

Summary
Denne artikel har til formål at give metoden for lentiviral transgenese i rotte embryoner ved hjælp af flere injektioner af en virus suspension i zygote perivitelline rummet. Kvindelige rotter, der er parret med en frugtbar mandlig stamme med en anden dominerende pels farve bruges til at generere pseudopregnant plejemødre.
Abstract
Transgene dyremodeller er grundlæggende vigtige for moderne biomedicinsk forskning. Inkorporeringen af fremmede gener i tidlige muse- eller rotteembryoner er et uvurderligt redskab til analyse af genfunktionen i levende organismer. Standardtransgenesemetoden er baseret på mikroindsprøjtning af udenlandske DNA-fragmenter i en pronucleus af en befrugtet oocyt. Denne teknik er meget udbredt i mus, men er stadig relativt ineffektiv og teknisk krævende hos andre dyrearter. Transgenet kan også indføres i et-trins embryoner via lentiviral infektion, hvilket giver et effektivt alternativ til standard pronuclear injektioner, især hos arter eller stammer med en mere udfordrende embryo struktur. I denne fremgangsmåde sprøjtes en suspension, der indeholder lentivirale vektorer, ind i perivitelline-rummet hos et befrugtet rottefoster, som er teknisk mindre krævende og har en højere succesrate. Lentivirale vektorer viste sig effektivt at indarbejde transgenet i genomet for at bestemme dannelsen af stabile transgene linjer. Trods visse begrænsninger (f.eks. biosikkerhedsniveau 2-krav, DNA-fragmentstørrelsesgrænser) er lentiviral transgenese en hurtig og effektiv transgenesemetode. Derudover, ved hjælp af kvindelige rotter, der er parret med en frugtbar mandlig stamme med en anden dominerende pels farve præsenteres som et alternativ til at generere pseudopregnant plejemødre.
Introduction
I mange år har laboratoriegnavere, såsom rotter og mus, været brugt til at modellere menneskelige fysiologiske og patologiske forhold. Dyreforsøg har ført til opdagelser, der var uopnåelige på nogen anden måde. I første omgang fokuserede genetiske undersøgelser på analyse af spontant forekommende lidelser og fænotyper, der anses for nøje at efterligne den menneskelige tilstand1. Udviklingen af genteknologiske metoder gjorde det muligt at indføre eller slette specifikke gener for at opnå en ønsket fænotype. Derfor er generation af transgene dyr anerkendt som en grundlæggende teknik i moderne forskning, der tillader undersøgelser af genfunktion i levende organismer.
Transgene dyreteknologi er blevet mulig gennem en kombination af resultater inden for eksperimentel embryologi og molekylærbiologi. I 1960'erne offentliggjorde den polske embryolog A. K. Tarkowski det første værk om museembryosmanipulation i de tidlige udviklingsstadier2. Derudover udviklede molekylærbiologer teknikker til at generere DNA-vektorer (dvs. bærere) til bl.a. Disse vektorer tillader udbredelse af udvalgte gener og deres passende modifikation, afhængigt af hvilken type forskning der udføres. Udtrykket "transgene dyr" blev indført af Gordon og Ruddle3.
Den første bredt accepterede arter, der blev brugt i neurobiologi, fysiologi, farmakologi, toksikologi, og mange andre områder af biologiske og medicinske videnskaber var Norge rotte, Rattus norvegicus4. Men på grund af vanskelighederne med at manipulere rotte embryoner, huset musen Mus muskulahar blevet den dominerende dyrearter i genetisk forskning5. En anden grund til musens forrang i en sådan forskning var tilgængeligheden af embryonal stamcelleteknologi til at generere knockout dyr til denne art. Den mest almindeligt anvendte teknik transgenese (2-10% af transgene afkom i forhold til alle fødte dyr) er mikroinjektion af DNA-fragmenter i en pronucleus af en befrugtet oocyt. I 1990 blev denne fremgangsmåde, som først blev indført i mus, tilpasset til rotter6,7. Rotte transgenese ved pronuclear injektion er karakteriseret ved lavere effektivitet8 sammenlignet med mus, som er strengt relateret til tilstedeværelsen af elastisk plasma og pronukleare membraner9. Selv om overlevelsen af embryoner efter manipulation er 40-50% lavere end hos mus, denne teknik betragtes som en standard i dannelsen af genetisk modificerede rotter10. Alternative tilgange, der kan garantere effektiv transgeniblanding og højere overlevelsesrater for injicerede zygoter, er blevet undersøgt.
Den vigtigste faktor for stabil transgene udtryk og overførsel til afkom er dens integration i værtscellegenomet. Lentivira (LVs) har det karakteristiske træk at være i stand til at inficere både dividere og ikke-dividere celler. Deres anvendelse som et redskab til inkorporering af heterologe gener i embryoner viste sig at være yderst effektiv11, og transgene individer er kendetegnet ved stabil teksion af det inkorporerede DNA-fragment. Effekten af lentivirale vektorer er blevet bekræftet til genetisk modifikation af mus12,13, rotter12,14og andre arter11. Ved denne metode injiceres LV-suspensionen under embryonets zona pellucida på stadiet af to pronuclei. Denne teknik væsentlige garanterer 100% overlevelse af embryoner, fordi oolemma forbliver upåvirket. Produktionen af forholdsvis koncentrerede LV-suspensioner af høj kvalitet er afgørende faktorer. Lavere koncentrationer af LV-suspensioner kan dog overvindes ved gentagne injektioner11, hvilket øger mængden af viruspartikler på ægoverfladen, mens det ikke påvirker membranintegrationen. Embryoner, der udsættes for gentagne injektioner i perivitellinrummet, udvikler sig yderligere, og transgene afkom kan overføre transgene gennem kønskiminen. Effektiviteten af transgene rotte generation af lentiviral transgenese kan være så højt som 80%12.
