

Summary
Eiwit transductie maakt de directe levering van biologisch actieve eiwitten in cellen. In tegenstelling tot conventionele methoden, zoals DNA-transfectie of virale transductie deze niet-invasieve paradigma maakt zeer efficiënte cellulaire manipulatie in een titreerbare manier te omzeilen cellulaire toxiciteit en het risico van oncogene transformatie door permanente genetische modificatie.
Abstract
Het eiwit transductie-techniek maakt de directe levering van biologisch actieve materiaal in cellen van zoogdieren [voor review zie 1,2]. Voor deze kan men gebruik maken van de transloceren vermogen van de zogenaamde cel-penetrerende peptiden (CPP) te maken, ook wel aangeduid als eiwit transductie domeinen (PTDs). De TAT-CPP afgeleid van het humaan immunodeficiëntie virus type 1 (HIV-1) Tat (trans-activator van transcriptie) eiwit is op grote schaal gebruikt. De positief geladen TAT bevordert celdoorlaatbaarheid waardoor het overwinnen van de barrières van de cellulaire membraan door endocytose en / of directe membraan penetratie 2. In combinatie met een nucleair lokalisatie signaal (NLS) fusie-eiwitten zijn in staat om de kern exposeren functionaliteit in te voeren. Onze video presentatie laat zien, als een voorbeeldgedrag voor de engineering van cel-permeabele eiwitten, de bouw, productie en toepassing van een cel-permeabele versie van de DNA-modificerende enzym Cre.
Cre is een site-specific recombinase die in staat is te herkennen en 34 basenparen loxP plaatsen recombineren in zoogdiercellen in vitro en in vivo. Daarom is het Cre / loxP systeem wordt veel gebruikt om voorwaardelijk mutaties te induceren in het genoom van levende cellen 3,4. De levering van de actieve Cre recombinase aan cellen, echter, is een beperking.
Beschrijven we de pSESAME vector-systeem, dat een directe insertie van het gen-of-interest mogelijk maakt en biedt een platform om snel kloon verschillende domeinen en tags gebruikt binnen de vector in een handige en gestandaardiseerde wijze. Herschikken van de verschillende labels is aangetoond dat de biochemische eigenschappen van de fusie-eiwitten die een mogelijkheid om hogere opbrengst en een betere oplosbaarheid te bereiken te wijzigen. Laten we zien hoe uit te drukken en recombinante cel-permeant eiwitten te zuiveren in en van E. coli. De functionaliteit van het recombinante eiwit Cre is uiteindelijk gevalideerd in cel cultuur door de beoordeling van de intracellulaire recombinase activiteit.
Protocol
Constructie van expressievector en expressie:
De pSESAME-Cre expressie vector werd geconstrueerd door het invoegen van een Cre-codering fragment in pSESAME via AvrII en Nhel restrictie sites met behulp van standaard-klonen methoden. pSESAME codeert voor een fusie-eiwit dat bestaat uit een histidine-tag, TAT-domein, NLS volgorde en Cre, afgekort HTNCre. Voor expressie van HTNCre de pSESAME-Cre werd getransformeerd in TUNER (DE3) pLacI en gebruikt om een glycerol voorraad te bereiden.
- Een over-night cultuur werd geënt met behulp van een pipet tip bekleed met veranderde bacteriën uit de glycerol voorraad. De over-avond cultuur bestond uit LB media aangevuld met 0,5% glucose [v / v] en carbenicilline bij een uiteindelijke concentratie van 50 ug / ml en mocht om te groeien bij 37 ° C gedurende 16 uur.
- De volgende dag de dichtbevolkte gegroeid nacht cultuur werd gebruikt om de uitdrukking cultuur inoculeren in een verhouding van 1 tot 40 en werd in een incubator bij 37 ° C. Expressie cultuur bestond uit TB media aangevuld met 0,5% glucose [v / v] en ampicilline bij een uiteindelijke concentratie van 100 ug / ml.
- Bij een OD 595 van 1,5 de uitdrukking cultuur werd geïnduceerd met 0,5 mM IPTG gedurende 1 uur
- Vervolgens bacteriën werden verzameld door centrifugeren bij 5000 rpm gedurende 10 minuten in een SLA3000 rotor.
