

Summary
Nous fournissons une méthode reproductible pour la culture des monocouches confluentes de cellules foetales humaines épithélium pigmentaire rétinien (hfRPE) cellules qui présentent la morphologie, la physiologie, la polarité et de protéines et les profils d'expression génique du tissu natif adulte. Ce travail a été étendue à un modèle animal de plusieurs maladies oculaires.
Abstract
Nous avons développé une procédure de culture cellulaire qui peut produire de grandes quantités de monocouches confluentes de fœtus humains primaires de l'épithélium pigmentaire rétinien (hfRPE) les cultures avec des caractéristiques morphologiques, physiologiques et génétiques de la RPE humaine native. Ces cultures de cellules pigmentaires présentent hfRPE lourds, et la microscopie électronique montrent vaste microvillosités membrane apicale. Les complexes de jonction ont été identifiés avec l'étiquetage immunofluorescence des différentes protéines de jonctions serrées. Polarité épithéliale et la fonction de ces cultures facilement reproductible primaires ressemblent déjà étudié les modèles de mammifères de l'EPR indigènes, y compris humaines. Ces résultats ont été étendues par le développement d'interventions thérapeutiques dans plusieurs modèles animaux de la maladie de l'œil humain. Nous nous sommes concentrés sur les stratégies pour l'élimination de l'accumulation de liquide anormal dans la rétine ou de l'espace sous-rétinien. L'espace extracellulaire sous-rétinienne sépare les segments externes des photorécepteurs et la membrane apicale de l'EPR et est essentiel pour l'entretien des pièces jointes de la rétine et une multitude de RPE / rétine interactions.
Protocol
1. Tissu foetal humain
Toutes les recherches tissus humains liés suit les principes de la Déclaration d'Helsinki et le conseil d'examen institutionnel NIH. Yeux du fœtus sont obtenus par une proxénète indépendante, Advanced Bioscience Ressources (ABR, Alameda, Californie), à partir de donneurs choisis au hasard à 16 à 22 semaines de gestation, placés dans un milieu RPMI-1640 contenant des tubes (fournis par l'ABR), emballés sur glace, et livrés par un service de livraison prioritaire pendant la nuit. Les tissus restent viables jusqu'à 48 heures après l'énucléation.
2. Milieu de culture cellulaire
MEM-alpha milieu modifié (Sigma-Aldrich) est utilisé comme milieu de base pour préparer 5% et 15% de sérum contenant des supports de culture des cellules de l'EPR (moyenne RPE; tableau 1 ci-dessous).
| Nom | Sigma | Gibco | Montant | Stockage |
| MEM, l'alpha de modification | M-4526 | 500 ml | 4 ° C | |
| N1 complément | N-6530 | 5 ml | 4 ° C | |
| Pénicilline-streptomycine | 15140-148 | 5 ml | -20 ° C | |
| Glutamax - I | 35050 | 5 ml | -20 ° C | |
| Acides aminés non essentiels | M-7145 | 5 ml | 4 ° C | |
| * THT | -80 ° C | |||
| Taurine | T-0625 | 125 mg | ||
| Hydrocortisone | H-0396 10 | 10 ug | ||
| Triiodo-thyronin | T-5516 | 0,0065 mg | ||
| Fœtal bovin ** sérique | 5% ou 15% | -80 ° C |
Tableau 1. Fœtal humain composants du milieu EPR pour la préparation de 500 ml de milieu
THT * est faite en dissolvant la taurine-hydrocortisone-triiodo-thyronin en 1 de 1,5 mL de PBS avant de prendre la moyenne. Aliquotes multiples sont faites et stockées à -80 ° C à simplifier la préparation de culture du milieu de culture.
** Sérum fœtal bovin n'est pas obtenue chez Sigma-Aldrich ou Gibco.
Sérum fœtal bovin utilisé dans la préparation des médias est obtenu à partir d'Atlanta Biologicals (Norcross, GA). Chaque bouteille de sérum est inactivé par la chaleur (56 ° C pendant 1h) avant utilisation. Pour assurer la cohérence des montants de culture de cellules grandes de sérum provenant du même lot sont achetés et stockés dans -20 ° C jusqu'à ce qu'ils soient utilisés pour la préparation des supports. Cellules clairsemée ensemencé à partir d'un lot spécifique de cellules sont cultivées dans hfRPE 24 - ou 12 - plaques ainsi pendant quelques semaines dans un milieu contenant 20% de FBS de lots différents. Les cellules les plus dynamiques melanated avec la morphologie RPE plus classique (pavés) indiquent un terrain approprié sérum qui peut être utilisé pour la culture cellulaire.