Her beskriver vi produktionen af HIV-1-afledt rekombinant lentivirus, der blev pseudotyped med vesikulær stomatitisvirus (VSV) G kuvertprotein. Brugen af anden generations emballeringssystem VSV pseudotype bestemmer den brede infektivitet af viruspartikler og tillader produktion af meget stabile vektorer, der kan koncentreres ved ultracentrifugering og kryopræpareret. Efter titer verifikation, vektorerne er klar til at blive brugt som et køretøj til transgene levering i albino Wistar rotte zygotes. Efter en række injektioner, kan embryonerne dyrkes natten over og overføres på to-celle stadium til plejemødre. På dette tidspunkt kan en af to alternative tilgange overvejes. Standardproceduren udnytter pseudogravide hunner som embryomodtagere. Men når graviditetsraten er lav efter parring med vasektomerede hanner, kan embryonerne implanteres i gravide Wistar/Sprague-Dawley (SD) hunner, der er parret med frugtbare hanrotter med en mørk pelsfarve (f.eks. Farven på pelsen tillader sondringen af afkom fra naturlig graviditet fra afkom, der stammer fra de overførte manipulerede embryoner.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
Produktionen og anvendelsen af virale vektorer var i overensstemmelse med retningslinjerne for biosikkerhedsniveau 2 og blev godkendt af det polske miljøministerium. Alle forsøgsdyreprocedurer, der er beskrevet nedenfor, blev godkendt af det lokale etiske udvalg. Dyrene blev anbragt i individuelt ventilerede bure ved en stabil temperatur (21-23 °C) og fugtighed (50-60%) med ad libitum adgang til vand og mad under en 12 h/12 h lys / mørk cyklus.
1. Lentiviral vektorproduktion
- Transfection af HEK 293T celler
BEMÆRK: Den protokol, der præsenteres heri, er designet til transfection af tyve Ø10 cm kulturretter, der producerer ca. 200 ml rå vektorsupernatant.- Dyrkning HEK 293T celler i DMEM medium, der suppleres med føtal kvæg serum (10%, v/ v) i en befugtet CO2 inkubator ved 37 °C. Til transfection, forberede tyve 10 cm diameter plader, og frø 1,5-2 x 106 HEK 293T celler pr fad.
- Når sammenløbet når ~70%, transfect cellerne ved hjælp af polyethylenimin (PEI) reagens, pH 7.0, med et forhold på 3 μg PEI pr 1 μg DNA.
- Klargør transfektionsblandingen til fem plader (forbered antallet af gentagelser i henhold til det samlede antal retter). Til 1 ml Af Dulbecco's Modified Eagle Medium (DMEM; uden serum), tilsæt blandingen af tre plasmider, så de når en endelig mængde på 25 μg VSVg plasmid, 50 μg delta R8.2 og 50 μg kodning plasmid.
- Pipetterop og ned, og der tilsættes 125 μL PEI i en koncentration på 3 μg/μL. Inkuber ved stuetemperatur i 15 minutter, og røret vendes tre gange under inkubationen. Der tilsættes 200 μL af transfektionsblandingen pr. plade. Derefter inkuberes pladerne i2 en befugtet CO 2-inkubator ved 37 °C.
- Koncentration af lentivirale vektorer
- 48 timer efter transfection, høste mediet, der indeholder LV partikler. Brug 50 ml koniske rør.
BEMÆRK: Når du bruger en plasmid med en fluorescerende tag, kan cellerne visualiseres på dette punkt for at kontrollere transfection effektivitet. En ny del af DMEM-mediet kan tilføjes, og celler kan inkuberes i yderligere 24 timer. LV-udbyttet er sammenligneligt, når det opsamles på 48- og 72-timers-tidspunkterne efter transfection. - Mediet centrifugeres ved 3.000 x g i 5 min. og stuetemperatur for at fjerne afmonterede celler.
- Supernatanten (0,45 μm) filtreres, og det hældes i nye rør.
BEMÆRK: Dette trin kan udelades. - Der tilsættes DNase I (RNase-fri, 1 μg/ml) og MgCl2 (1 mM) og inkuberes i et vandbad ved 37 °C i 15 min.
- Mellem- til engangspolyethylenrør overføres, og ultracentrifugen overføres i en svingende rotor ved 115.000 x g og 4 °C i 1,5 timer.
- Efter centrifugering drænes rørenes vægge forsigtigt fra de mellemstore rester.
- Iblødsætning pellet med sterile fosfat-buffered saltvand (PBS; 70-80 μL per rør).
- Inkuber i 2 timer ved 4-8 °C.
- Resuspender de virale vektorer i PBS ved skånsom pipettering.
FORSIGTIG: Undgå skumdannelse. - Der overføres til et centrifugerør på 1,5 ml og centrifugeved 7.000 x g og 4 °C i 30 s. Overfør supernatanten til et nyt rør. Gentag dette trin, indtil ingen cellulære snavs pellet er synlig.
- Aliquot og fryse ved -80 °C. Undgå at fryse LV aliquot'en igen.
- 48 timer efter transfection, høste mediet, der indeholder LV partikler. Brug 50 ml koniske rør.
- Bestemmelse af virustitre ved hjælp af kvantitativ polymerasekædereaktion
BEMÆRK: Titrering af virale vektorer udføres ved hjælp af kvantitativ PCR (qPCR). Denne metode er baseret på at forstærke et dobbeltstrenget 84 bp langt DNA-fragment inden for det lange terminalgentagelsesområde i det virale genom15.- Forbered standardkurven ved at foretage serielle fortyndinger af LV-kodningplasmid: 1:500, 1:1.000, 1:5.000, 1:10.000, 1:100.000 og 1:1.000.000. Bestem antallet af kopier af plasmid, der bruges til standardkurven. Brug følgende formel: antal kopier/μL = (koncentration [g/μL] x 6,02 x 1023 [tal/mol]) / (660 [g/mol] x plasmid størrelse [bp]), hvor 6,02 x 1023 tal/mol er Avogadros nummer, og 660 g/mol er bp-vægt.
BEMÆRK: Der kan anvendes online kopinummerberegnere. - Fortyndinger af lentiviral suspension: 1:100, 1:500 og 1:1,000.
- Der fremstilles reaktionsblandingen (volumenpr. brønd): 10 μL qPCR Mastermix, 1 μL af 10 μM Forward primer, 1 μL af 10 μM omvendt primer og 7 μL H2O. Pipette blandingen i brøndene på 96-brøndplader.
BEMÆRK: Fremadprimer: 5'-AGCTTGCCTTGAGTGCTTCA. Omvendt primer: 5'-TGACTAAAAGGGTCTGAGGGA. - Der tilsættes 1 μL af hver standardfortynding og lentiviral suspension i tre eksemplarer.
