

Summary
Genetiska studier i jäst kan användas för att undersöka de molekylära och cellulära funktioner av mänskliga gener i cellens DNA metabolism. Metoder beskrivs för genetisk karaktärisering av det mänskliga
Abstract
Förstå roller humana proteiner DNA-reparation i genetiska vägar är en oerhörd utmaning för många forskare. Genetiska studier i däggdjur system har varit begränsad på grund av avsaknaden av lättillgängliga verktyg som definieras mutant genetiska cellinjer, regelsystem uttryck, och lämpliga valbara markörer. För att kringgå dessa svårigheter har modell genetiska system i lägre eukaryoter bli ett attraktivt val för att studera funktionellt bevarade proteiner DNA-reparation och vägar. Vi har utvecklat ett system för modell jäst för att studera dåligt definierade genetiska funktioner Werner syndrom helikas-nukleas (
Protocol
1. Jäststammar
Stammar med vildtyp SGS1 Top3 (WT, W303-1A, genotyp, MAT ett ade2-1 CANL-100 his3-11, 15 leu2-3, 112 TRPL-l ura3-1) [2], en sgs1 mutant (W1292-3C ; genotyp MAT ett SUP4-o:: URA3 sgs1-25 ade2-1 can1-100 his3-11, 15 leu2-3, 112 trp1-1 ura3-1 rad5-535) och en sgs1 Top3 mutant (W1058-11C, genotyp, MAT en SUP4-o:: URA3 sgs1-25 Top3-2:: HIS3 ade2-1 can1-100 his3-11, 15 leu2-3, 112 trp1-1 ura3-1 rad5-535) har präglats [3] och var vänligt förutsatt av Dr Rodney Rothstein (Columbia University).
2. Plasmid-DNA konstruktioner
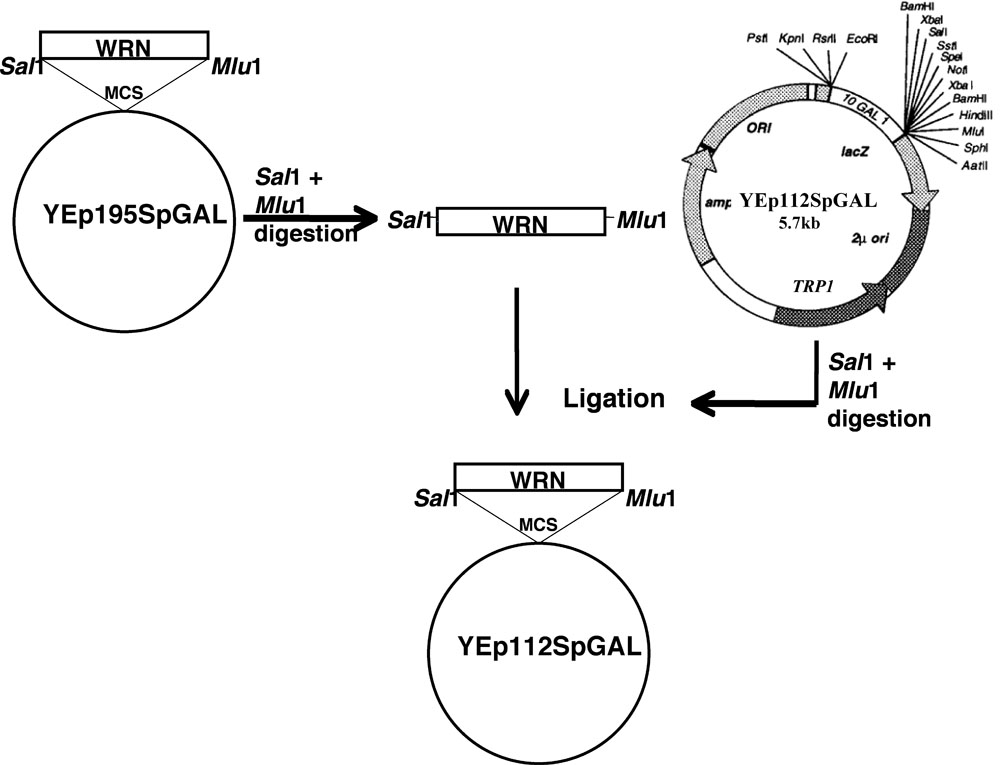
Den WRN gen klonas i YEp112SpGAL efter systemet som diskuterats.