Milieu de culture cellulaire contient également: N1 supplément (Sigma-Aldrich) 1:100 mL / mL, glutamax / pénicilline-streptomycine 1:100 mL / mL (Gibco), et non essentiels solution d'acides aminés (Sigma-Aldrich) 1:100 mL / mL. En outre, l'hydrocortisone (20 mg / L), de la taurine (250 mg / L), et triiodo-thyronin (0,013 mg / L) (THT) est préparée à l'avance par la dissolution de ces trois composantes dans le PBS à une concentration finale de 1:500 (ml / ml). Des aliquotes de THT sont stockés à 80 ° C jusqu'à ajoutés au milieu de l'EPR.
3. Culture de cellules
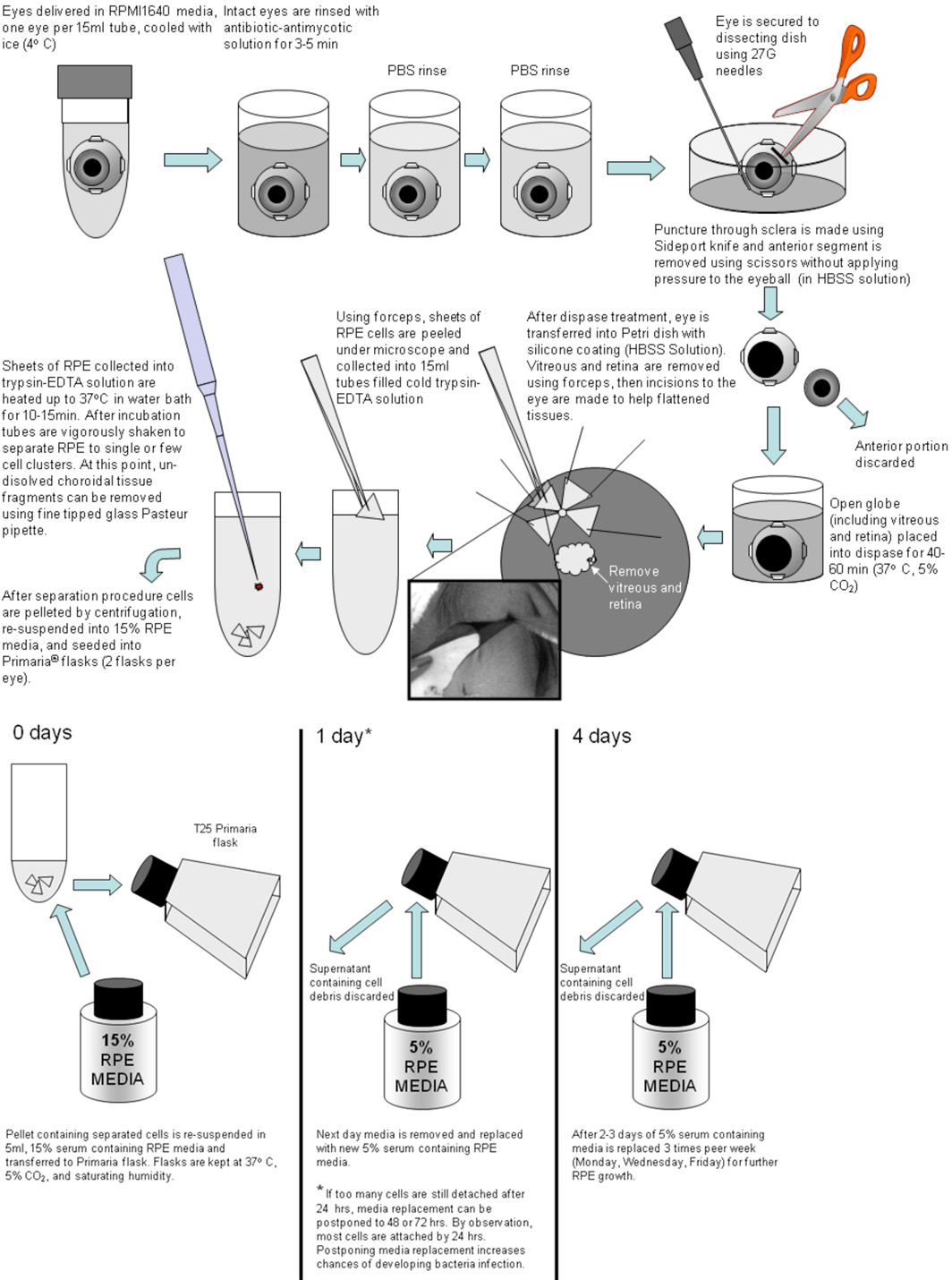
Dès réception, les globes intacts sont rincés dans une solution antibiotique antifongique (dilué 10X;.. Réf 15240-096; Invitrogen) plus gentamicine (1 mg / ml) pendant 3 à 5 minutes (voir figure 1 ci-dessous). 
