsøgsserier for at foretage sammenligninger mellem tilstande (dvs. forskellige fornemmelse).
Ud over den kolonne-kloge gennemsnit af epochs, dynamikken i oscillerende fremkaldt potentialer er blevet analyseret ved at filtrere enkelt-Trial målinger i en smal frekvensbånd omkring hyppigheden af interesse og computing konvolutten af magten signal ved hjælp af lavpasfiltrering26. Ligeledes er der gennemført en enkelt analyse undersøgelse for at karakterisere den overgangsperiode, der går forud for den stabile region SSVEP48, og ændringerne i amplitude og fase af SSVEP i det stabile område af svaret49. Mens enkelt forsøg analyser tillader diskrimination af relativt hurtige udsving i respons amplitude, eksperimentelt design til at analysere den gennemsnitlige respons i blokke adskilt med en given Inter-Block interval kun tegner sig for langsigtede variationer i amplitude af det fremkaldte potentiale50,51. Den kolonne-kloge gennemsnit af epoker står mellem disse to muligheder. Konvertering af det gennemsnitlige signal til frekvens domænet ved hjælp af FFT indebærer at analysere dynamikken i reaktionen med en opløsning svarende til længden af epoken. I eksemplet præsenteret her, blev SSVEP rapporteret hver 4 s. Selv om 4 s opløsning er tilstrækkelig til at beskrive dynamik, der opstår med intervaller af tid overgår snesevis af sekunder, såsom den af ssvep26, delvist overlappende epoker i de oprindelige optagelser gør det muligt at beskrive den tid, udvikling af Steady-State respons på en mere raffineret måde25.
Dynamikken i Steady-State respons opnået efter kolonne-Wise gennemsnit af epoker primært repræsenterer udviklingen af den oscillatoriske aktivitet, der er synkroniseret blandt de gennemsnitligt EEG segmenter (dem, der overlever gennemsnittet). Derfor er et vigtigt spørgsmål vedrørende gennemførligheden af metodologien den mulige dæmpning af respons amplituder som følge af variationer i fasen af neurale svingninger fra en uafhængig eksperimentel kørsel til en anden (dvs. blandt optagelser). Dette emne skal behandles eksperimentelt. Men, tyder på, at fasen af hjernens oscillatoriske respons er mindre variabel end forventet. Faktisk, flere undersøgelser har rapporteret en regelmæssighed i den forventede fase af human 80 Hz ASSR47,48,49. Når ventetider anslås på grundlag af den oscillatoriske aktivitetsfase, er den forudsigelige effekt af intensiteten og bære frekvensen for de akustiske stimuli på latenstid af de auditive responser blevet observeret (dvs. forsinkelsen falder, da intensitet og bærefrekvens stigning)52,53,54. Desuden er der også observeret typiske modale ændringer i amplitude og venstre mod højre-asymmetri i høre niveauerne, når latorer estimeres fra fase af ASSR47,55,56 , 57 , 58. ved beskrivelsen af tids udviklingen for SSVEP ved hjælp af analyse af enkelt forsøg bemærkede labecki et al.26 , at selv om variabiliteten af respons amplituder inden for samme forsøgsperiode var meget høj, var variabiliteten af fase var betydeligt mindre udtalt.
På grundlag af deres observationer foreslog Labecki et al.26 , at der skulle gennemsnitligt være et minimum af 50 forsøg for at opnå et pålideligt skøn over den gennemsnitlige effekt konvolut for responset. Disse resultater indikerer, at selv når amplituden af responset er beregnet i enkelt forsøg, gennemsnit (af konvolutter i dette tilfælde) er nødvendig for at rapportere troværdige resultater. Desuden tyder den inter-forsøg variabilitet i amplituden af SSVEP rapporteret af Labecki et al.26 på, at beregningen af denne parameter i enkelt forsøg kan være stærkt påvirket af baggrundsstøj. I betragtning af udviklingen i signal-støj-forholdet i figur 2reducerer beregningen af responset i det gennemsnitligt signal i stedet for enkelte forsøg i væsentlig grad antallet af EEG-segmenter, der skal forarbejdes for at opnå pålidelige Målinger. Desuden understøtter den lave variabilitet i fase opnået af labecki et al.26 idéen om, at den kolonne vise gennemsnit af epoker præsenteret her er en gyldig procedure til beregning af dynamikken i oscillatoriske fremkaldte potentialer.