- Site-directed mutationer av WRN (E84A exonuclease döda, K577M helikas döda, R834C och K1016A) i plasmiden YEp195SpGAL-WRN [4] byggdes med hjälp av mutagena primers och ett standardprotokoll från QuickChange II XL site-directed mutagenes kit (Stratagene) genom Löfstrand Labs (Gaithersburg, MD).
- DNA-fragment kodning WRN och dess varianter gel renat från respektive YEp195SpGAL-WRN konstruerar efter dubbla uppslutning med Sal I och MluI I.
- Gel renade fragment har sedan klonats i Sal jag MluI platser i vektor YEp112SpGAL, en 2 ìm flera exemplar plasmid som innehåller en TRP1 markör att bygga YEp112SpGAL WRN eller WRN varianter under kontroll av ett gal-inducerbara promotor som visas i figur 1 .
- Den bygger sålunda erhållna utsågs YEp112SpGAL-WRN, YEp112SpGAL-WRN E84A, YEp112SpGAL-WRN K577M, YEp112SpGAL-WRN K1016A och YEp112SpGAL-WRN R834C.
3. Omvandling
Jäst kulturer odlades enligt standardprotokoll och transformationer utfördes med hjälp av ett litium Acetat-baserat protokoll som Gietz et al [5]. I korthet var följande steg utförs.
- Kulturer av de ovan nämnda stammarna odlades över natten i 5 ml YPD (jästextrakt-pepton-dextros) medium. Kulturer har reinoculated i 50 ml YPD medium och tilläts växa fram en OD 600 1,0 - 1.2 har tagits uppnåtts.
- Kulturer har skördats, tvättas med 20 ml vatten och suspenderade i 1 ml av 100 mm litium acetat (LiAc). Cellerna inkuberades vid 30 ° C i 10 min.
- Cellpelleten var suspenderade i 100 mm LiAc till en slutlig volym på 500 l, vilket räcker för att göra tio transformationer. 50 l av cellsuspensionen var alikvoteras och cellpelleten samlades in genom centrifugering.
- Till cellpelleten lades i följande ordning: 250 l 50% polyetylenglykol (PEG), 36 l 1 M LiAc, 25 l på 2 mg / ml ssDNA (lax spermiens DNA), 5 ml av plasmid-DNA (100 ng 1000 ng), och 50 l vatten. Cellpelleten var vortexed i 1 min.
- Celler sedan inkuberas vid 30 ° C i 30 minuter följt av en värme chock vid 42 ° C i 20 min.
- Celler har skördats och suspenderade i 1 ml vatten. Eftersom plasmiden YEp112SpGAL innehåller trp som genetisk markör, skulle alla de muterade stammar hysa konstruktionen vara TRP skickliga. Därför har transformants utvalda av bordläggning upphävandet på SC (Syntetisk komplett) glukos medier saknar Trp och ruva plattorna vid 30 ° C i 3-4 dagar tills kolonierna visas.
4. Genetisk analys av den långsamma tillväxten fenotypen restaurering WRN omvandlas sgs1 Top3 stam.
1. Streak analys:
- För att undersöka effekten av WRN uttryck på tillväxten av vildtyp föräldrapenning (W303-1A), sgs1 eller sgs1 Top3 stammar, motsvarande stammar med YEp112SpGAL eller YEp112SpGAL-WRN var strimmiga på SC minus Trp plåt innehållande antingen 2% glukos ( GLU) eller 2% galaktos (GAL). Som en kontroll, var sgs1 Top3 / YEp112SpGAL-SGS1 ingår. Plattorna inkuberades vid 30 ° C i 2 dagar.