Figure 1. Après une incubation d'antibiotiques sont rincées deux fois avec support tel que HBSS ou PBS. Un globe oculaire à l'époque est transféré à 10 cm boîte de Petri avec des enduits de Sylgard-184 (WPI) et sécurisé avec 27G aiguilles. En utilisant un couteau à blanc Sideport (Alcon), une incision est faite à travers la sclère en dessous du corps ciliaire (1 / 3 de la distance de l'équateur oeil à la face antérieure). Cette incision est utilisée pour démarrer une découpe circulaire pour l'enlèvement de la partie antérieure de l'œil. Cette coupe est faite en utilisant un revêtement de carbure de tungstène courbes ciseaux à iris avec une lame dentelée (FST). Avant l'enlèvement de la partie antérieure de l'œil, une coupe est faite à travers le corps vitré pour éviter de détacher la retina de l'EPR au pôle postérieur. Après la partie antérieure de l'œil est enlevé, le pôle postérieur est incubée avec dispase-I solution (2 U / mL, n ° 04942086001;.. Roche Diagnostics, Indianapolis, IN) dans 5% de sérum de milieu contenant pendant 40-60 minutes à 37 ° C-CO 2 5%. Après le traitement dispase, les pôles postérieurs sont transférés à une HBSS en boîtes de Petri avec du silicone rembourrage (Sylgard 184; Dow Corning, Midland, MI) et disséqués en quadrants ou de plus grands morceaux de tissus suffisamment aplatir. Puis la rétine est délicatement retiré avec une pince. Couches RPE unicellulaires ont été décollée en feuilles et collectées directement en froid trypsine-EDTA (Gibco, # 25200-056) solution. Après l'EPR sont collectées, les tubes avec les tissus de la trypsine-EDTA sont scellés et transférés dans un bain d'eau pendant 10-15 minutes à 37 ° C. Après 10 min d'incubation, les tubes sont secoués vigoureusement pour séparer les EPR en petites grappes. Si la séparation n'est pas complète, les tubes sont replacés dans un autre bain d'eau pendant 5 mn. Après la trypsine-EDTA incubation, les tubes sont inspectés pour d'éventuelles non-dissous amas de cellules mixtes. Tout amas observés sont enlevés à l'aide fines pointes pipette Pasteur en verre. Après filer vers le bas (1,4 RPM sur centrifugeuse clinique pendant 4 minutes), les cellules hfRPE sont remis en suspension dans les médias RPE 15% et ensuite mis dans des flacons Primaria (exemple: cat pas 08-772-45; Fisher Scientific, Pittsburgh, PA.. ). Ce milieu est remplacé après un jour avec un milieu de RPE 5% de sérum contenant, et les modifications ultérieures ont été faites tous les 2 à 3 jours. Après 3 à 4 semaines, les cellules deviennent confluentes et uniformément pigmentés. Elles sont ensuite traitées à la trypsine à 0,25% de trypsine-EDTA pour les 10 à 15 minutes, remises en suspension dans 15% de sérum contenant un milieu de culture cellulaire RPE, et ensemencées sur des clairs inserts de culture cellulaire de 150 à 200K cellules par puits (Transwell; Corning Costar, Corning, NY), utilisant des plaquettes de diamètre de 12 mm, pores de 0,4 um, les membranes de polyester (exemple: cat pas 07-200-161, Fisher Scientific)... Avant le semis, les puits ont été revêtus de la matrice extracellulaire humaine (10 ug dans 150 ul par puits HBSS, n ° 354237;.. BD Biosciences, Franklin Lakes, NJ) et guéri par la lumière UV dans le capot pendant 2 heures. Dans certains cas, la procédure de traitement à la trypsine a été répété pour une deuxième fois, pour recueillir les cellules qui ne se détachent pas après la trypsinisation premier. Le même protocole (hors revêtement avec ECM) a été utilisé pour des cellules en culture sur les flacons de générer de la population de cellules P1. Ces cellules ont été utilisés dans des expériences où ils avaient une résistance des tissus au total ≥ 200 Ω • cm 2 et ont été uniformément pigmentés.