Gennemsnit af data på forskellige niveauer fører til en anden fortolkning af resultaterne. Med hensyn til oscillerende fremkaldt potentialer, computing respons amplitude efter den tid-domæne gennemsnit af uafhængige kørsler indebærer at analysere kun tid-låste svingninger (dvs. dem, der overlever gennemsnittet). Denne procedure kan filtrere relevante oplysninger om dynamikken i reaktionen i individuelle forsøg. Det garanterer imidlertid et tilstrækkeligt højt signal-støj-forhold mellem målingerne. Dette aspekt kan være af betydning, når svarene er tæt på den elektrofysiologiske tærskel, en tilstand, hvor påvisning af inddæmningen kan blive kompromitteret på grund af lav signal-til-støj ratio af målingen.
Disclosures
Forfatterne har intet at afsløre.
Acknowledgments
Forfatterne anerkender taknemmeligt Lucía Zepeda, Grace A. Whitaker og Nicolas Nieto for deres bidrag til video produktion. Dette arbejde blev delvist støttet af CONICYT programmer BASAL FB0008, MEC 80170124 og PhD Scholarship 21171741, samt National Institute on døvhed og andre kommunikationsforstyrrelser af National Institutes of Health under tildeling nummer P50DC015446. Indholdet er udelukkende ansvaret for forfatterne og ikke nødvendigvis repræsenterer de officielle synspunkter af de nationale institutter for sundhed.
Materials
| Name | Company | Catalog Number | Comments |
| Active electrodes | Biosemi | P32-1020-32ACMS (ABC) | for channels 1-32 |
| Active electrodes | Biosemi | P32-1020-32A (ABC) | for channels 33-64 |
| Active electrodes | Biosemi | 8 x TP FLAT | external electrodes |
| Active-Two adquisition system | Biosemi | version 7.0 | EEG adquisition system |
| alcohol | Salcobrand | Code: 3309011 | for cleaning the scalp |
| Electrode cap 64 channels | Biosemi | CAP MS xx yy | cap |
| Electrode cap 64 channels | Biosemi | CAP ML xx yy | cap |
| gel | Biosemi | SIGNA BOX12 | conductive gel |
| Laptop | Asus | Core i7 1TB DD + 128GB SSD 8GB RAM | computer for stimulation |
| Laptop | Asus | Core i7 1TB DD + 128GB SSD 8GB RAM | computer for recording |
| LED screen | in-house production | - | The screen consists of four light-emitting diodes (LEDs) situated on the center of a 50x50 cm black screen, as vertexes of a square of 5x5 cm |
| sterile gauze | Salcobrand | Code: 8730277 | for cleaning the scalp |
References
- Berger, H. Über das elektrenkephalogramm des menschen. European Archives of Psychiatry and Clinical Neuroscience. 87 (1), 527-570 (1929).
- Berger, H. Electroencephalogram of humans. Journal fur Psychologie und Neurologie. 40, 160-179 (1930).
- Walter, W. G. The location of cerebral tumours by electro-encephalography. The Lancet. 228 (5893), 305-308 (1936).
- Jasper, H. H., Andrews, H. L. Electro-encephalography: III. Normal differentiation of occipital and precentral regions in man. Archives of Neurology & Psychiatry. 39 (1), 96-115 (1938).
- Snyder, A. Z. Steady-state vibration evoked potentials: descriptions of technique and characterization of responses. Electroencephalography and Clinical Neurophysiology. 84 (3), 257-268 (1992).
- Giabbiconi, C. M., Trujillo-Barreto, N. J., Gruber, T., Muller, M. M. Sustained spatial attention to vibration is mediated in primary somatosensory cortex. Neuroimage. 35, 255-262 (2007).
- Rossion, B., Boremanse, A. Robust sensitivity to facial identity in the right human occipito-temporal cortex as revealed by steady-state visual-evoked potentials. Journal of Vision. 11 (2), (2011).
- Norcia, A. M., Appelbaum, L. G., Ales, J. M., Cottereau, B. R., Rossion, B. The steady-state visual evoked potential in vision research: a review. Journal of Vision. 15 (6), 4 (2015).
- Galambos, R., Makeig, S., Talmachoff, P. J. A 40-Hz auditory potential recorded from the human scalp. Proceedings of the National Academy of Sciences of the United States of America. 78 (4), 2643-2647 (1981).
- Picton, T. W., Skinner, C. R., Champagne, S. C., Kellett, A. J., Maiste, A. C. Potentials evoked by the sinusoidal modulation of the amplitude or frequency of a tone. The Journal of the Acoustical Society of America. 82 (1), 165-178 (1987).
- Dawson, G. D. A summation technique for the detection of small evoked potentials. Electroencephalography and Clinical Neurophysiology. 6 (1), 65-84 (1954).
- Savio, G., Cardenas, J., Abalo, M. P., Gonzalez, A., Valdes, J. The low and high frequency auditory steady state responses mature at different rates. Audiology and Neurotology. 6 (5), 279-287 (2001).
