

Summary
本文旨在提供大鼠胚胎中扁病毒转生的方法,使用多次注射病毒悬浮液进入酶体肠。雌性大鼠与肥沃的雄性菌株交配,具有不同的显性毛皮颜色,用于产生伪怀孕的养母。
Abstract
转基因动物模型在现代生物医学研究中至关重要。将外来基因植入早期小鼠或大鼠胚胎是活生物体基因功能分析的宝贵工具。标准转基因方法基于将外来DNA片段微注入受精卵母细胞的异核。这项技术在小鼠中被广泛应用,但在其他动物物种中仍然相对低效和技术要求高。转基因还可以通过慢病毒感染引入单细胞阶段胚胎,为标准前列腺注射提供有效的替代,特别是在胚胎结构更具挑战性的物种或菌株中。在这种方法中,含有慢病毒载体的悬浮液被注射到受精大鼠胚胎的围网空间,从技术上讲,这种胚胎要求较低,成功率也更高。线病毒载体被证明有效地将转基因纳入基因组,以确定稳定的转基因系的产生。尽管存在一些局限性(例如,生物安全等级 2 要求、DNA 片段大小限制),但慢病毒转基因是一种快速而高效的转基因方法。此外,使用雌性大鼠与肥沃的男性菌株交配,具有不同的显性毛皮颜色,作为产生伪孕养母的替代品。
Introduction
多年来,实验室啮齿动物,如大鼠和老鼠,一直被用来模拟人类的生理和病理状况。动物研究已经导致发现,这是无法通过任何其他手段。最初,基因研究侧重于分析自发发生的疾病和表型,被认为是密切模仿人类的条件1。基因工程方法的发展允许引入或删除特定基因来获得所需的表型。因此,转基因动物的产生被认为是现代研究中的一项基本技术,它允许对生物体内基因功能的研究。
转基因动物技术通过在实验胚胎学和分子生物学方面取得的成绩而成为可能。20世纪60年代,波兰胚胎学家A.K.Tarkowski发表了关于小鼠胚胎在发育早期阶段操纵的第一个工作。此外,分子生物学家还开发了产生DNA载体(即载体)的技术,以便除其他外将外来DNA引入动物的基因组。这些载体允许传播选定的基因及其适当的修饰,这取决于所进行的研究类型。"转基因动物"一词是由戈登和陆克文3引入的。
第一个被广泛接受的物种,用于神经生物学,生理学,药理学,毒理学和许多其他领域的生物和医学是挪威大鼠,拉图斯诺维吉库斯4。然而,由于难以操纵大鼠胚胎,室内小鼠肌肉已成为基因研究中的主要动物物种。在这种研究中,老鼠占主导地位的另一个原因是,有胚胎干细胞技术可以为这种物种生产淘汰动物。最常见的转基因技术(相对于所有出生的动物,转基因后代的2-10%)是将DNA片段微入受精卵母细胞的亲核中。1990年,这种方法首次引入小鼠体内,适用于66、7只大鼠。与小鼠相比,前列腺注射的鼠转基因具有效率低8的特点,这与弹性等离子体和前列腺膜的存在密切相关。虽然操纵后的胚胎存活率比小鼠低40-50%,但这项技术被认为是转基因大鼠10代的一个标准。研究了能够保证有效转基因结合和提高注射酶的存活率的替代方法。
稳定转基因表达和传染给后代的关键决定因素是它与宿主细胞基因组的集成。伦迪病毒(LV)具有能够感染分裂细胞和非分裂细胞的显著特征。它们用作将异质基因纳入胚胎的工具被证明是高效的11,而转基因个体的特点是结合DNA片段的稳定表达。扁病毒载体的疗效已确认为小鼠12、13、13大鼠12、14,14等11种进行基因改造。12在这种方法中,LV悬浮液在两个原核的阶段被注射在胚胎的zona pellucida下。这种技术基本上保证了胚胎100%的存活率,因为奥莱玛仍然不受影响。生产高质量和相对集中的LV悬架是关键因素。然而,低浓度的LV悬浮液可以通过重复注射11来克服,这增加了蛋表面的病毒颗粒量,同时不影响膜的整合。接受反复注射到围场的胚胎进一步发育,转基因后代可以通过生殖系传播转基因。扁病毒转变转基因大鼠的生成效率高达80%12。
在这里,我们描述了HIV-1衍生的重组性扁病毒的产生,这种病毒是伪型的,带有光眼口腔炎病毒(VSV)G包络蛋白。使用第二代包装系统 VSV 伪型决定了病毒颗粒的广泛感染性,并允许生产高度稳定的载体,这些载体可以通过超离心和低温保存进行浓缩。滴定验证后,载体可作为将基因转化为白化病的威斯塔大鼠酶体的工具。经过一系列的注射后,胚胎可以培养过夜,并在双细胞阶段转移到寄养母亲。在这一点上,可以考虑两种替代方法之一。标准程序利用伪怀孕女性作为胚胎接受者。然而,当与输精管结扎的雄配后怀孕率较低时,胚胎可以植入怀孕的 Wistar / Sprague - Dawley ( SD )雌性,这些雌性与具有深色毛皮颜色的肥沃雄性大鼠交配(例如,棕色挪威 [ BN ] 大鼠)。毛皮的颜色允许区分后代与自然怀孕的后代,这些后代来自转移的操纵胚胎。
Subscription Required. Please recommend JoVE to your librarian.
