The procedures described above allowed modeling the pathophysiological mechanisms underlying bacterial pneumonia-induced lung injury in mice. To begin to model, C57BL/6 wild-type (WT) mice were obtained from the Jackson Laboratory and bred at the institute's animal facility. Male WT C57BL/6 mice, 8 weeks old, underwent intratracheal inoculation of either TH broth (control), 3 x 106 CFU of live Spn, or 200 CFU of live Kpn. After the infection, the mice were monitored for 6 and 10 days for Spn and Kpn, respectively. Although the infected groups displayed lower body weight compared to the uninfected control, the Spn group recovered their body weight toward baseline, while Kpn-infected mice displayed slow recovery after 6 days of infection (Figure 2A). During the duration of the study, no mice required undergoing euthanasia due to body weight over 20%, and there was no evidence of pain and distress.
We measured lung injury over different intervals. BAL protein concentration and the total cell count for both the BAL and lungs were remarkedly higher in the infected groups (Figure 2). Representative histological sections displaying the inflammatory process in both models were obtained at day 2, 4, and 6 post-inoculation (Figure 3), showing evidence of persistent alveolar inflammation in the Kpn-infected mice (Figure 4), even at day 10. Kpn-infected mice continued the injury by day 10 (Figure 2 and Figure 4), while Spn-infected mice resolved the lung inflammation by day 6 (Figure 2 and Figure 3).
Lung single cell suspensions were used to discriminate the immune landscape by high-dimensional flow cytometry at day 6 post-infection in the Spn model, using an 18-color panel in fluorescence activated cell sorting (FACS). Using t-distributed stochastic neighbor embedding (t-SNE), overall differences in the immune cell composition can be visualized, being remarkable for an increased number of granulocytes (CD45+, CD11b+, CD24+, MHC-II-), interstitial macrophages (CD45+, CD11b+, MHC-II-, CD24-), monocytes (CD45+, CD11b+, MHC-II-, CD24-, CD64-), B cells (CD45+, CD19+), and T cells (CD45+, CD3+), including natural killer cells (CD45+, CD3+, NK1.1+), as displayed in Figure 5. Gating strategies are shown in Figure 6.

Figure 1: Surgical procedure for intratracheal instillation of live bacteria. (A) Position of the mouse in the sterile surgical area, hanging by the incisors. (B) Incision area and trachea exposure. (C) Intubation process inserting the 20 G catheter. Figure created with Biorender.com. Please click here to view a larger version of this figure.
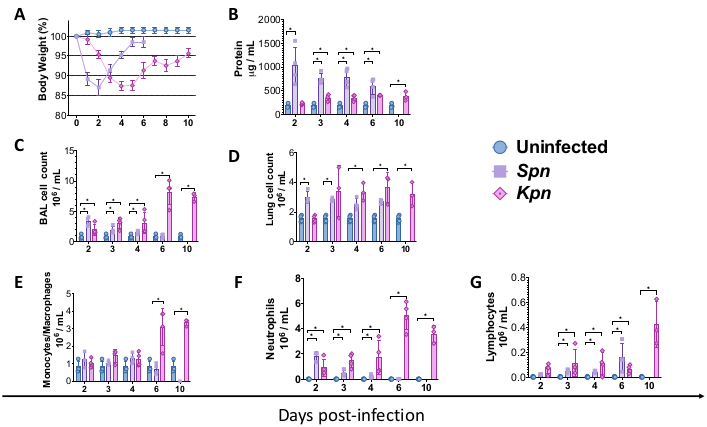
Figure 2: Acute lung injury (ALI) profiles after pneumonia models. (A) Body weight over time relative to baseline, control versus Kpn and Spn (n = 4, per group). (B) BAL total protein quantification by BCA assay (n = 4, per group). (C) BAL total cell counts in control (n = 3), Kpn (n = 4), and Spn (n = 3). (D) Lung total cell counts in control (n = 3), Kpn (n = 4), and Spn (n = 3). (E–G) BAL cell differential by manual histology count of the BAL cytospin in control (n = 3), Kpn (n = 4), and Spn (n = 3). Statistical tests were done by individual t-test, comparing the uninfected control versus the infected groups. *p < 0.05. Data is displayed using standard error (SE) for each plot. The y axis is days post-infection for all panels. Abbreviations: BAL = bronchoalveolar lavage; Spn = Streptococcus pneumoniae; Kpn = Klebsiella pneumoniae. Please click here to view a larger version of this figure.

Figure 3: Lung histopathologic findings during Spn infection. Hematoxylin and eosin (H&E) staining of histological sections of a representative BAL and lung section after the intratracheal infection of Spn and on days 2, 4, and 6. BAL magnification = 100x; lung magnification = 10x. Abbreviations: BAL = bronchoalveolar lavage; Spn = Streptococcus pneumoniae. Please click here to view a larger version of this figure.

Figure 4: Lung histopathologic findings during Kpn infection. Hematoxylin and eosin (H&E) staining of histological sections of a representative BAL and lung section after the intratracheal infection of Kpn on days 2, 4, 6, and 10. Images show a high power magnification (scale bar = 50 µm). Abbreviations: BAL = bronchoalveolar lavage; Kpn = Klebsiella pneumoniae. Please click here to view a larger version of this figure.

Figure 5: Immune cell landscape by multicolor flow cytometry after 6 days of Spn infection. (A) T-distributed stochastic neighbor embedding (t-SNE) was used to visualize the immune cell populations (CD45+) of the lung comparing the uninfected control versus infected group. (B) Summary of the immune cell frequencies out of the total CD45+ cells in the lung. (C) Total cell count of each individual population. Statistical comparisons were done by t-tests between the control and infected. *p < 0.05, **p < 0.01, ***p < 0,001. Data is displayed using standard error (SE) for each plot. Abbreviations: BAL = bronchoalveolar lavage; Spn = Streptococcus pneumoniae. Please click here to view a larger version of this figure.

Figure 6: Gating strategies for identifying the immune cell subpopulation in the lungs at baseline and after pneumonia. BAL and Lung cell suspension underwent staining for multicolor flow cytometry. Debris was gated out first using SSC-A and FSC-A, and the single cells were gated by two strategies (SSC-W vs. SSC-H and FSC-W vs. FSC-H). Live cells were identified by using a live/dead discriminator versus SSC-A. Subsequent cell populations were identified by previously identified markers 15. Abbreviation: BAL = bronchoalveolar lavage. Please click here to view a larger version of this figure.