- Bacteriën pellets werden opgeslagen bij min 20 ° C tot zuivering.
Zuivering van cel-permeabele eiwit:
- Bevroren bacteriën pellets werden geresuspendeerd in 10 ml lysis buffer per liter fles cultuur gedurende 15 minuten bij kamertemperatuur.
- Suspensie werd vervolgens geïncubeerd met 1 mg / ml lysozym voor nog eens 15 minuten tijdens het mengen bij kamertemperatuur.
- 25 U / mL benzonase is later toegevoegd en geïncubeerd tijdens het mengen gedurende 15 minuten bij kamertemperatuur.
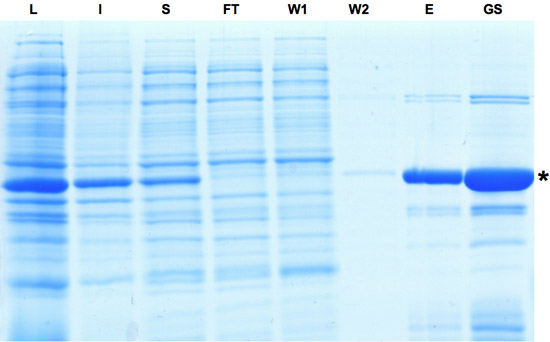
- Na sonificatie op ijs 1,5 min met 0,5 s pulsen bij 45% van het vermogen, 1 ml koud wijnsteenzuur zout buffer (TSB) per ml suspensie werd zorgvuldig toegevoegd tijdens het mengen en geïncubeerd gedurende 5 minuten op ijs. SDS-PAGE steekproef van lysaat fractie (L) werd genomen.
- Gewist lysaat werd verkregen door centrifugeren bij 4 ° C gedurende 30 min bij 30000 g. SDS-PAGE monsters van oplosbare (S) en de onoplosbare fracties (I) werden genomen.
- Het supernatant werd overgebracht in de frisse 50 ml Falcon buizen en werd vervolgens voorzichtig gemengd gedurende 1 uur bij 4 ° C met 2 ml van 50% Ni-NTA slurry per liter van de initiële expressie cultuur.
- De suspensie werd verpakt in een zwaartekracht EconoPac kolom (SDS-PAGE sample van flow-through fractie (FT) werd genomen) en tweemaal gewassen met 5 bed-volumes van wasbuffer. SDS-PAGE monsters van zowel het wassen van fracties (W1 en W2) werden verzameld.
- HTNCre-bevattende fracties werden geëlueerd met drie bed-volumes van elutie buffer en steekproef van eluaat fractie (E) voor SDS-PAGE-analyse werd genomen.
- Imidazool werd verwijderd door dialyseren elutie fractie tegen hoge zout-buffer tweemaal.
- Het eiwit-oplossing werd verder geconcentreerd door dialyseren tegen glycerol buffer tweemaal. In alle dialyse stappen de verhouding van de buffer om te proeven was op zijn minst 50. Deze procedure resulteerde in een glycerol voorraad oplossing die HTNCre op een normale concentratie tussen 200 en 450 uM, dwz 1 liter van meningsuiting cultuur resulteren in ~ 12 mg eiwit. Voorbeeld van glycerol voorraad (GS) voor SDS-PAGE-analyse werd verzameld. HTNCre voorraad oplossing kan worden bewaard bij min 20 ° C.

Figuur 1: SDS-PAGE-analyse van de monsters verzameld tijdens het zuiveringsproces van het Cre recombinase. Inductie van Cre expressie wordt aangegeven door dominante band in lysaat fractie. Hoewel een deel van het eiwit is onoplosbaar het Cre eiwit kan verder worden verrijkt zoals te zien in eluaat en glycerol voorraad fracties. L: Lysate, I: Niet oplosbaar, S: Supernatant, FT: Flow-through, W: Wassen, E: eluaat GS: Gylcerol Stock. Gelieve Klik hier voor een grotere versie van figuur 1 te zien.