Protocol
病毒载体的生产和应用符合生物安全二级准则,并经波兰环境部批准。下面描述的所有实验动物程序都得到当地伦理委员会的批准。这些动物被安置在稳定的温度(21~23°C)和湿度(50~60%)的单独通风的笼子里。在12 h/12 h的光/暗循环下,可以获得水和食物。
1. 伦迪病毒载体生产
- HEK 293T 细胞的转染
注:本文介绍的协议是为20×10厘米培养皿的转染而设计的,这些培养皿产生约200 mL的粗矢量上清液。- 培养DMEM介质中的HEK 293T细胞,在37°C的加湿CO2培养箱中补充胎儿牛血清(10%,v/v)。对于转染,准备直径为20 10厘米的板,并播种1.5⁄2 x 106 HEK 293T细胞每道菜。
- 当汇合达到+70%时,使用聚二烯胺(PEI)试剂(pH 7.0)转染细胞,每1μgDNA的PEI比例为3μg。
- 为五个盘子准备转染混合物(根据菜肴总数准备重复次数)。到1 mL的Dulbecco的改良鹰中(DMEM;无血清),加入三个质粒的混合物,使其达到最终量25μg的VSVg质粒,50μg的增量R8.2和50μg的编码质粒。
- 上下移液,在浓度为3μg/μL时加入125μL的PEI,在室温下孵育15分钟,在孵育过程中反转管子三次。每片加入200 μL的转染混合物。接下来,在37°C的加湿CO2培养箱中孵育板。
- 慢病毒载体的浓度
- 转染48小时后,收获含有LV颗粒的介质。使用 50 mL 锥形管。
注:当使用带有荧光标记的质粒时,此时可以可视化细胞以验证转染效率。可以添加 DMEM 介质的新部分,细胞可以孵育 24 小时。转染后在 48 和 72 小时时间点采集时,LV 屈服率可比较。 - 将介质在 3,000 x g下离心 5 分钟,室温可去除分离的细胞。
- 过滤上清液 (0.45 μm), 并将其倒入新管中。
注: 可以省略此步骤。 - 加入DNase I(无RNase,1 μg/mL)和MgCl2(1 mM),并在37°C的水浴中孵育15分钟。
- 将介质转移到一次性聚乙烯管,在摆动转子中以 115,000 x g和 4 °C 将超离心机转移到 1.5 h。
- 离心后,轻轻地从介质残留物中排干管壁。
- 用无菌磷酸盐缓冲盐水浸泡颗粒(PBS;每管70~80μL)。
- 在4~8°C下孵育2小时。
- 通过温和的移液重新悬浮PBS中的病毒载体。
注意:避免发泡。 - 在 7,000 x g 和 4 °C 下以 7,000 x g和 4 °C 将上清液转移到新管中,将上清器转移到新管中。重复此步骤,直到看不到细胞碎屑颗粒。
- 在-80°C下进行等值和冻结。避免重新冻结 LV 等号。
- 转染48小时后,收获含有LV颗粒的介质。使用 50 mL 锥形管。
- 利用定量聚合酶链反应确定病毒定位器
注:病毒载体的滴定使用定量PCR(qPCR)进行。该方法基于在病毒基因组15的长终端重复区域内放大双链84bp长DNA片段。- 通过对 LV 编码质粒进行序列稀释来准备标准曲线:1:500、1:1,000、1:5,000、1:10,000、1:100,000 和 1:1,000,000。确定用于标准曲线的质粒的副本数。使用以下公式:拷贝数/μL = (浓度 [g/μL] x 6.02 x 1023 [数字/摩尔]) / (660 [g/mol] x 质粒大小 [bp]),其中 6.02 x 1023个数字/mol 是 Avogadro 的数字,660 克/摩尔是 bp 重量。
注:可以使用在线拷贝编号计算器。 - 准备扁病毒悬浮液的稀释:1:100、1:500和1:1,000。
- 制备反应混合物(每孔体积):10 μL qPCR主混合物,1 μL 10 μM正向底漆,1 μL 10 μM反向底漆,7 μL H2O. 将混合物移制成96孔板的孔中。
注:前进引注:5'-AGCTTGCCTTGTGCTTTCA。反向引物:5'TGACTAAAAGGGTGGGA。 - 加入1 μL的每个标准稀释和扁病毒悬浮液三联。
- 根据以下参数运行qPCR:50°C为2分钟,96°C为5分钟,35个周期为96°C,为20秒,60°C为40s,70°C为1分钟,后为熔体曲线阶段:95°C为1分钟,60°C为30秒。
- 通过将每次稀释时收到的分子数与标准曲线进行比较,分析结果。确定矢量分子的浓度,作为每次稀释的三个复制的平均值。
注:呈现的定量给出了病毒颗粒的物理浓度。它不应被视为功能性手数。