S'il vous plaît cliquer ici pour agrandir la figure 1 .
4. Etape par étape les procédures
- Préparer à 12 puits plaque avec des solutions multiples (pour chaque oeil 4 puits nécessaire):
- Ajouter 10x solution antibiotique antifongique - 1 puits / les yeux
- Ajouter PBS / HBSS solution pour rincer - 2 puits / les yeux
- ajouter la solution dispase - 1 puits / les yeux
- Préparer la dissection boîte de Petri par le déballage 5 aiguilles de fixation et de le remplir avec du HBSS
- Déballez les yeux et les placer dans antibiotiques antimycosiques solution pour 3-5 minutes (préparé à l'étape 1)
- Rincer les yeux dans les deux puits (préparé à l'étape 1) avec du PBS / HBSS, puis de les transférer à disséquer plat.
- Garniture musculaire excessive et les tissus conjonctifs autour de l'oeil
- Utiliser des aiguilles 27G sécurisé (épingler au silicium de base) les yeux dans la dissection plat en les alignant dans un chemin où la cornée vers le haut.
- Aide d'un couteau faire une incision Sideport ci-dessous la cornée, où découpe circulaire va commencer
- Avec des ciseaux iris font coupées autour de l'œil, puis l'aide de ciseaux couper à travers la même vitreux et soulever la portion antérieure de l'oeil loin.
Remarque: dans les étapes 3 à 8 éviter toute pression excessive mécanique globe oculaire. - Transfert yeux ouverts dans la solution dispase (préparé à l'étape 1)
- Incuber œilleton pendant 40-60 minutes à 37 ° C avec 5% de CO 2
- Remplacer solution HBSS à disséquer plat avec un nouveau.
- Transfert des yeux de la solution dispase à plat de dissection.
- Position dans l'œil boîte de Pétri (tasse globe oculaire vers le haut) et le fixer avec deux aiguilles 27G.
- Soulevez doucement la rétine partiellement séparés et coupés avec des ciseaux la rétine rétine loin du nerf optique. Jeter rétine.
- Avec des ciseaux iris faire une incision de la périphérie de l'œil vers le nerf optique.
- Utiliser tous les cinq aiguilles 27G aplatir la couche RPE oeil faisant bien tendue.
- Avec des ciseaux rétiniennes font coupées circulaire autour du nerf optique qui sépare la couche RPE de l'attachement à nerf optique.
Remarque: les étapes 13 à 17 peut être fait avec un faible grossissement ou sans microscope stéréoscopique - Ajustez le grossissement 250x stéréomicroscopeou plus et de trouver bord de la feuille EPR à proximité du nerf optique le long de la coupure faite par les ciseaux à iris.
- En utilisant deux pinces membrane séparée RPE-Bruch de la couche de tissu choroïdien. Elle peut exiger un peu de tentatives pour trouver des zone le long du bord où la connexion de l'EPR et la choroïde est la plus faible.
- Placer dans des feuilles RPE froide trypsine-EDTA solution dans tube de 15 ml
- Après l'EPR sont recueillis, des tubes à bouchon et le transfert des tissus dans la trypsine-EDTA dans le bain d'eau pour 10 à 15mn à 37 ° C.
- Après 10 min d'incubation, agiter vigoureusement tubes de 15 ml de séparer l'EPR en petites grappes. Si la séparation n'est pas complète, placer les tubes de retour dans le bain d'eau pendant 5 mn.