- QPCR køres efter følgende parametre: 50 °C i 2 min, 96 °C i 5 min og 35 cyklusser på 96 °C i 20 s, 60 °C i 40 s og 70 °C i 1 min efterfulgt af smeltekurvetrin: 95 °C i 1 min. og 60 °C ved 30 s.
- Analysér resultaterne ved at sammenligne antallet af molekyler, der modtages for hver fortynding, med standardkurven. Koncentrationen af vektormolekyler bestemmes som gennemsnittet af tre replikater for hver fortynding.
BEMÆRK: Den præsenterede kvantificering giver den fysiske koncentration af de virale partikler. Det bør ikke behandles som en funktionel titer.
- Forbered standardkurven ved at foretage serielle fortyndinger af LV-kodningplasmid: 1:500, 1:1.000, 1:5.000, 1:10.000, 1:100.000 og 1:1.000.000. Bestem antallet af kopier af plasmid, der bruges til standardkurven. Brug følgende formel: antal kopier/μL = (koncentration [g/μL] x 6,02 x 1023 [tal/mol]) / (660 [g/mol] x plasmid størrelse [bp]), hvor 6,02 x 1023 tal/mol er Avogadros nummer, og 660 g/mol er bp-vægt.
2. Generation af transgene rotter
- Superovulation og indsamling af befrugtede embryoner
- Administrere gonadotropiner.
BEMÆRK: For at øge antallet af indsamlede embryoner (ca. 30 pr. kvinde) skal du bruge umodne 5 uger gamle Wistar-hunner til hormonbehandling.- På dag 1 (12 PM-1 PM), inperitoneally injicere gravid hoppens serum gonadotropin (PMSG; 25 IE per kvinde). Der fremstilles 1 ml arbejdsopløsning ved en koncentration på 125 IE/ml ved at opløse hormonpulver i 0,9% NaCl. Opbevares ved -20 °C i op til 1 måned eller -80 °C i op til 6 måneder.
- På dag 3 (12 PM-1 PM), intraperitoneally injicere menneskelige choriongonadotropin (hCG; 30 IE per kvinde). Der fremstilles 1 ml prøveaf arbejdsopløsning (150 IE/ml) ved at opløse hormonpulver i 0,9% NaCl. Opbevares ved -20 °C i op til 1 måned eller -80 °C i op til 6 måneder.
- Efter hCG administration, parre hunner 1:1 med seksuelt frugtbare hanner (3-10 måneder gammel).
- Den næste morgen (dag 4 på 8-10 AM), kontrollere hunnerne for tilstedeværelsen af en vaginal stik. Kontroller vaginal åbning for tilstedeværelsen af en hvidlig parring stik, som for bedste visualisering bør kontrolleres tidligt om morgenen efter parring nat. Til embryonsamling må hunner med synligt stik kun anvendes.
- Saml embryoner kl. Ofre dyrene til punktafgifter oviducts, og indsamle oviducts i en skål med forvarmet M2 medium.
- Oviducts overføres til en 35 mm skål, der indeholder forvarmet M2-medium med hyaluronidase fra kvægtestikler i en koncentration på 0,5 mg/ml.
- Åbn væggene i oviduct ved hjælp af fine pincet under et stereomikroskop og tryk på ampulla (dvs. den hævede del af oviduct, der indeholder befrugtede embryoner, der er omgivet af cumulus celler), indtil embryonerne er befriet.
BEMÆRK: Hyaluronidase enzymatisk fordøjer cumulus celler, frigive embryoner.
FORSIGTIG: Langvarig udsættelse for hyaluronidase er skadelig for embryoner; derfor bør dette trin ikke vare længere end 5 min. - For at lette frigivelsen af embryoner fra cumulus celler, forsigtigt pipette dem op og ned ved hjælp af en glasoverførsel pipette, der er forbundet til en mund-opererede aspirator rør.
- For at producere overførslen pipette, trække et glas Pasteur pipette over en flamme til at producere en lige ~ 5-10 cm tip. Bryd pipetten og efterlad en ~4 cm spids.
- Vask embryonerne et par gange i M2 medium for at fjerne hyaluronidase og cellulære snavs. Embryonerne overføres til en 60 mm skål, der indeholder (~50 μL) dråber forekvilibreret M16-medium, dækket af flydende paraffin eller mineralsk olie, i en befugtet 37 °C-inkubator med en 5% CO2-atmosfære.
- Administrere gonadotropiner.
- Mikroinjektion af lentivirale vektorer til et-celle-trins embryo under zona pellucida
BEMÆRK: Brug embryoner i et celletrin med to synlige pronukler til mikroinjektion (figur 1).- Tø LV aliquot ved stuetemperatur og centrifuge ved 10.000 x g og RT i 2 min til pellet eventuelle resterende cellulære snavs.
- Opsætning af mikroinjektion
- Forbered glas bedrift pipetter (borosilikat glas kapillær) ved hjælp af en mikroforge. Træk glaskapillær over en flamme for at producere en 5-10 cm spids. Bryd pipetten og efterlad en ~4 cm spids. Den udvendige diameter skal være ~80-120 μm.
BEMÆRK: Sørg for, at pipettspidsen er helt lige og glat. - Saml den trak pipette i en mikroforge med spidsen foran varmeglødetråden. Glødetråden opvarmes meget tæt på pipettespidsen, og lad den krympe til en diameter på ~15 μm (ca. 20 % af embryonens størrelse). Anbring pipettten vinkelret på varmeglødetråden, 2-3 mm fra pipettespidsen, og begynd at varme. Glasset vil blødgøre. Varm, indtil den når en 15° vinkel.
- Forbered mikroinjektion borosilikat glas kapillærer med en glødetråd ved hjælp af en pipette puller. Sæt kapillæren i trækkammeret. Kør en rampetest (første gang for nyt glas og hver gang efter udskiftning af glødetråden). Indstil varmen til rampeværdien -10, Træk til 100, Hastighed til 150 og Tid til 100.
BEMÆRK: Der skal ændres parametrene for at opnå en optimal injektionskapillær. - Under en biosikkerhedshætte til laminar belastning lægges ca. 2 μL af den virale opløsning i mikroinjektionspipetten med en mikroloaderspids.