- 通过对 LV 编码质粒进行序列稀释来准备标准曲线:1:500、1:1,000、1:5,000、1:10,000、1:100,000 和 1:1,000,000。确定用于标准曲线的质粒的副本数。使用以下公式:拷贝数/μL = (浓度 [g/μL] x 6.02 x 1023 [数字/摩尔]) / (660 [g/mol] x 质粒大小 [bp]),其中 6.02 x 1023个数字/mol 是 Avogadro 的数字,660 克/摩尔是 bp 重量。
2. 转基因大鼠的生成
- 受精胚胎的超级排卵和采集
- 管理促性腺激素。
注:为了增加采集的胚胎数量(每个女性大约30个),使用未成熟的5周大的Wistar雌性进行荷尔蒙刺激。- 在第1天(下午12时至下午1时),腹内注射怀孕母马血清促腺激素(PMSG;每名女性25 IU)。通过在0.9%NaCl中溶解激素粉末,在浓度为125 IU/mL时制备1 mL工作溶液。在 -20 °C 下储存长达 1 个月或 -80 °C,长达 6 个月。
- 在第3天(下午12时至下午1时),腹内注射人类胆囊性促性腺激素(hCG;每名女性30 IU)。通过在0.9%NaCl 中溶解激素粉末,制备 1 mL 工作溶液 (150 IU/mL) 的等位物。在 -20 °C 下储存长达 1 个月或 -80 °C,长达 6 个月。
- hCG管理后,与性繁殖的男性(3-10个月大)交配女性1:1。
- 第二天早上(第4天,上午8点到10点),检查女性是否存在阴道塞。检查阴道开口是否存在白色交配插头,为了获得最佳效果,应在交配之夜后清晨检查该插头。对于胚胎收集,只使用带有可见插头的女性。
- 上午10点收集胚胎。牺牲动物去切除排卵管,并在带有预热的M2介质的菜肴中收集卵管。
- 将排卵转移到35毫米的菜,含有预加热的M2介质,从牛睾牛睾口中含有透明质酶,浓度为0.5毫克/mL。
- 在立体显微镜下用细钳打开输卵管壁,按压安普拉(即,含有被积细胞包围的受精胚胎的排卵的肿胀部分),直到胚胎被释放。
注:透明质酶酶消化积细胞,释放胚胎。
注意:长时间接触透明质酶对胚胎有害;因此,此步骤不应超过 5 分钟。 - 为了便于从积细胞中释放胚胎,使用连接到口操作吸气管的玻璃转移移液器轻轻上下移液。
- 要产生转移移液器,将玻璃巴斯德移液器拉过火焰,以产生直线 ±5-10 厘米的尖端。打破移液器留下 ±4 厘米的尖端。
- 在M2介质中洗涤胚胎几次,去除透明质酶和细胞碎屑。将胚胎转移到含有(±50 μL)预平衡M16介质(液体石蜡或矿物油覆盖)的60毫米培养皿中,在加湿的37°C培养箱中,具有5%的CO2气氛。
- 管理促性腺激素。
- 将慢病毒载体微注射到佐纳盆下的单细胞阶段胚胎
注:使用带有两个可见原核的单细胞阶段胚胎进行微注射(图1)。- 在室温下解冻LV等位,在10,000 x g和RT下解冻2分钟,以颗粒任何剩余的细胞碎片。
- 微注射设置
- 使用微锻造准备玻璃持有移液器(硼硅酸盐玻璃毛细管)。将玻璃毛细管拉过火焰,产生 5-10 厘米的尖端。打破移液器留下 ±4 厘米的尖端。外径应为 ±80±120 μm。
注:确保移液器尖端完全笔直且光滑。 - 将拉拔的移液器用加热灯丝前面的尖端组装在微锻件中。将灯丝加热到离移液器尖端非常近,使其缩小到直径为±15 μm(大约胚胎大小的20%)。将移液器垂直放置在加热灯丝上,距离移液器尖端 2*3 mm,并开始加热。玻璃会变软的。加热,直到达到15°角。
- 使用移液器拉拔器使用灯丝制备微注射硼硅酸盐玻璃毛细管。将毛细管插入拉室。运行斜坡测试(首次用于新玻璃,每次更换灯丝后)。将热设置为斜坡值 -10,拉至 100,速度设置为 150,将时间设置为 100。
注:修改参数以获得最佳的注塑毛细管。 - 在生物安全层流罩下,使用微装载机尖端将大约 2 μL 的病毒溶液加载到微注射移液器中。
- 准备一个微注射盘(盖子60毫米培养皿),100 μL M2介质(中间)滴,由液体石蜡或矿物油覆盖。
- 将装有病毒溶液的保持移液器和微注射毛细血管安装在倒置显微镜下,将病毒溶液装入微操纵器和微注射盘。
- 使用微锻造准备玻璃持有移液器(硼硅酸盐玻璃毛细管)。将玻璃毛细管拉过火焰,产生 5-10 厘米的尖端。打破移液器留下 ±4 厘米的尖端。外径应为 ±80±120 μm。
- 执行微注射。
- 将15⁄20个单细胞阶段胚胎转移到微注射盘上的M2滴。使用保持移液器握住胚胎。
- 使用 400 倍放大倍率,使用连接到自动喷油器的玻璃毛细管将 Zona pellucida 下的 LV 溶液注入到围注素空间。将毛细血管放在佐纳贝卢维萨下一会儿。
