

Summary
Белки трансдукции позволяет прямой доставки биологически активных белков в клетки. В отличие от традиционных методов, таких как трансфекции ДНК или вирусной трансдукции этой неинвазивной парадигма позволяет высокоэффективных сотовых манипуляции в титруемой образом в обход сотового токсичность и риск онкогенных преобразование постоянного генетической модификации.
Abstract
Техника белков трансдукции позволяет прямой доставки биологически активных веществ в клетках млекопитающих [см. обзор 1,2]. Для этого можно использовать translocating способность так называемого ячейки проникающих пептидов (ППС), также обозначается как белка трансдукции доменов (PTDs). TAT-CPP происходит от иммунодефицита человека вирусом типа 1 (ВИЧ-1) Tat (транс-активатор транскрипции) белок широко используется. Положительно заряженные ТАТ способствует клеточной проницаемости тем самым преодолевая барьеры клеточной мембраны путем эндоцитоза и / или прямое проникновение мембраны 2. В сочетании с сигнал ядерной локализации (NLS) слитые белки способны проникать ядра выставке функциональность. Наши видео-презентацию демонстрирует, как иллюстрация для инженерных клеточных белков проницаемой, строительства, производства и применения клеточных проницаемой версию ДНК-модифицирующие фермент Cre.
Cre является конкретным участкам рекомбиназы, который может распознавать и рекомбинировать 34 база сайтов пара loxP в клетках млекопитающих в пробирке и в естественных условиях. Поэтому Cre / loxP система широко используется для условного вызывать мутации в геноме живых клеток 3,4. Доставки активных Cre рекомбиназы к клеткам, однако, представляет собой ограничение.
Мы описываем системы pSESAME вектор, который позволяет прямое включение гена интересов, и предоставляет платформу для быстрого клона разных доменах и тегов, используемых в вектор в удобной и стандартизованным образом. Реорганизация различных тегов, как было показано изменение биохимических свойств белков слияния предоставление возможности для достижения более высокой урожайности и лучшей растворимости. Мы демонстрируем, как можно выразить и очистить рекомбинантных клеточных белков в проникающий и от кишечной палочки. Функциональность рекомбинантный белок Cre, наконец, утверждены в культуре клеток, оценивая его внутриклеточной активности рекомбиназы.
Protocol
Строительство вектор экспрессии и выражения:
PSESAME-Cre вектор экспрессии была построена вставка Cre-кодирования фрагмента в pSESAME через AvrII и NheI сайты рестрикции с использованием стандартных методов клонирования. pSESAME кодирует белок слияния, состоящий из гистидина-теги, ТАТ-область, NLS последовательности и Cre, сокращенно HTNCre. Для выражения HTNCre pSESAME-Cre было преобразовано в TUNER (DE3) pLacI и используется для подготовки глицерина акций.
- В течение ночи культуру засевают использованием пипетки наконечник покрытием с преобразованным бактерий из глицерина акций. В течение ночи культуры состояла из LB среде с 0,5% глюкозы [об. /] и карбенициллин в конечной концентрации 50 мкг / мл и было позволено расти при температуре 37 ° С в течение 16 часов.
- На следующий день густо выросли на ночь культуры используют для инокуляции выражение культуры в соотношении 1 к 40 и был помещен в инкубатор при 37 ° C. Выражение культуры состояла из ТБ среде с 0,5% глюкозы [об. /] и ампициллин в конечной концентрации 100 мкг / мл.
- В OD 595 из 1,5 выражение культуры вызывали с 0,5 мМ IPTG в течение 1 ч.
- Впоследствии бактерий были собраны путем центрифугирования при 5000 оборотах в минуту в течение 10 минут в SLA3000 ротора.
- Бактерии гранулы хранились при температуре минус 20 ° С до очистки.
Очистка ячейки проницаемой белка:
- Замороженные гранулы бактерии ресуспендировали в 10 мл лизирующего буфера на литр культуры колбу в течение 15 минут при комнатной температуре.
- Затем суспензию инкубировали с 1 мг / мл лизоцима еще на 15 минут при перемешивании при комнатной температуре.
- 25 Ед / мл benzonase был добавлен позже и инкубировали при перемешивании в течение 15 минут при комнатной температуре.
- После озвучивания на льду в течение 1,5 мин с 0,5 импульсов с при 45% мощности, 1 мл холодного буфера соль винной (БСЭ) в мл суспензии осторожно добавляют при перемешивании и выдерживают в течение 5 мин на льду. SDS-PAGE образец лизат фракции (L) принято не было.
- Очищенные лизат получали центрифугированием при 4 ° С в течение 30 мин при 30000 g. SDS-PAGE образцов растворимого (S) и нерастворимых фракций (I) были приняты.
- Супернатант переносили в 50 мл свежего труб сокола и был затем осторожно перемешивают в течение 1 ч при 4 ° С с 2 мл 50% Ni-NTA шлама на литр первичной культуры выражения.