- Der fremstilles en mikroinjektionsskål (låg på 60 mm petriskål) med en dråbe M2-medium på 100 μL (i midten), dækket af flydende paraffin eller mineralsk olie.
- Monter den holdende pipette og mikroinjektionkapillar, der er fyldt med viral opløsning på en mikromanipulator og mikroinjektionsskål under et omvendt mikroskop.
- Forbered glas bedrift pipetter (borosilikat glas kapillær) ved hjælp af en mikroforge. Træk glaskapillær over en flamme for at producere en 5-10 cm spids. Bryd pipetten og efterlad en ~4 cm spids. Den udvendige diameter skal være ~80-120 μm.
- Udfør mikroinjektionen.
- 15-20 et-trins embryoner overføres til M2-dråben på mikroinjektionsskålen. Hold fosteret ved hjælp af en holdepipette.
- Ved hjælp af 400x forstørrelse, injicere LV opløsning under zona pellucida til perivitelline rummet ved hjælp af glaskapillary, der er forbundet til en automatisk injektor. Hold kapillær under zona pellucida et øjeblik.
BEMÆRK: Ved hjælp af et let positivt tryk vil den virale opløsning flyde kontinuerligt ud af injektionskapillaret, men mængden af den leverede suspension kan ikke kontrolleres. - Brug en fin pipette til at returnere embryonerne til dyrkningsskålen i rugemaskinen2 ved 37 °C i en 5% CO 2-atmosfære. Antallet af injektioner af en zygote kan variere og kan tilpasses på grundlag af den virale vektorkoncentration.
BEMÆRK: De injicerede embryoner kan overføres til plejemødre på encellestadiet eller inkuberes O/N i M16-medium, før de overføres i tocellestadiet. Langvarig in vitro-kultur af rotteembryoner bør undgås.
- Overførsel af injicerede embryoner til plejemødre
- Forbered plejemødre ved parring af kønsmodne SD-hunner med frugtbare BN-hanner eller med vasektomiserede SD-hanner (vasektomiproceduren er beskrevet i afsnit 3 nedenfor) på dag 3 (til overførsel af embryoner på encellestadiet) eller dag 4 (til overførsel af embryoner på tocellestadiet).
BEMÆRK: Til oviduct overførsel, bruge 0,5 dage efter coitum (dpc) kvinder. - Den næste morgen, kontrollere SD hunner for en vaginal stik, og bruge kun dem med et synligt stik.
- Udfør embryooverførsel.
BEMÆRK: Udfør den kirurgiske procedure med sterile instrumenter under et stereomikroskop. Før dagen for operationen, autoklave saks, fine pincet, nål indehaveren, og skalpel indehaveren.- Anæstesi en kvinde med i.p. administration af ketamin (50 mg/kg) og medetomidinopløsning (0,5 mg/kg). Test for reflekser for at bekræfte anæstesi før du starter den kirurgiske procedure.
- Dyret indsprøjtes subkutant med tolefenamissyre (2 mg/kg), menorphanoltartrat (1 mg/kg) og enrofloksacin (5-10 mg/kg) for at forebygge henholdsvis betændelse, smerte og infektion.
- Påfør oftalmologiske salvesmøring på begge øjne for at forhindre hornhindetørring. Barber pelsen fra bagsiden, og steriliser huden med kirurgisk krat efterfulgt af 70% alkohol ved hjælp af sterile ikke-overholde puder. Lad huden tørre.
- Dyret indsprøjtes subkutant med 100 μL 0,25% bupivacaine (lokalbedøvelse) på incisionsstedet. Overfør dyret i en udsat position til en ren overflade på en varmepude under formålet med et kirurgisk mikroskop. Dæk rotten med en steril drapere med et lille hul skåret over nederste del af ryggen.
- Udfør et ca. 2 cm hudsnit parallelt med lændehvirvelsøjlen.
- Brug skarpe saks, lave et snit i bugvæggen. Grab en æggestokkene fedt pad ved hjælp af pincet, og trække sig ud af æggestokkene og oviduk og placere dem på gaze, der er bevåd med 0,9% NaCl.
- Aspirate M2 medium, tre bobler af luft, og embryoner i overførslen kapillær. Anbefalet samlet antal embryoner, der skal overføres (ensidig eller bilateral): gravid kvinde (≤ 15-16 embryoner), pseudogravid kvinde (≤ 30 embryoner).
- Lav et lille snit i oviduket (mellem infundibulum og ampulla) ved hjælp af mikro-saks, og indsæt overførslen pipette i oviduct.
- Udvis forsigtigt embryoner og luftbobler fra pipetteringen til oviduket. Med stumpe pincet, placere forplantningsevnen tilbage i bughulen.
- Sutur bugvæggen med polyglykolsyre absorberbare suturer og lukke huden snit med sårclips. Afhængigt af antallet af embryoner, der er til rådighed, gentagedenne procedure for den anden oviduct.
- Der indsprøjtes dyret intraperitonealt med atipamezole (0,5 mg/kg) for at vende virkningen af anæstesi.
- Overfør dyret til et rent bur og opbevar det på en varmeplade for at komme sig helt efter bedøvelsen. Levering hos rotter sker efter ~ 21 dage.
BEMÆRK: Når han-BN-rotter anvendes til parring, er det kun hvide unger, der er potentielt transgene; brune hvalpe er fra naturlig graviditet. - Vævsfragmenter (helst fra øret) indsamles til genotype 3-ugers gamle unger.
- Forbered plejemødre ved parring af kønsmodne SD-hunner med frugtbare BN-hanner eller med vasektomiserede SD-hanner (vasektomiproceduren er beskrevet i afsnit 3 nedenfor) på dag 3 (til overførsel af embryoner på encellestadiet) eller dag 4 (til overførsel af embryoner på tocellestadiet).
3. Vasektomi
BEMÆRK: Før dagen for operationen, autoklave saks, fine pincet og nål indehaveren.
- Anæstesi en 5 uger gammel han-SD-rotte med i.p. administration af ketamin (50 mg/kg) og medetomidinopløsning (0,5 mg/kg). Test for reflekser for at bekræfte anæstesi før du starter den kirurgiske procedure.