注:使用温和的正压,病毒溶液将持续流出注射毛细血管,但输送的悬浮液的体积无法控制。 - 使用精细移液器,在5%CO2的温度下将胚胎返回到培养皿中的培养皿37°C。2一个zygote的注射次数可能有所不同,可以根据病毒载体浓度进行调整。
注:注射的胚胎可以在单细胞阶段移植给寄养母亲,或在两细胞阶段移植之前,在M16介质中孵育O/N。应避免大鼠胚胎的长时间体外培养。
- 将注射胚胎移植给养母
- 第 3 天(用于在单细胞阶段转移胚胎)或第 4 天(用于在双细胞阶段转移胚胎)与受精期雄配性成熟的 SD 雌性或输精管切除术 SD 雄性(输精管切除术程序如下第 3 节所述)。
注:对于排卵管转移,使用0.5天后共和 (dpc) 女性。 - 第二天早上,检查SD女性阴道塞,只使用那些有可见插头的。
- 进行胚胎移植。
注:在立体显微镜下使用无菌仪器进行外科手术。手术前,高压灭菌剪刀、细钳子、针夹和手术刀支架。- 用氯胺酮(50毫克/千克)和麦德多米丁(0.5毫克/千克)溶液给雌性麻醉。在开始外科手术之前,检查反射以确认麻醉。
- 用胆碱酸(2毫克/千克),但甲酸(1毫克/千克)和苯丙酮(5~10毫克/千克)注射,以防止炎症、疼痛和感染。
- 在双眼上涂抹眼药膏润滑,防止角膜干燥。从背部刮毛,用手术擦洗消毒皮肤,然后用无菌非粘附垫对皮肤进行70%的酒精消毒。让皮肤干燥。
- 在切口部位用100 μL 0.25%的bupivacain(局部麻醉剂)给动物下皮注射。在手术显微镜下,将易发位置的动物转移到加热垫上的清洁表面。用无菌窗帘盖住大鼠,在下背部切开一个小孔。
- 执行大约 2 厘米的皮肤切口,平行于腰椎柱。
- 用锋利的剪刀,在腹壁上切开一下。用钳子抓住卵巢脂肪垫,拉出卵巢和排卵管,并把它们放在用0.9%NaCl润湿的纱布上。
- 吸气M2介质,三泡空气,并将胚胎移植到毛细血管中。建议移植的胚胎总数(单边或双边):怀孕女性(+15-16个胚胎)、假怀孕女性(+30个胚胎)。
- 使用微剪刀在卵管(在支水管和安普拉拉之间)中做一个小切口,并将转移移液器插入卵管中。
- 将胚胎和气泡从移液器中轻轻排出到卵管中。用钝力将生殖道放回腹腔。
- 用多甘油酸可吸收缝合腹壁,用伤口夹闭合皮肤切口。根据可用的胚胎数量,对其他排卵器重复此过程。
- 在腹内注射阿提帕梅索(0.5毫克/千克),以扭转麻醉的效果。
- 将动物转移到一个干净的笼子里,并放在加热板上,从麻醉中完全恢复。大鼠分娩发生在21天后。
注:当雄性BN大鼠用于交配时,只有白鼠是潜在的转基因;棕色的小狗来自自然怀孕。 - 收集组织片段(最好是从耳朵)到基因型3周大的小狗。
- 第 3 天(用于在单细胞阶段转移胚胎)或第 4 天(用于在双细胞阶段转移胚胎)与受精期雄配性成熟的 SD 雌性或输精管切除术 SD 雄性(输精管切除术程序如下第 3 节所述)。
3. 血管切除术
注:手术前,高压灭菌剪刀、细钳子和针夹。
- 麻醉5周大雄性SD大鼠与氯胺酮(50毫克/千克)和麦德多米丁(0.5毫克/千克)溶液的i.p.给大鼠。在开始外科手术之前,检查反射以确认麻醉。
- 分别施用托芬纳酸(2毫克/千克),但亚诺醇酸(1毫克/千克)和苯丙酮(5~10毫克/千克)下皮,以防止炎症、疼痛和感染。
- 在双眼上涂抹眼药膏润滑,防止角膜干燥。将大鼠的软体放在加热垫上的干净表面上,用手术擦洗对睾酮上的皮肤进行消毒,然后用无菌非粘附垫对70%的酒精进行消毒。让皮肤干燥。用无菌窗帘盖住大鼠,在睾口上切上一个小孔。轻轻按压腹部,露出阴囊中的睾丸。
- 使用手术剪刀,在阴囊中间做±0.5厘米切口。在测试点之间找到中线墙(尖线)。
- 在靠近中线墙左侧的睾膜中进行 5 mm 切口。
- 小心地将睾牛向左推,并将血管延迟(在睾比和中线之间)定位为具有单个血管的白色管道。
- 使用制表师的钳子轻轻将血管从阴囊中拉出。用一对钳子握住血管延迟,用细剪刀(或用第二对钳子的红热提示烧焦)将其切割。拆下管道的 ±1 厘米碎片。
注:如果执行烧灼,则将火焰中第二对钳的尖端保持。 - 对其他睾测试重复上述步骤。用多甘油酸可吸收缝合缝合皮肤,用阿提帕美酮(0.5毫克/千克)注射动物腹内注射。
- 将大鼠放在一个干净的笼子里,放在加热板上,直到动物从麻醉中恢复过来。
注:在±2周的恢复期后,雄性可用于测试交配。确认不育后,可用于伪妊娠诱导。
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
