

Summary
Qui mostriamo i protocolli per l'esecuzione di singola molecola microscopia a fluorescenza a vivere cellule batteriche per permettere funzionali complessi molecolari per essere rilevati, monitorati e quantificati.
Abstract
Visione completa dei meccanismi delle cellule viventi può essere raggiunto solo attraverso lo studio dei processi chiave che suscitare e dirigere gli eventi a livello cellulare. Fino ad oggi la complessità dei sistemi biologici taglio ha causato preciso singola molecola sperimentazione di essere troppo esigente, concentrandosi invece su studi di singoli sistemi con massa relativamente greggio insieme alla media delle misurazioni. Tuttavia, molti processi importanti si verificano nella cellula vivente a livello di solo uno o poche molecole; misure ensemble generalmente mascherare la natura stocastica ed eterogenea di questi eventi. Qui, utilizzando avanzate di microscopia ottica e strumenti di analisi di analisi delle immagini ci mostrano come monitorare le proteine all'interno della cellula batterica vivente solo con una precisione di singole molecole e come possiamo osservare la dinamica all'interno di complessi molecolari del funzionamento macchine biologiche. Le tecniche sono direttamente rilevanti fisiologicamente. Si tratta di mini-perturbativa e non-invasivo per il campione biologico oggetto di studio e sono completamente in sintonia per le indagini in materia viva, caratteristiche non facilmente disponibili ad altri approcci singola molecola di biofisica. Inoltre, i campioni biologici studiati tutti i prodotti proteina fluorescente-tag a livelli che sono quasi identici ai ceppi cellulari non modificato ("codifica genomica"), in contrasto con l'approccio più comune ma meno ideale per la produzione di proteine molto più di quanto avverrebbe naturalmente ('plasmide di espressione'). Così, i campioni biologici reali, che saranno oggetto di indagine sono molto più vicini al organismi naturali, e quindi le osservazioni più rilevanti per le reali processi fisiologici.
Protocol
- Per iniziare questa procedura, 50 ml di scorte congelate della proteina fluorescente che esprimono Escherichia coli cellule batteriche vengono prima scongelate e coltivate aerobicamente con agitazione in 5 ml di LB mezzi di crescita durante la notte a 37 ° C. Al mattino, 50 ml di questa cultura satura viene estratto e sub-coltura in mezzi di glucosio minima M63 cultura, incubando a 30 gradi C per 4 a 6 ore. Qui mostriamo che utilizza due differenti ceppi cellulari, uno dei quali esprime un elettrone-trasporto citocromo fuso con GFP, l'altra che esprime una proteina coinvolta nel motore flagellare batterico fuso a GFP.
- Le cellule possono sia raccolto direttamente dal crescente sub-cultura, se loro visione come campioni immobilizzati, o possono essere tosato per troncare flagelli batterici se la visualizzazione in "legato" le condizioni.
- Tosatura prevede il posizionamento di solito 1-5ml di sub-cultura in un dispositivo costituito da due siringhe sterili uniti da tubi sterili. La tosatura viene effettuata alternando spingendo in ogni pompa a siringa per spingere la cultura attraverso il tubo stretto. Questo viene fatto 50-100 volte, a seconda della entità del taglio richiesto. La cultura viene poi centrifugato per agglomerare le cellule, che è risospeso in terreno minimo da rimuovere frammenti flagelli.
- Abbiamo poi preparare pulito BK7 coprioggetto di vetro immergendo in una soluzione satura di KOH in etanolo per 20 minuti, poi sciacquare accuratamente in acqua deionizzata e etanolo e lasciare asciugare all'aria per almeno 1 h.
- Costruiamo un semplice flusso di celle per ospitare le cellule al microscopio. Questa consiste nel definire le linee di grasso paraffina su un vetrino da microscopio BK7 vetro e quindi la creazione di un tunnel-panino mettendo uno dei coverlips pulita in cima, e premendo delicatamente con un paio di pinze, dando un volume di cella di flusso di 5-10 microlitri .
- Per osservare le cellule immobilizzate riempiamo il flusso delle cellule da iniezione con una w / v soluzione 0,1% di poli-L-lisina, e permettono di incubare a temperatura ambiente per almeno 1 minuto. Abbiamo poi scovare con 100 microlitri mezzi minimi, iniettando i media da un capo all'altro del flusso delle cellule e che la migrazione con carta velina dall'altro.
- Abbiamo poi lo stoppino attraverso 20 ml di una diluizione 1:500 di 200 nm di diametro in lattice microsfere (Polysciences) in terreno minimo per contrassegnare la superficie coprioggetto. Il flusso delle cellule è invertito in modo che il coprioggetto sia rivolto verso il basso, posto su una piattaforma chiara della superficie in una camera umida semplice, e incubate a temperatura ambiente per 5 min. Perline legato sono poi lavate via dalle traspirazione a 100 microlitri mezzi minimi.
- Se vogliamo osservare le cellule legato omettiamo la poli-L-lisina passo e invece riempire la cella a flusso con 5 mg / ml di anticorpi anti-flagellina. Il flusso delle cellule è posto nella camera umida per 10 minuti e viene poi eliminata attraverso di traspirazione.