- Administrere tolfenamic syre (2 mg/kg), butorphanol tartrat (1 mg/kg), og enrofloksacin (5-10 mg/kg) subkutant for at forhindre betændelse, smerte og infektion, henholdsvis.
- Påfør oftalmologiske salvesmøring på begge øjne for at forhindre hornhindetørring. Placer rotte supine på en ren overflade på en varmepude, og sterilisere huden på testiklerne med kirurgisk krat efterfulgt af 70% alkohol ved hjælp af sterile ikke-overholde puder. Lad huden tørre. Dæk rotten med en steril drapere med et lille hul skåret over testiklerne. Tryk forsigtigt på maven for at eksponere testiklerne i sakimten.
- Brug kirurgisk saks, lave en ~ 0,5 cm snit i midten af sakrotale sæk. Find midterlinjen væg (hvidlig linje) mellem testiklerne.
- Lav et 5 mm snit i testismembranen tæt på venstre side af midtervæggen.
- Skub forsigtigt testiklerne til venstre og find vas deferens (mellem testiklerne og midterlinjen) som en hvid kanal med et enkelt blodkar.
- Træk forsigtigt vas deferens ud af scrotal sac ved hjælp af en urmager's pincet. Hold vas deferens med et par pincet, og skær det med fine saks (eller ætse med rødglødende spidser af et andet par pincet). Fjern et ~1 cm fragment af kanalen.
BEMÆRK: Hvis ætsning ender, skal du holde spidsen af det andet par pincet i flammen. - Gentag ovenstående procedure for de andre testikler. Sutur huden med polyglykolsyre absorberbare suturer og injicere dyret intraperitoneally med atipamezole (0,5 mg/kg).
- Anbring rotten i et rent bur på en varmeplade, indtil dyret kommer sig efter bedøvelse.
BEMÆRK: Hanner kan anvendes i testparringerne efter en ~ 2-ugers restitutionsperiode. Efter sterilitet er bekræftet, de kan bruges til pseudograviditet induktion.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Ved hjælp af den protokol, der er beskrevet heri, blev der produceret udlånte vektorer, der bar Syn-TDP-43-eGFP-konstruktionen (fysisk LV-titer = 3,4 x 108/μL), og derefter kunne de anvendes til subzonale injektioner i ét celletrin. Kun embryoner med to synlige pronuclei er blevet underkastet proceduren. Antallet af injektioner af virale suspensioner blev bestemt eksperimentelt. Høj implantationseffektivitet og samtidig mangel på transgene afkom blev betragtet som indikatorer for et utilstrækkeligt antal viruspartikler til vellykket transduktion. I dette tilfælde blev antallet af injektioner øget. Den enkelte administration af LV resulterede i fødslen af 20 F0-generation rotter, hvoraf ingen var transgene. En stigning i antallet af injektioner med én størrelsesorden resulterede ikke i fødslen af rotter, men 100% af embryonerne udviklede sig til tocellestadiet. I efterfølgende forsøg blev antallet af injektioner øget med én sammenlignet med den værdi, som afkommet blev opnået for. For den variant af to injektioner blev der født otte rotter, hvoraf tre blev bekræftet til at bære transgene (sammenfattet i tabel 1). En af grundlæggerne overfører ikke transgenet til afkom. Antallet af embryoner, der blev injiceret og overført i hver forsøgsvariant, var 48 i varianter LV x1 og LV x2 og 45 i LV x10. Tre plejehunner blev brugt til hver forsøgsopstilling. Den valgte fremgangsmåde gjorde det muligt at skabe stabile transgene rottelinjer, der udtrykte TDP-43-eGFP-fusionsproteinet under kontrol af den neuronale Synapsin-1-promotor i hele centralnervesystemet (figur 2A,B)14. Lentivirus-baserede transgenese resulterede i en enkelt kopi indsættelse af transgene som påvist ved qPCR (Figur 2C).
I den eksperimentelle opsætning, der er beskrevet ovenfor, var overlevelsesraten for de injicerede embryoner 95%. Lignende resultater blev opnået , da den samme metode blev anvendt for andre lentivirale vektorer som sammenfattet i tabel 2. Procentdelen af embryoner, der overlevede de pronukleare injektioner, var betydeligt lavere (29-45 %). Tabel 2 opsummerer de repræsentative resultater af implantationseffektiviteten hos manipulerede zygoter i betragtning af overførslen af pseudogravide mod drægtige kvinder. Der blev tidligere rapporteret om anvendelse af ikke-manipulerede embryoner sammen med injicerede embryoner16. Vores samlede resultater tyder på, at gravide hunrotter kan bruges som plejemødre med tilsvarende effektivitet. Vi opnåede en lignende procentdel af implantation af udenlandske embryoner hos drægtige og pseudopregnant rotter (samlet gennemsnit for flere eksperimentelle opsætninger: 15% vs. 16%). Men implantationshastigheden var højere, når embryonerne gennemgik mere subtil manipulation, hvilket betyder en subzonal injektion (10% vs. 21%). Navnlig viste de numeriske data, der blev analyseret for de enkelte mikroinjektionsrunder, at implantationens effektivitet afhang af antallet af injektioner af et embryon(tabel 1, sidste kolonne) og indirekte afhang af viral belastning.
| Vektor | antal injektioner/embryoner | antal indsprøjtede embryoner | antal unger | antallet af plejemødre | antal transgene stiftere | Implantationseffektivitet for hver variant |
| Syn-TDP-43WTLV | 1 | 48 | 20 | 3 | 0 | 42% |
| 10 | 45 | 0 | 3 | 0 | 0% | |
| 2 | 48 | 8 | 3 | 3 | 17% |
Tabel 1: Oversigt over antallet af subzonale injektioner af zygoter med Syn-TDP-43WTlentivirale vektorer.
| Metode | Vektor | Titer/ Koncentration | Antal injicerede embryoner | Overlevede embryoner | Overlevelsesraten | Antal plejemødre | Antal unger | Implantationseffektivitet | Graviditet (P) /Pseudopregnancy (PP) |
| PNI (PNI) | TTYH1-Thy1-EGFP | 1 ng/μL | 1083 | 424 | 39% | 16 | 54 | 13% | S |
| PNI (PNI) | H3mKirsebær | 0,5-2 ng/μL | 2229 | 647 | 29% | 29 | 67 | 10% | S |
| PNI (PNI) | Syn-TDP-43-A315T | 2 ng/μL | 1256 | 562 | 45% | 31 | 42 | 7% | S |
| Lv | Syn-TDP-43-A315T | 8,7 x 108 | 115 | 106 | 92% | 7 | 18 | 17% | P |
| Lv | Syn-TDP-43 WT | 3,4 x 108 | 152 | 141 | 93% | 9 | 28 | 20% | P |
| Lv | LVH3mkirsebær | 1,3 x 107 | 504 | 450 | 89% | 13 | 115 | 26% | S |
Tabel 2: Embryooverlevelsesrate og implantationseffektivitet, afhængigt af den anvendte injektionsmetode og graviditet versus pseudograviditetinduktion. PNI, pronuklear injektion; LV, lentiviral vektor subzonal injektion.