使用本文所述的协议,产生了携带Syn-TDP-43-eGFP构造的慢病毒载体(物理LV定位 = 3.4 x 108/μL),然后可用于单细胞阶段胚胎子区注射。只有具有两个可见原核的胚胎接受手术。注射病毒悬浮液的数量是实验性的。高植入效率以及同时缺乏转基因后代被认为是病毒颗粒数量不足以成功转导的指标。在这种情况下,注射次数增加。LV的单一管理导致20只F0代大鼠出生,其中没有一只是转基因的。注射次数增加一个数量级并没有导致大鼠的出生,而是100%的胚胎发育到双细胞阶段。在随后的实验中,与获得后代的价值相比,注射次数增加了一次。对于两次注射的变种,有8只大鼠出生,其中3只被证实携带转基因(表1概述)。其中一位创始人没有将转基因转移到后代身上。每个实验变种中注入和转移的胚胎数量为48个,变异为LV x1和LV x2,LV x10为45。每个实验设置都使用了三个寄养女性。所选择的方法允许生成稳定的转基因大鼠线,在整个中枢神经系统(图2A,B)14中,在神经元Synapsin-1促进剂的控制下表达TDP-43-eGFP融合蛋白。14基于伦迪病毒的转变导致一个基因的拷贝插入,如qPCR(图2C)所示。
在上述实验设置中,被注射胚胎的存活率为95%。当对表2中概述的其他慢病毒载体使用相同的方法时,也得出了类似的结果。在前列腺注射中幸存下来的胚胎比例要低得多(29~45%)。表2总结了受操纵的酶的植入效率的代表性结果,考虑了假怀孕与怀孕女性的转移。此前有报道称,将非操纵胚胎与注射胚胎一起使用。我们的总体结果表明,怀孕的雌性大鼠可以作为具有同等效率的养母。我们在怀孕和伪怀孕大鼠中植入外来胚胎的比例相似(几个实验装置的总体平均值:15%对16%)。然而,当胚胎进行更微妙的操作时,植入率较高,这意味着亚区注射(10%对21%)。值得注意的是,对个别注射子弹进行分析的数字数据表明,植入的有效性取决于一个胚胎(表1,最后一列)的注射次数,间接取决于病毒载量。
| 向量 | 注射/胚胎数量 | 注射的胚胎数量 | 小狗数量 | 养母人数 | 转基因创始人数量 | 每种变种的植入效率 |
| Syn-TDP-43WTLV | 1 | 48 | 20 | 3 | 0 | 42% |
| 10 | 45 | 0 | 3 | 0 | 0% | |
| 2 | 48 | 8 | 3 | 3 | 17% |
表1:使用Syn-TDP-43WTWT慢病毒载体的酶注射子分区注射次数摘要。
| 方法 | 向量 | 蒂特/浓度 | 注射胚胎数量 | 存活的胚胎 | 成活率 | 养母人数 | 小狗数量 | 植入效率 | 妊娠 (P) /伪怀孕 (PP) |
| PNI | TTYH1-Thy1-EGFP | 1 纳克/μL | 1083 | 424 | 39% | 16 | 54 | 13% | Pp |
| PNI | H3m樱桃 | 0.5-2 纳克/μL | 2229 | 647 | 29% | 29 | 67 | 10% | Pp |
| PNI | Syn-TDP-43-A315T | 2 纳克/μL | 1256 | 562 | 45% | 31 | 42 | 7% | Pp |
| 低压 | Syn-TDP-43-A315T | 8.7 x 108 | 115 | 106 | 92% | 7 | 18 | 17% | P |
| 低压 | Syn-TDP-43 WT | 3.4 x 108 | 152 | 141 | 93% | 9 | 28 | 20% | P |
| 低压 | LVH3mcherry | 1.3 x 107 | 504 | 450 | 89% | 13 | 115 | 26% | Pp |
表2:胚胎存活率和植入效率,取决于所使用的注射方法和怀孕与假妊娠的诱导。PNI,亲核注入;LV,扁病毒载体亚区注射。

图1:为亚区小病毒载体注射准备的单细胞级大鼠胚胎的微观照片。胚胎被固定的移液器固定。两个含有母体和父系遗传物质和极性身体的原核是可见的。刻度柱 = 20 μm。请点击此处查看此图形的较大版本。
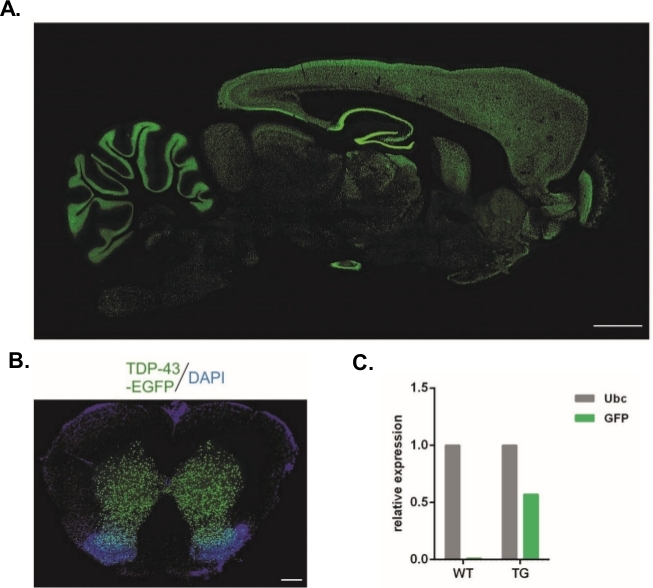
图2:生成稳定的转基因大鼠线,在整个中枢神经系统的神经元突触酶-1促进剂的控制下表达TDP-43-eGFP融合蛋白。(A) 突触酶-1 (Syn) 驱动的 hTDP-43-eGFP 表达模式,位于转基因大鼠大脑的下垂部分。鳞杆 = 3 毫米 (B) 转基因大鼠脊髓的冠状部分,其中 eGFP 荧光,反染色 DAPI,仅限于脊髓的灰质。刻度柱 = 250 μm。(C) GFP转基因转基因转录与泛素C参考笔录的相对表达。n = 2 种野生类型。n = 2 转基因。这个数字从14修改。请点击此处查看此图形的较大版本。
Subscription Required. Please recommend JoVE to your librarian.