- För att bestämma förmåga WRN att påverka tillväxten i sgs1 Top3 påfrestningar på lägre uttryck WRN protein nivåer, sgs1 Top3 stam förvandlas med YEp112SpGAL, YEp112SpGAL-WRN eller YEp112SpGAL-SGS1 var streckad på SC minus Trp plåt som innehåller varierande koncentrationer av gal från så låg som 0,005% till så mycket som 2%. Plattorna inkuberas vid 30 ° C i 2 dagar.
- För att bedöma de funktionella kraven i WRN att återställa den långsamma tillväxten fenotyp i sgs1 Top3 bakgrunden, sgs1 Top3 omvandlas med YEp112SpGAL-WRN (E84A, K577M, R834C och K1016A) var streaked på SC minus Trp plåt som innehåller antingen glu eller gal vid angivna koncentrationer. För kontroller var sgs1 Top3 stam förvandlas med YEp112SpGAL, YEp112SpGAL-WRN eller YEp112Sp GAL-SGS1 ingår. Plattorna inkuberas vid 30 ° C i 2 dagar.
2. Flytande kultur analys:
- För att utvärdera tillväxt omvandlas sgs1 Top3 stammar i flytande kultur var jästceller odlas i SC RAF minus Trp vid 30 ° C över natten. Kulturer har reinoculated i SC RAF minus Trp och vuxit vid 30 ° C till tidig logga fas (OD 600 på ~ 0,5). Kulturer har sedan reinoculated vid OD 0,05 i SC media som innehåller 2% gal och tillväxten följdes genom att mäta absorbans vid den angivna tidsintervall.
5. Genetisk analys av hydroxiurea eller methylmethane-sulfonat känslighet i WRN förvandlade jäststammar.
- För att bestämma effekten av WRN uttryck på MMS och HU känslighet sgs1 eller sgs1 Top3 stammar, stammar förvandlas med YEp112SpGAL eller YEp112SpGAL-WRN odlades i SC RAF minus Trp vid 30 ° C.
- Kulturer har reinoculated i SC RAF minus Trp och vuxit vid 30 ° C till tidig logga fas (OD 600 på ~ 0,6 till 0,8).
- Tio gånger spädningar av dessa stammar har sammanställts med SC medelstora saknas Trp. Ur ett exponentiellt växande kultur, var 5 x 10 6 celler tas före och efter utspädning seriellt upp till 10.000 gånger. Fyra mikroliter av varje spädning sågs på SC gal minus Trp plattor som innehåller den angivna koncentrationer av HU eller MMS. Plattorna inkuberades vid 30 ° C.
- Som en kontroll, vildtyp moderstammen (W303-1A) omvandlas med YEp112SpGAL eller YEp112SpGAL-WRN eller sgs1 Top3 / YEp112SpGAL-SGS1 behandlades som beskrivs ovan.
6. Cellcykeln distribution av WRN omvandlas sgs1 Top3 stam.
- För att studera fördelningen cellcykeln av WRN omvandlas sgs1 Top3 stam, kulturer av sgs1 Top3 förvandlas med YEp112SpGAL, YEp112SpGAL-WRN eller YEp112SpGAL-SGS1 och vektor-transformerade vildtyp moderstammen (W303-1A) odlades i SC RAF minus trp vid 30 ° C över natten.
- Kulturer var reinoculated vid OD 0,05 och tillåts växa til tidigt log fas (OD 600 0,5 -0,6). Var WRN uttryck inducerad genom att lägga gal till 2% slutlig koncentration. Kulturer har sedan tillåts växa vid 30 ° C i 6 timmar
- Kulturer har sedan bearbetas för DAPI (4 ", 6-diamidino-2-phenylindole) färgning.
- Kulturer skördades genom centrifugering vid 5800 xgi 10 minuter i rumstemperatur (RT).
- Cellerna tvättades en gång med 1X fosfatbuffrad saltlösning (PBS) och var sedan fast i 70% etanol i 20 min vid RT.
- Fast celler tvättas två gånger med 1 X PBS, och slutligen suspenderade i 1X PBS.