- Inspectez les tubes pour d'éventuelles non-dissous amas de cellules mixtes. Tout amas observés doivent être retirés à l'aide fines pointes pipette Pasteur en verre.
- Isoler (1,4 RPM sur centrifugeuse clinique pendant 4 mn) hfRPE cellules, enlever le surnageant et remettre en suspension les cellules dans 15% des médias RPE (9 mL au total)
- Mettez 3 ml de suspension cellulaire dans des flacons Primaria ajouter 2 mL de Fresh Media RPE 15%, flacon placer dans pépinière jusqu'au lendemain (37 ° C, 5% CO 2)
5. Les résultats représentatifs
Validation fonctionnelle de la culture cellulaire hfRPE
Dans les expériences précédentes, nous avons utilisé ces cultures primaires de déterminer l'expression, la polarisation et la fonction des protéines plasmatiques de transport membranaire et d'identifier des voies de signalisation qui régulent la fonction EPR 1-11 Ces caractéristiques car elles sont accumulées fournir un ensemble de validation des propriétés qui peuvent être appliquées à chaque génération cellulaire. Les expériences résumées ci-dessous identifier les protéines qui déterminent le transport transépithélial du liquide et l'intégrité de la voie paracellulaire. Ces expériences in vitro ont été validés dans un modèle animal de la rétine nouveau rattachement 6.

Figure 2. Localisation de la protéine CFTR dans les cellules hfRPE panneau supérieur gauche:. CFTR a été détecté dans les cellules en utilisant une membrane hfRPE enrichi d'extraits. M, marqueur de poids moléculaire; piste 1, les principales matures (bande C) et immatures (bandes A et B) Panneau central faible: la localisation par immunofluorescence de la protéine CFTR (étiquette verte) Haut panneau de droite: vue agrandie en coupe par le plan z montrant.. intensité maximale de projection à travers l'axe z. ZO-1 (étiqueté comme rouge) sert de marqueur de jonctions serrées délimitant les côtés apical et basolatéral de l'EPR. DAPI (bleu) les étiquettes des noyaux situés à proximité de la membrane basale. ZO-1 apparaît comme jaune / orange, car la fluorescence de ses chevauchements label rouge vert de la protéine CFTR.

Figure 3. IFN induit des changements physiologiques dans hfRPE A:. Ajout de IFN au bain basale accrue de transport de fluides transépithélial (J v) à travers des monocouches primaires, des cellules en culture hfRPE J v est tracé en fonction du temps dans la trace en haut et fluide net. d'absorption (apicale à bain d'basale) est indiqué par des valeurs positives; potentiel transépithélial (TEP) et la résistance des tissus au total (R T) sont tracées en fonction du temps dans les traces du bas. B: Ajout de CFTRinh-172 (5 um) pour le bain basale inhibé l'IFN-J v stimulé augmenter.
Voici un exemple d'expériences modèles animaux confirment hfRPE résultats in vitro. Dans cette expérience, créées artificiellement décollement de la rétine a été considérablement réduite après l'addition de IFNg à la surface de l'œil extérieur (Figure 4 A, B). Cet effet peut être partiellement bloquée par: (1) ajout de bloqueur AMPc (figure 4D), (2) a complètement bloqué après ajout de bloqueurs JAK AMPc +. Images ci-dessous sont obtenus en utilisant scanner octobre

Figure 4. Expériences in vivo décollement de la rétine. Les procédures pour ces expériences in vivo ont été précédemment décrit en détail 6. Dans ces expériences, décollement de la rétine ont été créés dans l'oeil de rat par injection de 0,5-3 ul de PBS modifiés solution dans l'espace sous-rétinien (SRS), seul ou avec une combinaison de JAK-STAT et inhibiteurs de la voie PKA. Chaque expérience était une période de contrôle initial de 40-70 min après la création du décollement de rétine. Pendant ce temps, le taux de variation de volume de bulle a été mesurée afin d'assurer la stabilité bulle. Tomographie par cohérence optique d'imagerie (Institut de Physique Appliquée de l'Académie russe des sciences, de Nijni-Novgorod, en Russie) a été utilisé pour mesurer l'évolution temporelle de la variation de volume dans SRS.