Discussion
转基因技术的进步使啮齿动物模型成为生物医学研究的宝贵工具。它们提供了研究体内基因型-表型关系的机会。在这里,我们提出了一种广泛可用的替代常规转基因通过前核注入。使用慢病毒基因转导绕过了要求苛刻的微注射的需要,因为病毒载体可以在zona pellucida下注射。这种方法不影响胚胎的完整性,这基本上保证了注射酶的100%存活率。通过慢病毒载体结合的转基因可稳稳地融入宿主基因组,允许长期表达和生殖系传播。此外,我们还提出了两种替代技术,用于为寄养母亲进行胚胎移植。一项技术利用胚胎移植给伪怀孕的雌性,这些雌性以前是通过与输精管化不孕雄配而准备的。另一种技术是基于使用自然怀孕的雌性,与肥沃的雄配,但与不同的毛皮颜色(即 BN 大鼠)。这种更生理的怀孕过程允许胚胎的正确发育,经过具有挑战性的基因改造16。
1990年报道了首次成功产生转基因大鼠的尝试。然而,由于鼠转变困难,近几十年来产生的转基因大鼠线相对较少。使用微注射观察到小鼠和大鼠的转基因几大差异。对于大鼠,主要用于异种线(例如,Wistar 和 SD)用于变性。对于小鼠,研究人员主要使用F1杂交的近亲繁殖菌株,因为它们的生育能力较高,对荷尔蒙超排卵的反应更好,并且相对容易地从单细胞阶段发育到胚泡18。大鼠超排卵的诱导比使用标准PMSG/hCG激素刺激的小鼠效率低得多。因此,已经尝试开发替代协议,以管理这些激素的老鼠利用连续FSH输注,而不是一个单一的PMSG管理19。然而,由PMSG/hCG或FSH/hCG引起的超排卵已被证明具有可比的效率20。我们认为,影响超排效果的最关键因素是选定女性的年龄。然而,应测试每个大鼠菌株、实验室等的确切参数。
将DNA溶液注入单细胞胚胎的proNUCLEUS的过程对两种啮齿动物都相似。然而,大鼠酶的正核没有像小鼠那样的常规形状,而且往往在细胞的细胞质中更难定义。此外,大鼠酶细胞膜和原核膜具有更弹性和粘性,从而使装有DNA溶液的玻璃微移液的插入复杂化。这些因素导致注射后大鼠卵子存活率降低(小鼠为31-65%对80%),并解释大鼠9的转基因效率较低。此外,对胚胎的密集、机械操作也会影响植入效率,在许多实验室,包括我们的实验室,植入效率最高可达10%。即使在植入适当数量的胚胎21后,观察这个相对较低的产率。
克服上述困难的一种方法是感染单细胞胚胎感染逆转录病毒。逆转录病毒含有RNA形式的遗传物质,在进入受感染细胞时,通过病毒的逆转录酶转录成DNA。然后,DNA通过核孔输送到细胞核,以亲病毒的形式融入细胞的基因组。伦特病毒载体已被用来产生转基因小鼠和大鼠12,14,22。12,14,22缺乏佐纳骨盆的单细胞胚胎可以用慢病毒载体孵育在溶液中,也可以将载体注射在佐那骨盆下进入腹腔空间。该方法的主要优点是效率极高,达到80%以上的转基因后代。感染慢病毒载体后,不同部位的许多拷贝可能集成到酶基因组中,而通过前列腺微注射的转生方法则相反,其中通常观察到一个整合位点12。在使用慢病毒载体制成的转基因创始人的后代中,转基因的单个副本被分离,这可能表现为每个后代的转基因的不同表达特征。但是,这会增加从转基因派生所需的表达配置文件接收受试者的机会。这些限制主要适用于转基因的大小,其范围为约8 kb23。
大鼠转基因的另一个困难是一代女性作为转基因胚胎的代孕母亲。在标准程序中,女性与无菌输精管化雄叉,以诱导伪怀孕。在大鼠中,伪怀孕评估技术比小鼠困难得多,因此,在与雄配前几天,有时使用促性腺激素释放激素激动剂的刺激。出于这些原因,在所述协议中,我们提供了两种替代方法,以获得养母。在使用怀孕或伪怀孕女性时,纵的酶体的整体植入效率是相似的。然而,自然、非操纵的胚胎的存在以及纵的胚胎可以提高怀孕率16 。虽然植入率的主要区别是操纵技术(即PNI与LV,10%对20%;见表2),使用假怀孕女性作为寄养母亲可能有利于一些实验。
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
作者(W.K.)拥有波兰共和国专利局专利权"转基因动物生产方法"(号P 355353;21.03.2008)。
Acknowledgments
这项研究得到了ANIMOD项目的支持,该项目由波兰科学基金会团队技术核心设施加方案内支持,该项目由欧洲联盟在欧洲区域发展基金下共同资助。
Materials
| Name | Company | Catalog Number | Comments |
| 7500 Real Time PCR System | Applied Biosystems | ||
| Aerrane (isoflurane) | Baxter | FDG9623 | |
| Aspirator tube assemblies for calibrated microcapillary pipettes | Sigma | A5177-5EA | |
| Atipam 5 mg/ml | Eurovet Animal Health BV | N/A | 0.5 mg/kg |
| Baytril 25 mg/ml (enrofloksacin) | Bayer | N/A | 5-10 mg/kg |
| Borosilicate glass capillaries with filament GC100TF-15 | Harvard Apparatus Limited | 30-0039 | injection capillary |
| Bupivacaine 25 mg/ml | Advanz Pharma | N/A | 0.25% in 0.9% NaCl |
| Butomidor 10 mg/ml (butorphanol tartrate) | Orion Pharma | N/A | 1 mg/kg |
| CELLSTAR Tissue Cell Culture Dish 35-mm | Greiner Bio-One | 627160 | |
| CELLSTAR Tissue Cell Culture Dish 60-mm | Greiner Bio-One | 628160 | |