- 20 l di la coltura cellulare viene poi malvagio attraverso il flusso delle cellule, sia utilizzando il campione tranciati se osservando le cellule legato, incatenato o il campione unsheared nella visione cellule immobilizzate.
- Il flusso delle cellule è invertito e collocato nella camera umida per 20 minuti. Le cellule non legati vengono lavati da traspirazione a 100 microlitri mezzi minimi.
- Una goccia di olio di immersione si trova nel centro della superficie superiore del vetrino e il flusso delle cellule viene poi messo gentilmente a supporto del campione del costume microscopio a fluorescenza costruito, il contatto ottico con le lenti ad alta apertura numerica dell'obiettivo.
- L'elettrone-moltiplicando microscopio fotocamera è accesa e la fotocamera impostata per essere raffreddato a -70 ° C, il software è impostato per acquisire immagini ad un frame rate tipico di 25 Hz in frame-modalità di trasferimento, inizialmente disenabling il controllo di guadagno sul fotocamera.
- L'illuminazione campo chiaro è acceso e l'immagine viene messa a fuoco, la selezione di un cellulare adeguata o di un gruppo di cellule da acquisire sulla base del loro essere bloccato con le loro asse parallelo alla superficie lungo coprioggetto. L'attenzione è finemente regolata per garantire che i 200 sfere di lattice nm attaccato alla superficie coprioggetto sono solo a fuoco.
- Una sequenza di immagini viene acquisita in campo chiaro per registrare il contorno del corpo cellulare. L'illuminazione campo chiaro è spento e il guadagno della telecamera è abilitato al massimo.
- Per un totale di acquisizione standard utilizzando la fluorescenza di riflessione interna (o TIRF) un fascio laser sulla lunghezza d'onda appropriata (in questo caso l'eccitazione proteina fluorescente verde, si usa un laser a 473 nm) è pre-impostato in modo da essere concentrati nel piano focale posteriore della la lente dell'obiettivo, ma spostato lateralmente rispetto all'asse ottico per generare un campo evanescente per l'eccitazione di fluorescenza nel campione di cellule.
- L'acquisizione della fotocamera viene avviato e l'otturatore aperto laser per eccitare le proteine fluorescenti all'interno del batterio. I parametri per l'intensità del laser e la velocità di acquisizione devono essere ottimizzati per il particolare sistema biologico sotto inchiesta da sperimentare con valori diversi, ma una gamma tipica rignificative a studiare complessi proteici mobile nella membrana cellulare sono 10-10 mW di potenza del laser su una superficie di eccitazione circolare del diametro di circa 30 micron, con un tempo di esposizione per ogni fotogramma di 50-40 ms. I campioni sono illuminate fino fotodecolorate, di solito per circa 10 s.
- Questo protocollo può essere utilizzato sia per le cellule legato in cui il corpo della cellula ruota su un punto di attacco tra il coprioggetti e un mozzicone di flagelli, e per le cellule immobilizzate che sono rigidamente fissate alla superficie coprioggetto.
- Alcuni ceppi di cellule che hanno un elevato numero di complessi copia beneficiare di una prima diffrazione limitata candeggina laser focalizzato su un polo della cellula prima TIRF imaging. Questa candeggina è equivalente a quella utilizzata per il recupero di fluorescenza dopo photobleaching (o FRAP). Utilizzando il nostro microscopio personalizzati parte della luce laser di eccitazione può essere alimentata via in un secondo percorso indipendente utilizzato per FRAP-tipo sbiancamento. Tipicamente 1-10mW potenza del laser è utilizzato, con un tempo di candeggina tipico nella gamma 10-300 ms. Il risultato è in contrasto con immagini molto più alta nella zona sbiancata della cellula permettendo complessi individuali che successivamente diffusa in quella zona per essere più facili da visualizzare.
- Per visualizzare i complessi nel citoplasma cellulare che diffondono molto più velocemente rispetto a quelli della membrana cellulare è impiegato un modo diverso di illuminazione chiamato "slimfield". Ecco un laser focalizzato si espande lateralmente fino a comprendere solo una singola cella. Questo produce un campo molto intenso che permette esposizioni molto più veloce di solito un millisecondo da adottare.
- A seguito di acquisizione dati, le immagini sono inseriti in software personalizzato scritto (codificato in LabVIEW 8.5). Questo rileva automaticamente la posizione delle macchie fluorescenti nelle cellule con una precisione di pochi nanometri tipicamente ed estratti delle loro dimensioni e luminosità. Il brightnessof la traccia photobleaching rispetto al tempo di un complesso molecolare tracciato viene quindi utilizzato per stimare la stechiometria, cioè quante singole proteine fluorescenti formano un unico complesso molecolare.