- Celler placerades på bilden och monteras i Vectashield monteringsmedium med DAPI (1 mikrogram / ml) (Vector, USA). Celler undersöktes med en Axiovert 200 M-mikroskop (Zeiss, 100x lins) och komposit differential interferens kontrast (DIC). Fluorescens bilder analyserades med hjälp AxioVision, version 3,0-programmet (Zeiss).
7. Western blot analyser.
1. Att fastställa proteinets uttryck WRN eller WRN varianter omvandlas sgs1 eller sgs1 Top3 stammar var lysates utarbetats av följande metod och proteiner analyseras av immunblotting.
- Omvandlas sgs1 eller sgs1 Top3 stammar odlades i SC RAF minus Trp vid 30 ° C över natten.
- Kulturer har reinoculated och inkuberas vid 30 ° C fram till i början log fas (OD 600 på 0,5 - 0,8) och sedan framkallade genom att lägga gal till 2% slutlig koncentration. Kulturer har sedan tillåts växa vid 30 ° C i 6 timmar
- Celler (3 ml) samlades in genom centrifugering och tvättas med PBS. Cellpelleten var suspenderade i alkaliska lyseringsbuffert [50 mM NaOH, pH 10,5, 2 mM EDTA, 1 mm phenylmethylsulfonyl fluorid (PMSF), 2% SDS, 10% glycerol, 5% 2-merkaptoetanol och proteashämmare (Roche Molecular Biochemicals)] , kokas i 5 min, klargöras genom centrifugering, och neutraliseras med 1 M HCl.
- Proteiner från motsvarande mängder av cell lysat löstes på 8-16% polyakrylamid SDS geler.
2. För att fastställa graden av uttryck av WRN protein under påverkan av olika koncentrationer av galaktos, var kulturer behandlas enligt följande.
- Omvandlas sgs1 Top3 stammar odlades i SC RAF minus Trp vid 30 ° C över natten.
- Kulturer har reinoculated och inkuberas vid 30 ° C fram till i början log fas (OD 600 på 0,5 - 0,8) och sedan framkallade genom att lägga gal till 2% slutlig koncentration. Kulturer har sedan tillåts växa vid 30 ° C i 6 timmar
- Cellerna skördades genom centrifugering och suspenderade i buffert (50 mM Tris-Cl, pH 7,5, 1% natriumdeoxikolat, 1% Triton X-100, 0,1% natriumdodecylsulfat [SDS]) innehållande proteashämmare (0,05% PMSF, 0,05 mikrogram / ml leupeptin).
- Glaspärlor (425 till 600 mikrometer) har lagts till blandningen, och cellerna bröts genom att vortexa.
- Den resulterande homogenatet var centrifugeras i 5 min vid 15 tusen XG och supernatanten fraktionen samlades in.
- Protein i lysat fraktioner uppskattades av Bradford metoden.
- Lika koncentration av protein från olika lysat fraktioner beslutades på 8-16% polyakrylamid SDS geler.
3. Redovisning av WRN eller WRN mutant protein bestämdes genom Western blot med en WRN monoklonal antikropp riktad mot en epitop i renad C-terminala fragment av WRN [6] (1:1000, Spring Valley Labs).
4. Primär antikropp inkubation följdes av inkubering med en sekundär (pepparrotsperoxidaskonjugerade) antikropp (1:5000). Blotting har bearbetats med en ECL Plus västra detektionssystem enligt tillverkarens protokollet.
5. För kvantitativ Western blot-analys, vilket ökar koncentrationen av renade rekombinanta Hans-märkta full längd WRN proteinet fanns med på geler så att när en kvantifiering av blot en vanlig linjär kurva kan genereras. Vidare var cell extrakt från motsvarande mängd celler eller 20 mikrogram av proven jästen lysat lastas på geler att uppskatta koncentrationen av WRN.
6. ImageQuant analys gjordes sedan på geler och standard linjär kurva genererades.
7. Koncentration av WRN protein vid olika gal koncentrationer beräknades med hjälp av vanliga linjära kurvan genereras ovan.