| CellTram Oil | Eppendorf | 5176 000.025 | |
| Cepetor (Medetomidine) 1 mg/ml | cp-pharma | N/A | 0.5 mg/kg |
| Chorulon, Human Chorionic Gonadotrophin | Intervet | N/A | 150 IU/ ml ml 0.9% NaCl |
| DMEM low glucose | Sigma Aldrich | D6048 | |
| DNase, RNase-free | A&A Biotechnology | 1009-100 | |
| EmbryoMax Filtered Light Mineral Oil | Sigma | ES-005-C | |
| Envelope protein coding plasmid for lentiviral vectors (VSVg plasmid) | ADDGENE | 14888 | |
| FemtoJet | Eppendorf | 4i /5252 000.013 | |
| Fetal Bovine Serum | Sigma Aldrich | F9665-500ML | |
| Folligon, Pregnant Mare’s Serum Gonadotropin | Intervet | N/A | 125 IU/ml in .9% NaCl |
| HEK 293T cells | ATCC | ATCC CRL-3216 | |
| Hyaluronidase from Bovine Testis | Sigma | H4272-30MG | 0.5 mg/ml in M2 medium |
| Inverted Microscope | Zeiss | Axiovert 200 | |
| Ketamine 100mg/ml | Biowet Pulawy | N/A | 50 mg/kg |
| Liquid Paraffin | Merck Millipore | 8042-47-5 | |
| M16 medium EmbryoMax | Sigma | MR-016-D | |
| M2 medium | Sigma | M7167 | |
| Magnesium Chloride 1M | Sigma Aldrich | 63069-100ML | |
| Microforge | Narishige | MF-900 | |
| Mineral Oil | Sigma | M8410-500ML | |
| NaCl 0.9% | POLPHARMA OTC | N/A | sterile, 5ml ampules |
| Operation microscope | Inami Ophthalmic Instruments | Deca-21 | |
| Packaging system coding plasmid for lentiviral vectors (delta R8.2 plasmid) | ADDGENE | 12263 | |
| PEI reagent (Polyethylenimine, Mw ~ 25,000,), | Polysciences, Inc | 23966-1 | |
| Penicilin-streptomycin | Sigma Aldrich | P0781-100ML | |
| Phosphate Buffered Saline, pH 7.4, liquid, sterile-filtered, suitable for cell culture | Sigma Aldrich | 806552-500ML | |
| Puller | Sutter Instrument Co. | P-97 | |
| Reflex Clip Applier/Reflex Clips | World Precision Instruments | 500345/500346 | |
| Safil, polyglycolic acid, braided, coated, absorbable threads | B.Braun Surgical | 1048029 | |
| Stereomicroscope | Olympus | SZX16 | |
| Surgical Sewing Thread | B.Braun | C1048040 | |
| SYBR Green PCR Master Mix | Applied Biosystem | 4334973 | |
| Tolfedine 4% (tolfenamic acid) | Vetoquinol | N/A | 2 mg/kg |
| TransferMan NK2 | Eppendorf | N/A | |
| Trypsin EDTA solution | Sigma Aldrich | T3924-500ML | |
| Ultracentrifuge | Beckman Coulter | Optima L-100 XP | |
| VacuTip | Eppendorf | 5175108.000 | holders capillary |
| Vita-POS | Ursapharm | N/A | eye ointment |
| Warming Plate | Semic | N/A | |
| Watchmaker Forceps | VWR | 470018-868 |
References
- Lazar, J., Moreno, C., Jacob, H. J., Kwitek, A. E. Impact of genomics on research in the rat. Genome Research. 15 (12), 1717-1728 (2005).