- Суспензию упакованы в самотеком EconoPac колонке (SDS-PAGE образец проточного фракции (FT) была сделана) и дважды промывают 5 постельное объемов моющий буфер. SDS-PAGE образцах как мытье фракций (W1 и W2) были собраны.
- HTNCre-содержащие фракции элюировали 3 постельное объемами буфера элюирования и образца элюата фракции (Е) для SDS-PAGE анализа был взят.
- Имидазола удаляли диализа элюирования фракция против высоких буфера соли в два раза.
- Раствора белка еще более концентрируется диализа против буфера глицерина в два раза. Во всех диализных шаги отношение буфер для образца, по крайней мере 50. Эта процедура привела к решению глицерина семенного материала, HTNCre в обычной концентрации от 200 до 450 мкм, то есть 1 литр выражение культуры приведет к ~ 12 мг белка. Пример глицерина акций (GS) для SDS-PAGE анализа была собрана. HTNCre маточный раствор можно хранить при температуре минус 20 ° С.

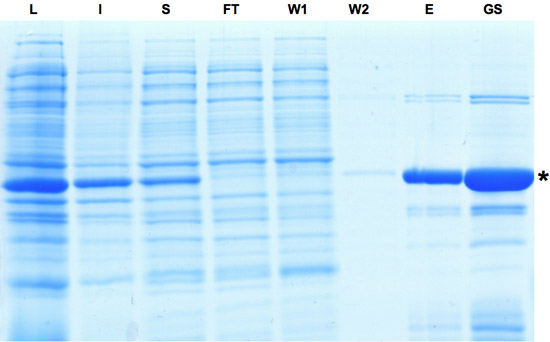
Рисунок 1: SDS-PAGE анализа проб, взятых при очистке Cre рекомбиназы. Индукция выражение Cre указывается доминирующей группой в лизат фракции. Хотя часть белка нерастворимые белки Cre может быть обогащен как видно из элюата и фракции глицерина акций. L: лизат, я: Не растворяется, S: Супернатант, FT: проточные, Вт: стиральная, E: элюата, Г. С.: Gylcerol бирже. Пожалуйста, нажмите здесь , чтобы видеть большую версию рисунке 1.
Белки трансдукции в мышиных эмбриональных стволовых (ЭС) клеток:
- ES клеток, несущих условной β-галактозидазы репортер 5 построить высевают как отдельные клетки, используя TrypLE ™ Express для диссоциации прилипшие клетки. После 4-х до 6 часов клетки было повторное подключение и среду удаляли.
- Впоследствии ЭС клетки инкубировали с HTNCre среде, содержащей в течение 16 часов.
- Соответствующее количество белка HTNCre (соответствует 10 мкМ) из глицерина акции разбавляли в среду ES, а затем стерильной фильтрации (0,22 мкм).
- После среду белков трансдукции был изменен обратно в нормальный средний рост.
- После двух дней клетки промывали PBS и фиксировали 4% Параформальдегид (PFA) в течение 10 минут.
- Два дополнительныхных этапов промывки с PBS были казнены до X-Gal окрашивание проводилось.
- Фиксированные клетки были покрыты слоем X-Gal красящим раствором 6 и инкубировали в течение ночи при температуре 37 ° C.
Представитель Результаты:
На следующий день X-Gal окрашивание раствора стремилась и клетки были покрыты слоем PBS для микроскопии анализа. От 80 до 100% от рекомбинированное клетки можно было наблюдать в мышиных клетках ES судить по β-галактозидазы.
Discussion
Во время процесса очистки слитый белок Cre важно не пропустить добавлением льда буфера холодной TBS перед центрифугированием. В противном случае Cre рекомбиназы имеет тенденцию осаждаться внутри глицерина буфера.
Если элюата фракция появляется, чтобы стать мутной из-за высокой концентрации гибридный белок дополнительного буфера элюирования следует добавить, пока раствор не прояснилось снова.
Применение 10 мкмоль белка слияния Cre обычно приводит к рекомбинации эффективность от 80 до 100%. Эмбриональной телячьей сыворотки (FCS) является одной из основных компонент среднего ячейки ES сильно подавляет белок трансдукции. Поэтому высокая концентрация Cre рекомбиназы должны были быть использованы. При работе в бессывороточной условиях меньше белка (0,5 - 2 мкм) может быть использован для достижения аналогичных эффективность рекомбинации.
С системой pSESAME вектор в стороны, можно применить технику белков трансдукции к другим белкам, включая транскрипционные факторы, такие как Oct4 и Sox2 7 и Scl/Tal1 8.
Acknowledgments
Мы благодарим Оливера Brüstle и всех членов инженерно стволовых клеток группы, Боннский университет, за поддержку и ценные обсуждения. Мы благодарим Сабина Шенк для подготовки SDS-PAGE и прочной поддержкой на протяжении всего проекта. Николь Руси и Анна Magerhans при условии, отличную техническую поддержку. Кроме того, мы хотели бы поблагодарить Андреаса и Шейла Бэр Мертенса для производства фильма. Эта работа была поддержана грантами фонда Фольксваген (Az I/77864) и германского министерства образования и научных исследований (BMBF, 01 GN 0813).