8. Representativa resultat
Alla stammar som används i denna studie är Trp auxotrophs. Därför sgs1, sgs1 Top3 och vild typ W303-1A stammar förvandlas med YEp112SpGAL eller YEp112SpGAL-WRN valdes ut på grundval av deras förmåga att växa i närvaro av SC minus Trp medier. För att undersöka effekten av WRN uttryck på tillväxten av förvandlade sgs1 Top3 muterade celler, var de kulturer strimmiga på tallrikar med SC minus Trp media med 2% gal att inducera WRN uttryck. Top3 decatenates sammanflätade DNA-molekyler som genereras av Sgs1 helikas vid replikering [1, 2] och därför i avsaknad av Top3 är vridande stress inte lindras vilket resulterar i långsam tillväxt och Hyper-rekombination. Den genetiska funktion sgs1 är att undertrycka den långsamma tillväxten fenotyp av en Top3 mutant. Om WRN kan ersätta SGS1 inom genetisk interaktion med Top3 skulle restaurering av Top3 långsam tillväxt fenotyp i sgs1 Top3 förväntas.
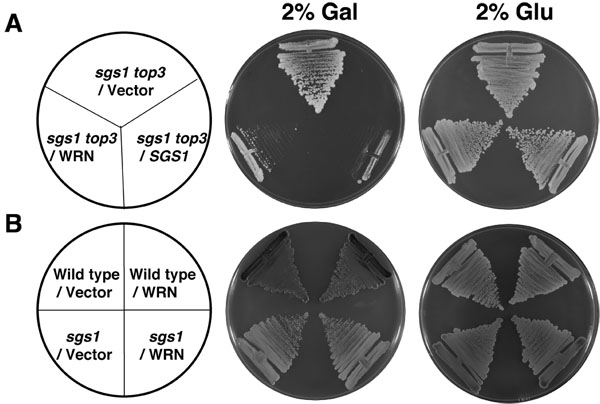
Som framgår av Figur 2A, förvandlade WRN sgs1 Top3 stam växte betydligt långsammare jämfört med vektor transformeras sgs1 Top3 stam. Sgs1 Top3 förändrats med YEp112SpGAL-SGS1 plasmid ingick var som en positiv kontroll (Figur 2A), som visar att vildtyp Sgs1 uttrycks i sgs1 Top3 mutanten kunde genetiskt komplettera tillväxt fenotyp. Den förvandlade sgs1 Top3 stammar växte på liknande sätt i avsaknad av gal som visas under 2 dagar (Figur 2A). Redovisning av WRN hade ingen effekt på tillväxten av föräldrarnas vildtyp (W3031A) eller sgs1 stammar (Figur 2B), vilket indikerar att effekten av WRN på celltillväxt var specifik med vad som observerats i sgs1 Top3 mutant bakgrund. Genetiska studier som utförts med WRN varianter visat att WRN helikas men inte exonuclease aktivitet som krävs för att återställa Top3 tillväxt fenotyp (Figur 3B). Ett naturligt missense polymorfism i WRN som stör helikas aktivitet avskaffas dess förmåga att återställa Top3 långsam tillväxt fenotyp.
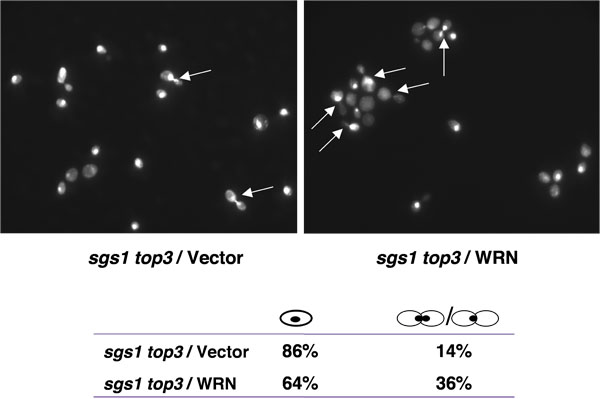
Top3 muterade stammar blir försenad i slutet S/G2 fasen av cellcykeln [1], en egenskap som kan stå för sin långsamma tillväxt. Mutation av SGS1 genen i Top3 bakgrunden undertrycker den försenade S/G2 fasen av cellcykeln. Om WRN uttryck kunde återställa fördröjning i S/G2 fasen av cellcykeln i sgs1 Top3, skulle en ökad population av stora ympade celler med odelade kärnor kan förväntas för sgs1 Top3 muterade celler som uttrycker WRN. Som framgår av Figur 4, var andelen stora ympade celler högre för sgs1 Top3 / WRN än sgs1 Top3 / vektor, vilket tyder på restaurering av S/G2 de låg kännetecknar Top3.