- Tarkowski, A. K. Studies on mouse chimeras developed from eggs fused in vitro. National Cancer Institute Monographs. 11, 51-71 (1963).
- Gordon, J. W., Ruddle, F. H. Integration and stable germ line transmission of genes injected into mouse pronuclei. Science. 214 (4526), 1244-1246 (1981).
- Gill, T. J., Smith, G. J., Wissler, R. W., Kunz, H. W. The Rat as an Experimental Animal. Science. 245 (4915), 269-276 (1989).
- Aitman, T. J., et al. Progress and prospects in rat genetics: a community view. Nature Genetics. 40 (5), 516-522 (2008).
- Hammer, R. E., Maika, S. D., Richardson, J. A., Tang, J. P., Taurog, J. D. Spontaneous inflammatory disease in transgenic rats expressing HLA-B27 and human beta 2m: an animal model of HLA-B27-associated human disorders. Cell. 63 (5), 1099-1112 (1990).
- Mullins, J. J., Peters, J., Ganten, D. Fulminant hypertension in transgenic rats harbouring the mouse Ren-2 gene. Nature. 344 (6266), 541-544 (1990).
- Menoret, S., Remy, S., Usal, C., Tesson, L., Anegon, I. Generation of Transgenic Rats by Microinjection of Short DNA Fragments. Rat Genomics: Methods and Protocols. 597, 81-92 (2010).
- Tesson, L., et al.
- Charreau, B., Tesson, L., Soulillou, J. P., Pourcel, C., Anegon, I. Transgenesis in rats: Technical aspects and models. Transgenic Research. 5 (4), 223-234 (1996).
- Ritchie, W. A., Neil, C., King, T., Whitelaw, C. B. Transgenic embryos and mice produced from low titre lentiviral vectors. Transgenic Research. 16 (5), 661-664 (2007).
- Lois, C., Hong, E. J., Pease, S., Brown, E. J., Baltimore, D. Germline transmission and tissue-specific expression of transgenes delivered by lentiviral vectors. Science. 295 (5556), 868-872 (2002).
- Pfeifer, A., Ikawa, M., Dayn, Y., Verma, I. M. Transgenesis by lentiviral vectors: lack of gene silencing in mammalian embryonic stem cells and preimplantation embryos. Proceedings of the National Academy of Sciences of the United States of America. 99 (4), 2140-2145 (2002).
- Koza, P., et al. Neuronal TDP-43 depletion affects activity-dependent plasticity. Neurobiology of Disease. 130, 104499 (2019).
- Scherr, M., Battmer, K., Blomer, U., Ganser, A., Grez, M. Quantitative determination of lentiviral vector particle numbers by real-time PCR. Biotechniques. 31 (3), 520 (2001).
- Canseco, R. S., et al. Gene transfer efficiency during gestation and the influence of co-transfer of non-manipulated embryos on production of transgenic mice. Transgenic Research. 3 (1), 20-25 (1994).
- Charreau, B., Tesson, L., Soulillou, J. P., Pourcel, C., Anegon, I. Transgenesis in rats: technical aspects and models. Transgenic Research. 5 (4), 223-234 (1996).
- Brinster, R. L., Chen, H. Y., Trumbauer, M. E., Yagle, M. K., Palmiter, R. D. Factors affecting the efficiency of introducing foreign DNA into mice by microinjecting eggs. Proceedings of the National Academy of Sciences of the United States of America. 82 (13), 4438-4442 (1985).
- Armstrong, D. T., Opavsky, M. A. Superovulation of immature rats by continuous infusion of follicle-stimulating hormone. Biology of Reproduction. 39 (3), 511-518 (1988).
- Popova, E., Krivokharchenko, A., Ganten, D., Bader, M. Comparison between PMSG- and FSH-induced superovulation for the generation of transgenic rats. Molecular Reproduction and Development. 63 (2), 177-182 (2002).
- Johnson, L. W., Moffatt, R. J., Bartol, F. F., Pinkert, C. A. Optimization of embryo transfer protocols for mice. Theriogenology. 46 (7), 1267-1276 (1996).
- van den Brandt, J., Wang, D., Kwon, S. H., Heinkelein, M., Reichardt, H. M. Lentivirally generated eGFP-transgenic rats allow efficient cell tracking in vivo. Genesis. 39 (2), 94-99 (2004).
- Remy, S., et al. The Use of Lentiviral Vectors to Obtain Transgenic Rats. Rat Genomics: Methods and Protocols. 597, 109-125 (2010).