Le cours du temps de re-attachement volchangement de UME a été mesuré par tomographie par cohérence optique (OCT). Images OCT de 4 expériences différentes (panneaux sur la gauche; AD) qui montrent un changement dans les tailles de détachement (flèches en A, B et D) après l'ajout de IFN à la surface antérieure de 40 à 70 min. Les panneaux A et B montrent que IFN augmenté Jv de son taux de contrôle (≈ 2 pl • cm 2 • h -1) à 14 et 12 cm 2 ul • • h-1, respectivement. Après ajout de la voie JAK-STAT et de la PKA de la sérotonine (C et D), l'IFN - Les taux d'absorption induites ont été significativement réduite à 0,2 et 7,9 cm 2 ul • • h-1, respectivement. Les flèches indiquent la limite de la bulle pour la comparaison avec la zone délimitée à l'intérieur de la ligne pointillée, (volume de départ). Le côté droit de la figure: en haut du panneau-sommaire des taux Jv mesuré à partir de plusieurs expériences; milieu du panneau-film ( cliquez ici ); panneau inférieur 3D-sections d'expérience résumée dans B. PseudoColor en bleu indique l'étendue spatiale de détachement à l'instant t = 0 et 40 min après l'addition de INFγ.
Toutes les expériences animales ont été menées en conformité avec l'Association for Research in Vision et la déclaration d'ophtalmologie. Le protocole a été approuvé par le Comité de protection des animaux et de l'utilisation des National Institutes of Health.
Discussion
Dans les présentes expériences, nous décrivons des modifications supplémentaires de nos techniques déjà publié 3 conçu pour simplifier les multiples étapes nécessaires à la production cohérente hfRPE cultures primaires avec un nombre élevé de cellules disponibles pour chaque œil. Chaque changement dans la procédure initiale a été rigoureusement testés dans la physiologie et de multiples expériences de biologie moléculaire afin de s'assurer que les changements ne pas introduire d'artefacts et d'être constamment testée par de nombreux autres laboratoires utilisant ces cellules. Enfin, des expériences complémentaires ont été effectuées pour comparer les résultats obtenus in vitro à des perturbations similaires dans des modèles animaux.
Disclosures
Aucun conflit d'intérêt déclaré.
Acknowledgments
Nous remercions les membres Lab Jeffrey Adijanto, Tina Banzon, Rong Li, Qin Wan, Congxiao Zhang, Jing Zhao, Connie Zhi, Awais Zia, Natalia Strunnikova pour les aider à la caractérisation de ces cultures de cellules. Un merci spécial à Jing Zhao, Connie Zhi, et Tina Banzon pour leur aide dans le maintien de stocks importants de cultures cellulaires.
Ce travail a été soutenu par le National Institutes of Health Research Programme intra-muros.
Materials
| Name | Company | Catalog Number | Comments |
| Instruments needed for dissection protocol | |||
| Stereo Microscope - any available with working magnification x250 needed for dissection | |||
| Dissecting dish - e.g. Kimble 100 x 20mm #23062 (one per dissection) any source | |||
| Sylgard 184 - WPI Cat. #SYLG184* | |||
| * To prepare dissecting dish follow Sylgard 184 included mixing instructions, pour mixed liquid elastomer into Petri dish to form 5-8mm layer. Allow elastomer to cure for at least 24hrs. Dish with cured elastomer can be sterilized using 70% ethanol and reused multiple times (~100 times) | |||
| 27 G needles from B-D PrecissionGlide 1¼ length (5 per dissection) any source | |||
| HBSS 1X Solution containing Ca and Mg salts - GIBCO Cat.#14025 500ml (good for multiple dissections) | |||
| Iris scissors carbide serrated blade, curved - FST Cat.# 14559-11 (one) | |||
| Retinal scissors - Katena Cat.#K4-5300 (one) | |||
| Forceps - e.g. Dumont Tweezers #5 with polished tips from WPI Cat.#500085 (2 pieces) | |||
| Sideport knife - Alcon, ClearCut 1mm, Dual Bevel, angled Cat.#8065921540 | |||
| Centrifuge tube 15ml - (one per eye) any source | |||
| Pasteur glass pipette - (one per dissection) any source | |||
| Plate 12 well (one per dissection) any source | |||
| Primaria 25cm2 flask - (2-3 per eye) any source | |||
References
- Bharti, K., Miller, S. S., Arnheiter, H. The new paradigm: Retinal pigment epithelium cells generated from embryonic stem cells or induced pluripotent cells. Pigment Cell & Melanoma Research. , Forthcoming Forthcoming.