Den sgs1 Top3 dubbla mutant är mindre känslig för MMS eller HU än Top3 enda mutant [2]. Sedan WRN påverkas celltillväxt i sgs1 Top3 vi granskat nästa dess inverkan på känslighet för methylmethane sulfonat (MMS, ett alkylerande medel) och hydroxiurea (HU, en replikering-hämmare). Sgs1 Top3 / WRN visas känslighet för både droger jämförbara sgs1 Top3 / SGS1 (Figur 5). Genetisk analys av WRN varianter visade att WRN helikas / ATPas, men inte WRN exonuclease aktivitet, krävdes för att dess effekt på känsligheten för MMS och HU.

Figur 1. Schematisk presentation för kloning WRN / WRN varianter YEp112SpGAL vektor. Vänligen klicka här för en större version av figur 1.

Figur 2. WRN uttryck i sgs1 Top3 återställer den långsamma tillväxten fenotyp Top3. Panel A, sgs1 Top3 stam förvandlas med YEp112SpGAL eller YEp112SpGAL WRN var strimmiga på en SC-TRP skylt som innehåller antingen 2% glu eller 2% gal. Som en kontroll sgs1 Top3 stam förvandlas med YEp112SpGAL SGS1 var streckad på båda plattorna. Plattorna inkuberades vid 30 ° C i 2 dagar och sedan fotograferas. Panel B, vildtyp moderstammen W303-1A eller sgs1 stam omvandlas med YEp112SpGAL eller YEp112SpGAL WRN var strimmiga på en SC-TRP plåt som antingen innehåller 2% glu eller 2% gal . Plattorna inkuberades vid 30 ° C i 4 dagar och sedan fotograferas. Sammansättning av plattorna var i panel A och panel B respektive. Vänligen klicka här för att se en större version av figur 2.

Figur 3. WRN ATPas / helikas, men inte exonuclease aktivitet krävs för att återställa den långsamma tillväxten fenotyp Top3 i sgs1 Top3 bakgrunden. Sgs1 Top3 stam förvandlas med ATPas / helikas-döda (YEp195SpGAL WRN K577M), exonuclease-döda (YEp195SpGAL- WRN E84A) var RQC mutant (YEp195SpGAL-WRN K1016A) eller polymorf mutant (YEp195SpGAL-WRN R834C) streckad på SC-TRP plattor innehållande antingen 2% glu (Panel C) eller 2% gal (panel B). Plattorna inkuberades vid 30 ° C i 2 dagar och sedan fotograferas. Sammansättning av plattorna var i panel a. Vänligen klicka här för att se en större version av figur 3.

Figur 4. WRN uttrycket framkallar S/G2 greps i sgs1 Top3 celler. Logaritmiskt växande kulturer sgs1 Top3 stam förvandlas med YEp112SpGAL, YEp112SpGAL WRN eller YEp112SpGAL SGS1 och vektor-transformerade vildtyp moderstammen var framkallade på 2% gal koncentration för 6 h. Kulturer har skördats, behandlas för DAPI färgning som beskrivs i Material och Metoder och sågs använda Axiovert 200 M mikroskop (Zeiss, 100x objektiv). Visas är DAPI färgning av sgs1 Top3 förvandlas med YEp112SpGAL (övre vänster) och med YEp112SpGAL WRN (övre höger). Pilarna visar celler med odelade kärnor. Fördelning av celler i G1 (enskilda celler) och S/G2 (ympade celler) visas i nedre panelen. Vänligen klicka här för att se en större version av figur 4.