Figur 1: Mikroskopisk fotografi af et-celle-trins rotte embryo, der blev forberedt til subzonal lentiviral vektor injektion. Fosteret blev immobiliseret med en holdepipette. To pronuclei, der indeholdt moderlig og faderlig genetisk materiale og polære krop er synlige. Skalabjælke = 20 μm. Klik her for at se en større version af dette tal.
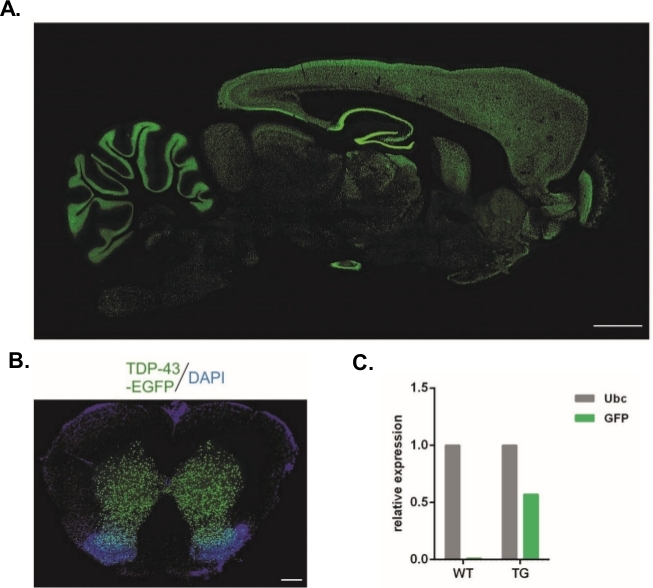
Figur 2: Generation af stabile transgene rottelinjer, der udtrykte TDP-43-eGFP-fusionsproteinet under kontrol af den neuronale Synapsin-1-promotor i hele centralnervesystemet. (A) Synapsin-1 (Syn)-drevet hTDP-43-eGFP ekspressionsmønster i en sagittal del af den transgene rottehjerne. Scale bar = 3 mm. (B) Koronal del af rygmarven af en transgen rotte, hvor eGFP fluorescens, imødegås med DAPI, var begrænset til grå stof af rygmarven. Skalabjælke = 250 μm. (C) Relativ udtryk for GFP transgene udskrift sammenlignet med ubiquitin C reference udskrift. n = 2 wildtype. n = 2 transgene. Tallet blev ændret fra14. Klik her for at se en større version af dette tal.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Fremskridt inden for transgene teknologier har gjort gnavere modeller et uvurderligt redskab i biomedicinsk forskning. De giver mulighed for at studere genotype-fænotype relationer in vivo. Her præsenterer vi et bredt tilgængeligt alternativ til konventionel transgenese ved pronuclear injektioner. Brugen af lentiviral gentransduktion omgår behovet for krævende mikroinjektioner, fordi virale vektorer kan injiceres under zona pellucida. Denne fremgangsmåde påvirker ikke embryonens integritet, som i det væsentlige garanterer en overlevelsesrate på 100 % for injicerede zygoter. Transgene, der er indarbejdet ved hjælp af lentivirale vektorer er stabilt integreret i værten genom, så langsigtede udtryk og germline transmission. Derudover præsenterer vi to alternative teknikker til modificeret embryooverførsel til plejemødre. En teknik udnytter embryo overførsel til pseudopregnant hunner, der tidligere er udarbejdet ved parring med vasectomized ufrugtbar hanner. Den anden teknik er baseret på brugen af naturligt gravide hunner, der er parret med frugtbare hanner, men med en anden pels farve (dvs. BN rotter). Denne mere fysiologiske forløb af graviditeten giver mulighed for korrekt udvikling af embryoner, der gennemgår udfordrende genetiske modifikationer16.
De første vellykkede forsøg på at generere transgene rotter blev rapporteret i 19907. På grund af vanskeligheder med rottetransgenesis17er der imidlertid i de senesteårtierblevet genereret et relativt lille antal transgene rottelinjer . Flere væsentlige forskelle er observeret mellem mus og rotte transgenesis ved hjælp af mikroinjektioner. For rotter anvendes hovedsagelig udavlede linjer (f.eks. For mus, forskere primært bruge F1 krydsning af indavlede stammer på grund af deres højere frugtbarhed, bedre respons på hormonelle superovulation, og relativt nem udvikling af embryoner in vitro fra en-celle fase til blastocyster18. Induktion af superovulation hos rotter er langt mindre effektiv end hos mus ved hjælp af standard PMSG/hCG hormon stimulation. Af denne grund, forsøg er blevet gjort for at udvikle alternative protokoller til at administrere disse hormoner i rotter, der udnytter kontinuerlig FSH infusion i stedet for en enkelt PMSG administration19. Det har imidlertid vist sig, at superovulation, der skyldes PMSG/hCG eller FSH/hCG, har en sammenlignelig effektivitet20. Efter vores mening er den mest kritiske faktor, der påvirker effektiviteten af superovulation en alder af udvalgte kvinder. Ikke desto mindre bør de nøjagtige parametre testes for hver rottestamme, laboratorium osv.