- Luts, H., Desloovere, C., Kumar, A., Vandermeersch, E., Wouters, J. Objective assessment of frequencyspecific hearing thresholds in babies. International Journal of Pediatric Otorhinolaryngology. 68, 915-926 (2004).
- Valdes, J. L., et al. Comparison of statistical indicators for the automatic detection of 80 Hz auditory steady state responses. Ear and Hearing. 18 (5), 420-429 (1997).
- Wilding, T., McKay, C., Baker, R., Kluk, K. Auditory steady state responses in normal-hearing and hearing-impaired adults: an analysis of between-session amplitude and latency repeatability, test time, and F ratio detection paradigms. Ear and Hearing. 33 (2), 267 (2012).
- de Resende, L. M., et al. Auditory steady-state responses in school-aged children: a pilot study. Journal of NeuroEngineering and Rehabilitation. 12 (1), 13 (2015).
- Halbleib, A., et al. Topographic analysis of engagement and disengagement of neural oscillators in photic driving: a combined electroencephalogram/magnetoencephalogram study. Journal of Clinical Neurophysiology. 29 (1), 33-41 (2012).
- Salchow, C., et al. Rod Driven Frequency Entrainment and Resonance Phenomena. Frontiers in Human Neuroscience. 10, 413 (2016).
- Wacker, M., et al. A time-variant processing approach for the analysis of alpha and gamma MEG oscillations during flicker stimulus generated entrainment. IEEE Transactions on Biomedical Engineering. 58 (11), 3069-3077 (2011).
- Glaser, E. M., Ruchkin, D. S. Principles of Neurobiological Signal Analysis. Journal of Clinical Engineering. 2 (4), 382-383 (1977).
- Prado-Gutierrez, P., et al. Habituation of auditory steady state responses evoked by amplitude-modulated acoustic signals in rats. Audiology Research. 5 (1), (2015).
- Prado-Gutierrez, P., Martínez-Montes, E., Weinstein, A., Zañartu, M. Estimation of auditory steady-state responses based on the averaging of independent EEG epochs. PLoS ONE. 14 (1), (2019).
- Ritter, W., Vaughan, H. G. Jr, Costa, L. D. Orienting and habituation to auditory stimuli: a study of short terms changes in average evoked responses. Electroencephalography and Clinical Neurophysiology. 25 (6), 550-556 (1968).
- Malmierca, M. S., Cristaudo, S., Pérez-González, D., Covey, E. Stimulus-specific adaptation in the inferior colliculus of the anesthetized rat. Journal of Neuroscience. 29 (17), 5483-5493 (2009).
- Van Eeckhoutte, M., Luke, R., Wouters, J., Francart, T. Stability of auditory steady state responses over time. Ear and Hearing. 39 (2), 260-268 (2018).
- Labecki, M., Nowicka, M. M., Suffczynski, P. Temporal Modulation of Steady-State Visual Evoked Potentials. International Journal of Neural Systems. 29 (3), 1850050 (2019).
- Zhang, S., et al. A study on dynamic model of steady-state visual evoked potentials. Journal of Neural Engineering. 15 (4), 046010 (2018).
- Wilson, U. S., Kaf, W. A., Danesh, A. A., Lichtenhan, J. T. Assessment of low-frequency hearing with narrowband chirp evoked 40-Hz sinusoidal auditory steady state response. International Journal of Audiology. 55 (4), 239-247 (2016).
- Kaf, W. A., Mohamed, E. S., Elshafiey, H. 40-Hz Sinusoidal Auditory Steady-State Response and Tone Burst Auditory Brainstem Response Using a Kalman Filter to Determine Thresholds Pre- and Post-Myringotomy with Grommet Tube in Children With Mild, Low-Frequency Conductive Hearing Loss. American Journal of Audiology. 25 (1), 41-53 (2016).
- Luke, R., Wouters, J. Kalman filter based estimation of auditory steady state response parameters. IEEE Transactions on Neural Systems and Rehabilitation Engineering. 25, 196-204 (2017).
- Homan, R. W. The 10-20 Electrode System and Cerebral Location. American Journal of EEG Technology. 28 (4), 269-279 (1988).
- Kappenman, E. S., Steven, J. L. The Effects of Electrode Impedance on Data Quality and Statistical Significance in ERP Recordings. Psychophysiology. 47 (5), 888-904 (2010).
- Moore, R., Gordon-Hickey, S., Jones, A. Most comfortable listening levels, background noise levels, and acceptable noise levels for children and adults with normal hearing. Journal of the American Academy of Audiology. 22 (5), 286-293 (2011).