Figur 5. Effekt av WRN uttryck på MMS och HU känslighet sgs1 Top3 påfrestningar. Växande Logaritmiskt kulturer sgs1 Top3 stam förvandlas med YEp112SpGAL, YEp112SpGAL WRN, exonuclease-döda (YEp112SpGAL-WRN E84A), ATPas / helikas-döda (YEp112SpGAL WRN K577M) RQC mutant (YEp112SpGAL-WRN K1016A), polymorf mutant (YEp112SpGAL-WRN R834C), omvandlas YEp112SpGAL SGS1 och vektor vildtyp moderstammarna sågs i en tio gånger seriespädningar på SC-TRP plattor som innehåller glu eller tjej och antingen MMS eller HU vid angivna koncentrationer. Plattorna inkuberades vid 30 ° C under 2 dagar (kontroll plattor) och 4 dagar (MMS och HU tallrikar) och sedan fotograferas. Vänligen9/1639fig5.jpg "> klicka här för att se en större version av figur 5.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
En av styrkorna med att använda jäst som modellsystem är tillgången på mutanter i definierade DNA-replikation och vägar reparation som bevaras mellan jäst och människa. Ytterligare urval av transformants hysa de specifika gener är enkelt och tillförlitligt eftersom laboratoriestammar är auxotrophic mutanter och vektorer med auxotrophic markörer är lätt tillgängliga. Med hjälp av dessa vektorer uttrycket av genen produkter kan regleras genom att placera dem under kontroll av en inducerbara promotor (t.ex. gal inducible promotor, etc.). Med tanke på dessa fördelar har vi utvecklat en jäst baserade modellsystem för att studera de funktionella kraven på mänskliga WRN genen defekt i Werner syndrom i en väg som potentiellt är bevarad mellan människa och jäst. Med hjälp av beskrivna metoder, förutsatt att vi det första beviset att WRN kan fungera i en genetisk väg som påverkar Top3-relaterade fenotyper. Våra observationer föranleda ytterligare utredning av möjligheten att WRN funktionellt interagerar med Top3α i mänskliga celler under cellulärt DNA-replikation eller rekombination. Möjligen, i en BLM-nedsatt kondition, kan WRN ersätta delvis för BLM genom sitt protein partnerskap med en topoisomeras.
Subscription Required. Please recommend JoVE to your librarian.
Acknowledgments
Detta arbete stöddes fullt ut av den Interna Forskningsprogram NIH, National Institute on Aging. Vi tackar Dr Rodney Rothstein (Columbia University) för jäststammar och Dr Brad Johnson (University of Pennsylvania School of Medicine, Philadelphia) för SGS1 uttryck plasmiden.
References
- Aggarwal, M., Brosh, R. M. Jr WRN helicase defective in the premature aging disorder Werner Syndrome genetically interacts with Topoisomerase 3 and restores the top3 slow growth phenotype of sgs1 top3. Aging. 1, 219-233 (2009).
- Gangloff, S., McDonald, J. P., Bendixen, C., Arthur, L., Rothstein, R. The yeast type I topoisomerase Top3 interacts with Sgs1, a DNA helicase homolog: a potential eukaryotic reverse gyrase. Mol Cell Biol. 14, 8391-8398 (1994).
- Shor, E., Gangloff, S., Wagner, M., Weinstein, J., Price, G., Rothstein, R. Mutations in homologous recombination genes rescue top3 slow growth in Saccharomyces cerevisiae. Genetics. 162, 647-662 (2002).
- Sharma, S., Sommers, J. A., Brosh, R. M. Jr In vivo function of the conserved non-catalytic domain of Werner syndrome helicase in DNA replication. Hum Mol Genet. 13, 2247-2261 (2004).
- Gietz, R. D., Schiestl, R. H., Willems, A. R., Woods, R. A. Studies on the transformation of intact yeast cells by the LiAc/SS-DNA/PEG procedure. Yeast. 11, 355-360 (1995).
- von Kobbe, C., Thoma, N. H., Czyzewski, B. K., Pavletich, N. P., Bohr, V. A. Werner syndrome protein contains three structure specific DNA binding domains. J Biol Chem. 278, 52997-53006 (2003).

{kind=link}
{kind=link}
{kind=link}
{kind=link}