Proceduren for injektion af DNA-opløsning i pronucleus af et encellet embryo er den samme for begge gnavere arter. Men pronuclei af rotte zygoter ikke har sådanne regelmæssige former som i mus og har tendens til at være vanskeligere at definere i cytoplasmaet af cellen. Derudover er rottezygotcellemembranen og den pronukleare membran mere elastisk og tyktflydende, hvilket komplicerer indføringen af et glasmikropipette, der er fyldt med DNA-opløsning. Disse faktorer fører til lavere overlevelsesrater for rotteæg efter mikroinjektionen (31-65 % vs. 80 % hos mus) og forklarer den lavere transgeneseeffektivitet hos rotter9. Desuden kan intensiv, mekanisk manipulation af fosteret også påvirke implantationseffektiviteten, som i mange laboratorier, herunder vores, når op på maksimalt 10 %. Dette relativt lave udbytte observeres selv efter implantation af et passende antal embryoner21.
En metode, der overvinder ovennævnte vanskeligheder, er infektionen af encellede embryoner med retrovirus. Retrovirus indeholder genetisk materiale i form af RNA, som ved indtræden i den inficerede celle transskriberes til DNA ved omvendt transkriptase af virus. DNA'et transporteres derefter gennem de nukleare porer til cellekernen, hvor det integreres i cellens genom i form af en provirus. Lentivirale vektorer er blevet anvendt til at generere transgene mus og rotter12,14,22. Encellede embryoner, der mangler en zona pellucida kan inkuberes i en opløsning med en lentiviral vektor, eller vektoren kan injiceres under zona pellucida i perivitelline rummet. Den største fordel ved denne metode er dens ekstremt høje effektivitet, nå mere end 80% af transgene afkom. Efter infektion med lentiviral vektor kan mange kopier på forskellige steder integreres i zygote genomet i modsætning til transgenesemetoden ved pronuclear microinjection, hvor et integrationssted normalt observeres12. Hos afkom af den transgene grundlægger, der er lavet ved hjælp af lentivirale vektorer, adskilles individuelle kopier af transgenet, hvilket kan manifesteresig ved forskellige udtryksprofiler af transgenet i hvert af afkom. Dette kan dog øge chancen for at modtage et emne med den ønskede udtryksprofil, der er afledt af transgene. Restriktionerne gælder primært for størrelsen af transgene, som er begrænset til ca 8 kb23.
En anden vanskelighed i rotte transgenesis er den generation af hunner, der tjener som rugemødre for genetisk modificerede embryoner. I standardproceduren krydses hunnerne med sterile vasectomized hanner for at fremkalde pseudograviditet. Hos rotter er pseudopregnancy vurderingsteknikken meget vanskeligere end hos mus, så stimulering med gonadotropin frigivelse hormon agonist bruges nogle gange et par dage før parring med hanner. Af disse grunde, i den beskrevne protokol, vi giver to alternative tilgange til at opnå plejemødre. Den samlede implantationseffektivitet hos manipulerede zygoter, når der anvendes gravide eller pseudogravide hunner, er den samme. Men, tilstedeværelsen af naturlige, ikke-manipulerede embryoner sammen med manipulerede dem kan forbedre graviditet sats16. Selv om den største forskel i implantationshastigheden er manipulationsteknikken (dvs. Table 2
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Forfatteren (W.K.) har ret til patentet, "Metode til fremstilling af et transgent dyr," fra patentkontoret i Republikken Polen (nr. P 355353; 21.03.2008).
Acknowledgments
Denne undersøgelse blev støttet af ANIMOD-projektet inden for Team Tech Core Facility Plus-programmet under Foundation for Polish Science, der samfinansieres af EU under Den Europæiske Fond for Regionaludvikling til WK.
Materials
| Name | Company | Catalog Number | Comments |
| 7500 Real Time PCR System | Applied Biosystems | ||
| Aerrane (isoflurane) | Baxter | FDG9623 | |
| Aspirator tube assemblies for calibrated microcapillary pipettes | Sigma | A5177-5EA | |
| Atipam 5 mg/ml | Eurovet Animal Health BV | N/A | 0.5 mg/kg |
| Baytril 25 mg/ml (enrofloksacin) | Bayer | N/A | 5-10 mg/kg |
| Borosilicate glass capillaries with filament GC100TF-15 | Harvard Apparatus Limited | 30-0039 | injection capillary |
| Bupivacaine 25 mg/ml | Advanz Pharma | N/A | 0.25% in 0.9% NaCl |
| Butomidor 10 mg/ml (butorphanol tartrate) | Orion Pharma | N/A | 1 mg/kg |
| CELLSTAR Tissue Cell Culture Dish 35-mm | Greiner Bio-One | 627160 | |
| CELLSTAR Tissue Cell Culture Dish 60-mm | Greiner Bio-One | 628160 | |
| CellTram Oil | Eppendorf | 5176 000.025 | |
| Cepetor (Medetomidine) 1 mg/ml | cp-pharma | N/A | 0.5 mg/kg |
| Chorulon, Human Chorionic Gonadotrophin | Intervet | N/A | 150 IU/ ml ml 0.9% NaCl |
| DMEM low glucose | Sigma Aldrich | D6048 | |
| DNase, RNase-free | A&A Biotechnology | 1009-100 | |
| EmbryoMax Filtered Light Mineral Oil | Sigma | ES-005-C | |
| Envelope protein coding plasmid for lentiviral vectors (VSVg plasmid) | ADDGENE | 14888 | |
| FemtoJet | Eppendorf | 4i /5252 000.013 | |
| Fetal Bovine Serum | Sigma Aldrich | F9665-500ML | |
| Folligon, Pregnant Mare’s Serum Gonadotropin | Intervet | N/A | 125 IU/ml in .9% NaCl |
| HEK 293T cells | ATCC | ATCC CRL-3216 | |
| Hyaluronidase from Bovine Testis | Sigma | H4272-30MG | 0.5 mg/ml in M2 medium |
| Inverted Microscope | Zeiss | Axiovert 200 | |
| Ketamine 100mg/ml | Biowet Pulawy | N/A | 50 mg/kg |
| Liquid Paraffin | Merck Millipore | 8042-47-5 | |
| M16 medium EmbryoMax | Sigma | MR-016-D | |
| M2 medium | Sigma | M7167 | |
| Magnesium Chloride 1M | Sigma Aldrich | 63069-100ML | |
| Microforge | Narishige | MF-900 | |
| Mineral Oil | Sigma | M8410-500ML | |
| NaCl 0.9% | POLPHARMA OTC | N/A | sterile, 5ml ampules |
| Operation microscope | Inami Ophthalmic Instruments | Deca-21 | |
| Packaging system coding plasmid for lentiviral vectors (delta R8.2 plasmid) | ADDGENE | 12263 | |
| PEI reagent (Polyethylenimine, Mw ~ 25,000,), | Polysciences, Inc | 23966-1 | |
| Penicilin-streptomycin | Sigma Aldrich | P0781-100ML | |
| Phosphate Buffered Saline, pH 7.4, liquid, sterile-filtered, suitable for cell culture | Sigma Aldrich | 806552-500ML | |
| Puller | Sutter Instrument Co. | P-97 | |
| Reflex Clip Applier/Reflex Clips | World Precision Instruments | 500345/500346 | |
| Safil, polyglycolic acid, braided, coated, absorbable threads | B.Braun Surgical | 1048029 | |
| Stereomicroscope | Olympus | SZX16 | |
| Surgical Sewing Thread | B.Braun | C1048040 | |
| SYBR Green PCR Master Mix | Applied Biosystem | 4334973 | |
| Tolfedine 4% (tolfenamic acid) | Vetoquinol | N/A | 2 mg/kg |
| TransferMan NK2 | Eppendorf | N/A | |
| Trypsin EDTA solution | Sigma Aldrich | T3924-500ML | |
| Ultracentrifuge | Beckman Coulter | Optima L-100 XP | |
| VacuTip | Eppendorf | 5175108.000 | holders capillary |
| Vita-POS | Ursapharm | N/A | eye ointment |
| Warming Plate | Semic | N/A | |
| Watchmaker Forceps | VWR | 470018-868 |
References
- Lazar, J., Moreno, C., Jacob, H. J., Kwitek, A. E. Impact of genomics on research in the rat. Genome Research. 15 (12), 1717-1728 (2005).