Proteïne transductie in muriene embryonale stamcellen (ES)-cellen:
- ES-cellen die een voorwaardelijke β-galactosidase reporter 5 construct werden gezaaid als enkele cellen met behulp van TrypLE ™ Express voor dissociatie van hechtende cellen. Na 4 tot 6 uur de cellen opnieuw had aangesloten en het medium werd verwijderd.
- Vervolgens ES-cellen werden geïncubeerd met HTNCre-bevattend medium voor 16 uur.
- Een gepaste hoeveelheid HTNCre eiwit (overeenkomend met 10 uM) uit de glycerol voorraad werd verdund in ES medium en vervolgens steriel gefiltreerd (0,22 pm).
- Na het proteïne transductie medium werd veranderd terug naar de normale groei medium.
- Na twee dagen werden de cellen gewassen met PBS en gefixeerd met 4% paraformaldehyde (PFA) gedurende 10 minuten.
- Twee extrationale wasstappen met PBS werden uitgevoerd voor de X-Gal kleuring werd uitgevoerd.
- Vaste cellen werden bedekt met een laag van X-Gal kleuroplossing 6 en geïncubeerd gedurende de nacht bij 37 ° C.
Representatieve resultaten:
De volgende dag X-Gal kleuroplossing werd streefde en de cellen werden bedekt met een laag van PBS voor microscopie analyse. 80 tot 100% van de gerecombineerde cellen kon worden waargenomen binnen de muriene embryonale stamcellen beoordeeld door β-galactosidase-activiteit.
Discussion
Tijdens het zuiveringsproces van het Cre fusie-eiwit is het belangrijk niet te vergeten de toevoeging van ijskoude TBS-buffer voorafgaand aan de centrifugeren. Anders Cre recombinase de neiging neer te slaan binnen de glycerol buffer.
Als het eluaat fractie lijkt te worden om te troebel als gevolg van de hoge concentratie van fusie-eiwit extra elutiebuffer moeten worden toegevoegd totdat de oplossing is weer gewist.
De toepassing van 10 uM van Cre fusie-eiwit resulteert typisch in een recombinatie rendement van 80 tot 100%. Foetaal Calf Serum (FCS) als een belangrijk onderdeel van ES-cel medium sterk remt proteïne transductie. Daarom is een hoge concentratie van Cre-recombinase moest worden gebruikt. Bij het werken in serum-vrije omstandigheden minder eiwit (0,5 - 2 uM) kan worden gebruikt om vergelijkbare recombinatie efficiëntie te bereiken.
Met het pSESAME vector-systeem bij de hand kan men toepassen van de techniek van eiwitten transductie aan andere eiwitten, met inbegrip transcriptiefactoren, zoals Oct4 en Sox2 7 en 8 Scl/Tal1.
Acknowledgments
Wij danken Oliver Brüstle en alle leden van de Stem Cell Engineering Group, Universiteit van Bonn, voor ondersteuning en waardevolle discussies. Wij danken Sabine Schenk voor de voorbereiding van de SDS-PAGE en blijvende ondersteuning gedurende het project. Nicole Russ en Anna Magerhans voorzien uitstekende technische ondersteuning. Verder willen we Andreas Bär en Sheila Mertens bedanken voor de productie van de film. Dit werk werd ondersteund door subsidies van de Volkswagen Foundation (Az I/77864) en het Duitse ministerie van Onderwijs en Onderzoek (BMBF, 01 GN 0813).