Rappresentante dei risultati:
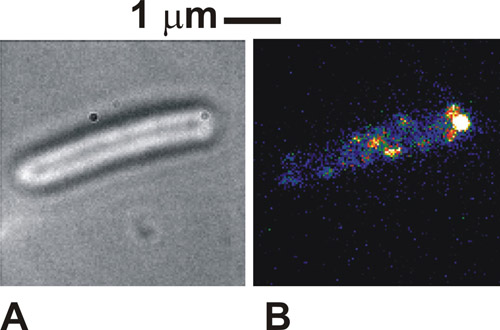
Quando il protocollo è fatto correttamente le immagini delle cellule visto in campo chiaro è molto distinto, con i perimetri dei corpi cella buia contro un corpo cellulare bianco / grigio (Figura 1a). In fluorescenza utilizzando cellule immobilizzate, possiamo vedere l'intensità distinte macchie, di solito 250-300 nm di larghezza (figura 1b). Sane, le cellule si vedrà legato a ruotare attorno al punto di attacco legare in campo chiaro le immagini. Sotto eccitazione di fluorescenza alcuni complessi molecolari nel nostro caso potrebbe anche essere visto al punto di attacco, che indica una localizzazione della proteina taggati con il motore flagellare. Queste macchie sono individuali complessi molecolari e il numero di viste dipende dalla modalità di illuminazione utilizzati e quanti dei complessi sono in realtà presenti nella cellula in un dato momento. La mobilità dei punti dipende dal sistema biologico specifico oggetto di studio. Se la densità dei punti è inizialmente molto alta, come è il caso con i citocromi marcato usato qui, poi eseguire una candeggina FRAP iniziale può migliorare il contrasto delle immagini.

Figura 1. (A) Brightfield e (B) immagine TIRF (falsi colori) per un immobilizzato cellule di Escherichia coli che esprimono una proteina fusa alla proteina fluorescente verde (GFP) che è noto per essere coinvolto nei motori flagellare di batteri. Si prega di cliccare qui per vedere una versione più grande della figura 1.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Bisogna fare attenzione a non "oltre al taglio" delle cellule per guardare i batteri legati, poiché questo può compromettere la funzionalità del motore flagellare. E 'importante utilizzare le cellule per molto più tempo di un'ora una volta sul vetrino da microscopio in quanto possono diventare ossigeno scarica. Notevole ottimizzazione può essere richiesto di trovare migliori condizioni di imaging microscopio soddisfatti al vostro specifico sistema biologico sotto inchiesta. Potrebbe essere saggio tentare l'imaging utilizzando purificato GFP solo per accertare la corretta eccitazione laser intensità richiesta per il sistema microscopio particolare.
Subscription Required. Please recommend JoVE to your librarian.
Acknowledgments
Noi riconosciamo le donazioni tipo di ceppi batterici dai gruppi del Prof. Judith Armitage (Università di Oxford, UK) e il Prof. Corrado Mullineaux (Queen Mary University of London, UK). IMD è finanziato congiuntamente dal Dipartimento di Biochimica (Oxford University) e OCISB; AR è finanziato da un Engineering and Physical Sciences Research Council (EPSRC) borsa di studio DTC; ND è finanziata dal Biotechnology and Biological Sciences Research Council (BBSRC); MCL è finanziato da una borsa di studio della Royal Society University Research.
References
- Leake, M. C., Chandler, J. H., Wadhams, G. H., Bai, F., Berry, R. M., Armitage, J. P. Stoichiometry and turnover in single, functioning membrane protein complexes. Nature. 443, 355-358 (2006).
- Leake, M. C., Greene, N. P., Godun, R. M., Granjon, T., Buchanan, G., Chen, S., Berry, R. M., Palmer, T., Berks, B. C. Variable stoichiometry of the TatA component of the twin-arginine protein transport system observed by in vivo single-molecule imaging. Proc Natl Acad Sci U S A. 105, 15376-15381 (2008).
- Leake, M. C., Wilson, D., Gautel, M., Simmons, R. M., M, R. The elasticity of single titin molecules using a two-bead optical tweezers assay. Biophys. J. 87, 1112-1135 (2004).
- Leake, M. C., Wilson, D., Bullard, B., Simmons, R. M. The elasticity of single kettin molecules using a two-bead laser-tweezers assay. FEBS Lett. , 535-555 (2003).
- Lenn, T., Leake, M. C., Mullineaux, C. W. In vivo clustering and dynamics of cytochrome bd complexes in the Escherichia coli plasma membrane. Mol. Microbiol. 70, 1397-1407 (2008).
- Lenn, T., Leake, M. C., Mullineaux, C. W. Are Escherichia coli OXPHOS complexes concentrated in specialised zones within the plasma membrane. Biochem. Soc. Trans. 36, 1032-1036 (2008).
- Lo, C. J., Leake, M. C., Pilizota, T., Berry, R. M. Single-cell measurements of Membrane Potential, Sodium-Motive Force and Flagellar Motor Speed in Escherichia coli. Biophys. 93, 294-302 (2007).
- Lo, C. J., Leake, M. C., Berry, R. M. Fluorescence measurement of intracellular sodium concentration in single Escherichia coli cells. Biophys. J. 90, 357-3565 (2006).

{kind=link}