- Rance, G. The Auditory Steady-state Response: Generation, Recording, and Clinical Application. Plural Publishing. 335, (2008).
- Luck, S. J. An introduction to the event-related potential technique. Second Edition. , MIT Press. 406 (2014).
- Urigüen, J. A., Garcia-Zapirain, B. EEG artifact removal-state-of-the-art and guidelines. Journal of Neural Engineering. 12 (3), (2015).
- Jung, T. P., et al. Removing electroencephalographic artifacts by blind source separation. Psychophysiology. 37 (2), 163-178 (2000).
- Li, Y., Ma, Z., Lu, W., Li, Y. Automatic removal of the eye blink artifact from EEG using an ICA-based template matching approach. Physiological Measurement. 27 (4), 425 (2006).
- Mannan, M. M. N., Jeong, M. Y., Kamran, M. A. Hybrid ICA-Regression: automatic identification and removal of ocular artifacts from electroencephalographic signals. Frontiers in Human Neuroscience. 10, 193 (2016).
- John, M. S., Dimitrijevic, A., Picton, T. W. Weighted averaging of steady-state responses. Clinical Neurophysiology. 112 (3), 555-562 (2001).
- Zurek, P. M. Detectability of transient and sinusoidal otoacoustic emissions. Ear and Hearing. 13 (5), 307-310 (1992).
- Lins, O. G., Picton, P. E., Picton, T. W., Champagne, S. C., Durieux-Smith, A. Auditory steady-state responses to tones amplitude-modulated at 80-110 Hz. Journal of the Acoustical Society of America. 97 (5), 3051-3063 (1995).
- John, M. S., Picton, T. W. Human auditory steady-state responses to amplitude-modulated tones: phase and latency measurements. Hearing Research. 141 (1-2), 57-79 (2000).
- Fortuny, A. T., et al. Criterio de parada de la promediación de la respuesta auditiva de estado estable. Acta Otorrinolaringológica Española. 62 (3), 173-254 (2011).
- John, M. S., Purcell, D. W., Dimitrijevic, A., Picton, T. W. Advantages and caveats when recording steady-state responses to multiple simultaneous stimuli. Journal of the American Academy of Audiology. 13 (5), 246-259 (2002).
- Luts, H., Van Dun, B., Alaerts, J., Wouters, J. The influence of the detection paradigm in recording auditory steady-state responses. Ear and Hearing. 29 (4), 638-650 (2008).
- Choi, J. M., Purcell, D. W., John, M. S. Phase stability of auditory steady state responses in newborn infants. Ear and Hearing. 32 (5), 593-604 (2011).
- Xu, M., et al. Use of a steady-state baseline to address evoked vs. oscillation models of visual evoked potential origin. Neuroimage. 134, 204-212 (2016).
- Peachey, N. S., Demarco, P. J. Jr, Ubilluz, R., Yee, W. Short-term changes in the response characteristics of the human visual evoked potential. Vision Research. 34 (21), 2823-2831 (1994).
- Woods, D. L., Elmasian, R. The habituation of event-related potentials to speech soundsand tones. Electroencephalography and Clinical Neurophysiology. 65, 447-459 (1986).
- Ravden, D., Polich, J. Habituation of P300 from visual stimuli. International Journal of Psychophysiology. 30 (3), 359-365 (1998).
- Picton, T. W., Dimitrijevic, A., John, M. S., Van Roon, P. The use of phase in the detection of auditory steady-state responses. Clinical Neurophysiology. 112 (9), 1698-1711 (2001).
- Alaerts, J., Luts, H., Van Dun, B., Desloovere, C., Wouters, J. Latencies of auditory steady-state responses recorded in early infancy. Audiology and Neurotology. 15 (2), 116-127 (2010).
- John, M. S., Brown, D. K., Muir, P. J., Picton, T. W. Recording auditory steady-state responses in young infants. Ear and Hearing. 25 (6), 539-553 (2004).
- Purcell, D. W., John, S. M., Schneider, B. A., Picton, T. W. Human temporal auditory acuity as assessed by envelope following responses. Journal of the Acoustical Society of America. 116 (6), 3581-3593 (2004).
- Kuwada, S., et al. Sources of the scalp-recorded amplitude-modulation following response. Journal of the American Academy of Audiology. 13 (4), 188-204 (2002).
- Pauli-Magnus, D., et al. Detection and differentiation of sensorineural hearing loss in mice using auditory steady-state responses and transient auditory brainstem responses. Neuroscience. 149 (3), 673-684 (2007).
- Prado-Gutierrez, P., et al. Maturational time course of the Envelope Following Response to amplitude-modulated acoustic signals in rats. International Journal of Audiology. 51 (4), 309-316 (2012).