Materials
| Name | Company | Catalog Number | Comments |
| TUNER (DE3) pLacI | Novagen, EMD Millipore | 70625 | |
| Glycerol | Carl Roth GmbH | 3783.2 | |
| Na2HPO4 | Carl Roth Gmbh | T876.1 | |
| Trizma Base | Sigma-Aldrich | T1503 | |
| HCl | Carl Roth Gmbh | 4625.1 | |
| Imidazol | Carl Roth Gmbh | X998.4 | |
| NaCl | Carl Roth Gmbh | 9265.2 | |
| Yeast Extract | Carl Roth Gmbh | 2363.4 | |
| Trypton/Pepton | Carl Roth Gmbh | 8952.4 | |
| K2HPO4 | Carl Roth Gmbh | P749.2 | |
| KH2PO4 | Carl Roth Gmbh | 3904.1 | |
| Ampicillin | Sigma-Aldrich | A9518 | |
| Carbenicillin | Sigma-Aldrich | 6344.2 | |
| HEPES | Sigma-Aldrich | H3375 | |
| Lysozyme | Sigma-Aldrich | 62971 | |
| Benzonase | Novagen, EMD Millipore | ||
| L-Tartaric acid, disodium salt | Sigma-Aldrich | ||
| 50% Ni-NTA slurry | Invitrogen | R901-15 | |
| EconoPac columns | Bio-Rad | 732-1010 | |
| Sterile filter 0,22μm | Whatman, GE Healthcare | ||
| Paraformaldehyde (PFA) | Sigma-Aldrich | ||
| LB medium | Yeast extract, Trypton/Pepton, NaCl | ||
| TB medium | Yeast extract, Trypton/Pepton, Glycerol, K2HPO4, KH2PO4 | ||
| Lysis Buffer | 50 mM Na2HPO4, 5 mM Tris, pH 7.8 | ||
| Tartaric Salt Buffer (TSB) | PTB containing 2 M L-Tartaric acid, disodium salt, and 20 mM Imidazol | ||
| Washing Buffer | PTB, 500 mM NaCl, 15 mM Imidazol | ||
| Elution Buffer | PTB, 500 mM NaCl, 250 mM Imidazol | ||
| High Salt Buffer | 600 mM NaCl, 20 mM HEPES, pH 7.4 | ||
| Gylcerol Buffer | 50% glycerol, 500 mM NaCl, 20 mM HEPES, pH 7.4 | ||
| TrypLE™ Express | Invitrogen | ||
| ESGRO (LIF) | EMD Millipore | ||
| NEAA | GIBCO, by Life Technologies | 11140035 | |
| L-Glutamin | GIBCO, by Life Technologies | 25030024 | |
| β-Mercapt–thanol | GIBCO, by Life Technologies | 31350010 | |
| DMEM | GIBCO, by Life Technologies | 11960044 | |
| PBS | GIBCO, by Life Technologies | ||
| Fetal Calf Serum (FCS) | PAA Laboratories | ||
| X-Gal staining solution: | 4 mM K3(FeIII(CN)6), 4 mM K4(FeII(CN)6), 2mM MgCl2 0.4 mg/mL X-Gal solved in PBS |
||
| K3(FeIII(CN)6) | Sigma-Aldrich | P-3367 | |
| K4 (FeII(CN)6) | Sigma-Aldrich | P-9387 | |
| MgCl2 | Sigma-Aldrich | M8266 | |
| X-Gal | Sigma-Aldrich | B4252 |
References
- Gump, J. M., Dowdy, S. F. TAT transduction: the molecular mechanism and therapeutic prospects. Trends Mol Med. 13 (10), 443-448 (2007).
- Edenhofer, F. Protein transduction revisited: novel insights into the mechanism underlying intracellular delivery of proteins. Curr Pharm Des. 14 (34), 3628-3636 (2008).
- Branda, C. S., Dymecki, S. M. Talking about a revolution: The impact of site-specific recombinases on genetic analyses in mice. Dev Cell. 6 (1), 7-28 (2004).
- Nolden, L. Site-specific recombination in human embryonic stem cells induced by cell-permeant Cre recombinase. Nat Methods. 3 (6), 461-467 (2006).
- Zhang, Y. Inducible site-directed recombination in mouse embryonic stem cells. Nucleic Acids Res. 24 (4), 543-548 (1996).
- Peitz, M. Enhanced purification of cell-permeant Cre and germline transmission after transduction into mouse embryonic stem cells. Genesis. 45 (8), 508-517 (2007).
- Bosnali, M., Edenhofer, F. Generation of transducible versions of transcription factors Oct4 and Sox2. Biol Chem. 389 (7), 851-861 (2008).
- Landry, J. R. Runx genes are direct targets of Scl/Tal1 in the yolk sac and fetal liver. Blood. 111 (6), 3005-3014 (2008).

{kind=link}