Materials
| Name | Company | Catalog Number | Comments |
| TUNER (DE3) pLacI | Novagen, EMD Millipore | 70625 | |
| Glycerol | Carl Roth GmbH | 3783.2 | |
| Na2HPO4 | Carl Roth Gmbh | T876.1 | |
| Trizma Base | Sigma-Aldrich | T1503 | |
| HCl | Carl Roth Gmbh | 4625.1 | |
| Imidazol | Carl Roth Gmbh | X998.4 | |
| NaCl | Carl Roth Gmbh | 9265.2 | |
| Yeast Extract | Carl Roth Gmbh | 2363.4 | |
| Trypton/Pepton | Carl Roth Gmbh | 8952.4 | |
| K2HPO4 | Carl Roth Gmbh | P749.2 | |
| KH2PO4 | Carl Roth Gmbh | 3904.1 | |
| Ampicillin | Sigma-Aldrich | A9518 | |
| Carbenicillin | Sigma-Aldrich | 6344.2 | |
| HEPES | Sigma-Aldrich | H3375 | |
| Lysozyme | Sigma-Aldrich | 62971 | |
| Benzonase | Novagen, EMD Millipore | ||
| L-Tartaric acid, disodium salt | Sigma-Aldrich | ||
| 50% Ni-NTA slurry | Invitrogen | R901-15 | |
| EconoPac columns | Bio-Rad | 732-1010 | |
| Sterile filter 0,22μm | Whatman, GE Healthcare | ||
| Paraformaldehyde (PFA) | Sigma-Aldrich | ||
| LB medium | Yeast extract, Trypton/Pepton, NaCl | ||
| TB medium | Yeast extract, Trypton/Pepton, Glycerol, K2HPO4, KH2PO4 | ||
| Lysis Buffer | 50 mM Na2HPO4, 5 mM Tris, pH 7.8 | ||
| Tartaric Salt Buffer (TSB) | PTB containing 2 M L-Tartaric acid, disodium salt, and 20 mM Imidazol | ||
| Washing Buffer | PTB, 500 mM NaCl, 15 mM Imidazol | ||
| Elution Buffer | PTB, 500 mM NaCl, 250 mM Imidazol | ||
| High Salt Buffer | 600 mM NaCl, 20 mM HEPES, pH 7.4 | ||
| Gylcerol Buffer | 50% glycerol, 500 mM NaCl, 20 mM HEPES, pH 7.4 | ||
| TrypLE™ Express | Invitrogen | ||
| ESGRO (LIF) | EMD Millipore | ||
| NEAA | GIBCO, by Life Technologies | 11140035 | |
| L-Glutamin | GIBCO, by Life Technologies | 25030024 | |
| β-Mercapt–thanol | GIBCO, by Life Technologies | 31350010 | |
| DMEM | GIBCO, by Life Technologies | 11960044 | |
| PBS | GIBCO, by Life Technologies | ||
| Fetal Calf Serum (FCS) | PAA Laboratories | ||
| X-Gal staining solution: | 4 mM K3(FeIII(CN)6), 4 mM K4(FeII(CN)6), 2mM MgCl2 0.4 mg/mL X-Gal solved in PBS |
||
| K3(FeIII(CN)6) | Sigma-Aldrich | P-3367 | |
| K4 (FeII(CN)6) | Sigma-Aldrich | P-9387 | |
| MgCl2 | Sigma-Aldrich | M8266 | |
| X-Gal | Sigma-Aldrich | B4252 |
References
- Gump, J. M., Dowdy, S. F. TAT transduction: the molecular mechanism and therapeutic prospects. Trends Mol Med. 13 (10), 443-448 (2007).
- Edenhofer, F. Protein transduction revisited: novel insights into the mechanism underlying intracellular delivery of proteins. Curr Pharm Des. 14 (34), 3628-3636 (2008).
- Branda, C. S., Dymecki, S. M. Talking about a revolution: The impact of site-specific recombinases on genetic analyses in mice. Dev Cell. 6 (1), 7-28 (2004).
- Nolden, L. Site-specific recombination in human embryonic stem cells induced by cell-permeant Cre recombinase. Nat Methods. 3 (6), 461-467 (2006).
- Zhang, Y. Inducible site-directed recombination in mouse embryonic stem cells. Nucleic Acids Res. 24 (4), 543-548 (1996).
- Peitz, M. Enhanced purification of cell-permeant Cre and germline transmission after transduction into mouse embryonic stem cells. Genesis. 45 (8), 508-517 (2007).
- Bosnali, M., Edenhofer, F. Generation of transducible versions of transcription factors Oct4 and Sox2. Biol Chem. 389 (7), 851-861 (2008).
- Landry, J. R. Runx genes are direct targets of Scl/Tal1 in the yolk sac and fetal liver. Blood. 111 (6), 3005-3014 (2008).

{kind=link}