- Tarkowski, A. K. Studies on mouse chimeras developed from eggs fused in vitro. National Cancer Institute Monographs. 11, 51-71 (1963).
- Gordon, J. W., Ruddle, F. H. Integration and stable germ line transmission of genes injected into mouse pronuclei. Science. 214 (4526), 1244-1246 (1981).
- Gill, T. J., Smith, G. J., Wissler, R. W., Kunz, H. W. The Rat as an Experimental Animal. Science. 245 (4915), 269-276 (1989).
- Aitman, T. J., et al. Progress and prospects in rat genetics: a community view. Nature Genetics. 40 (5), 516-522 (2008).
- Hammer, R. E., Maika, S. D., Richardson, J. A., Tang, J. P., Taurog, J. D. Spontaneous inflammatory disease in transgenic rats expressing HLA-B27 and human beta 2m: an animal model of HLA-B27-associated human disorders. Cell. 63 (5), 1099-1112 (1990).
- Mullins, J. J., Peters, J., Ganten, D. Fulminant hypertension in transgenic rats harbouring the mouse Ren-2 gene. Nature. 344 (6266), 541-544 (1990).
- Menoret, S., Remy, S., Usal, C., Tesson, L., Anegon, I. Generation of Transgenic Rats by Microinjection of Short DNA Fragments. Rat Genomics: Methods and Protocols. 597, 81-92 (2010).
- Tesson, L., et al.
- Charreau, B., Tesson, L., Soulillou, J. P., Pourcel, C., Anegon, I. Transgenesis in rats: Technical aspects and models. Transgenic Research. 5 (4), 223-234 (1996).
- Ritchie, W. A., Neil, C., King, T., Whitelaw, C. B. Transgenic embryos and mice produced from low titre lentiviral vectors. Transgenic Research. 16 (5), 661-664 (2007).
- Lois, C., Hong, E. J., Pease, S., Brown, E. J., Baltimore, D. Germline transmission and tissue-specific expression of transgenes delivered by lentiviral vectors. Science. 295 (5556), 868-872 (2002).
- Pfeifer, A., Ikawa, M., Dayn, Y., Verma, I. M. Transgenesis by lentiviral vectors: lack of gene silencing in mammalian embryonic stem cells and preimplantation embryos. Proceedings of the National Academy of Sciences of the United States of America. 99 (4), 2140-2145 (2002).
- Koza, P., et al. Neuronal TDP-43 depletion affects activity-dependent plasticity. Neurobiology of Disease. 130, 104499 (2019).
- Scherr, M., Battmer, K., Blomer, U., Ganser, A., Grez, M. Quantitative determination of lentiviral vector particle numbers by real-time PCR. Biotechniques. 31 (3), 520 (2001).
- Canseco, R. S., et al. Gene transfer efficiency during gestation and the influence of co-transfer of non-manipulated embryos on production of transgenic mice. Transgenic Research. 3 (1), 20-25 (1994).
- Charreau, B., Tesson, L., Soulillou, J. P., Pourcel, C., Anegon, I. Transgenesis in rats: technical aspects and models. Transgenic Research. 5 (4), 223-234 (1996).
- Brinster, R. L., Chen, H. Y., Trumbauer, M. E., Yagle, M. K., Palmiter, R. D. Factors affecting the efficiency of introducing foreign DNA into mice by microinjecting eggs. Proceedings of the National Academy of Sciences of the United States of America. 82 (13), 4438-4442 (1985).
- Armstrong, D. T., Opavsky, M. A. Superovulation of immature rats by continuous infusion of follicle-stimulating hormone. Biology of Reproduction. 39 (3), 511-518 (1988).
- Popova, E., Krivokharchenko, A., Ganten, D., Bader, M. Comparison between PMSG- and FSH-induced superovulation for the generation of transgenic rats. Molecular Reproduction and Development. 63 (2), 177-182 (2002).
- Johnson, L. W., Moffatt, R. J., Bartol, F. F., Pinkert, C. A. Optimization of embryo transfer protocols for mice. Theriogenology. 46 (7), 1267-1276 (1996).
- van den Brandt, J., Wang, D., Kwon, S. H., Heinkelein, M., Reichardt, H. M. Lentivirally generated eGFP-transgenic rats allow efficient cell tracking in vivo. Genesis. 39 (2), 94-99 (2004).
- Remy, S., et al. The Use of Lentiviral Vectors to Obtain Transgenic Rats. Rat Genomics: Methods and Protocols. 597, 109-125 (2010).