- Strunnikova, N. V., Maminishkis, A., Barb, J. J., Wang, F., Zhi, C., Sergeev, Y., Chen, W., Edwards, A. O., Stambolian, D., Abecasis, G., Swaroop, A., Munson, P. J., Miller, S. S., S, S. Transcriptome analysis and molecular signature of human retinal pigment epithelium. Hum Mol Genet. 19 (12), 2468-2486 (2010).
- Miller, S. S., Maminishkis, A., Li, R., Adijanto, J. Chapter 184: Phototransduction: RPE transport Retina Phototransduction: RPE transport. Encyclopedia of the Eye. , Elsevier. 2540-2540 (2010).
- Bryant, D. M., Mostov, K. E. From cells to organs: Building polarized tissue. Nature Reviews. Molecular Cell Biology. 9 (11), 887-901 (2008).
- Maminishkis, A., Chen, S., Jalickee, S., Banzon, T., Shi, G., Wang, F. E., Ehalt, T., Hammer, J. A., Miller, S. S. Confluent monolayers of cultured human fetal retinal pigment epithelium exhibit morphology and physiology of native tissue. Invest Ophthalmol Vis Sci. 47, 3612-3624 (2006).
- Shi, G., Maminishkis, A., Banzon, T., Jalickee, S., Li, R., Hammer, J., Miller, S. S. Control of chemokine gradients by the retinal pigment epithelium. Invest Ophthalmol Vis Sci. 49, 4620-4630 (2008).
- Economopoulou, M., Hammer, J., Wang, F. E., Fariss, R., Maminishkis, A., Miller, S. S. Expression, localization, and function of junctional adhesion molecule-C (JAM-C) in human retinal pigment epithelium. Invest Ophthalmol Vis Sci. 50 (3), 1454-1463 (2008).
- Li, R., Maminishkis, A., Banzon, T., Wan, Q., Jalickee, S., Chen, S., Miller, S. S. IFN{gamma} Regulates Retinal Pigment Epithelial Fluid Transport. Am J Physiol Cell Physiol. 297, C1452-C1465 (2009).
- Li, R., Maminishkis, A., Wang, F. E., Miller, S. S. PDGF-C and -D induced proliferation/migration of human RPE is abolished by inflammatory cytokines. Invest Ophthalmol Vis Sci. 48, 5722-5732 (2007).
- Maminishkis, A., Jalickee, S., Blaug, S. A., Rymer, J., Yerxa, B. R., Peterson, W. M., Miller, S. S. The P2Y(2) receptor agonist INS37217 stimulates RPE fluid transport in vitro and retinal reattachment in rat. Invest Ophthalmol Vis Sci. 43, 3555-3566 (2002).
- Wang, F. E., Zhang, C., Maminishkis, A., Dong, L., Zhi, C., Li, R., Zhao, J., Majerciak, V., Gaur, A. B., Chen, S. MicroRNA-204/211 alters epithelial physiology. FASEB J. 24, (2010).
- Adijanto, J., Banzon, T., Jalickee, S., Wang, N. S., Miller, S. S. CO2-induced ion and fluid transport in human retinal pigment epithelium. J Gen Physiol. 133, 603-622 (2009).
- Li, R., Maminishkis, A., Zahn, G., Vossmeyer, D., Miller, S. S. Integrin alpha5beta1 mediates attachment, migration, and proliferation in human retinal pigment epithelium: relevance for proliferative retinal disease. Invest Ophthalmol Vis Sci. 50, 5988-5996 (2009).
- Peterson, W. M., Meggyesy, C., Yu, K., Miller, S. S. Extracellular ATP activates calcium signaling, ion, and fluid transport in retinal pigment epithelium. J Neurosci. 17, 2324-2337 (1997).
- Voloboueva, L. A., Liu, J., Suh, J. H., Ames, B. N., Miller, S. S. R)-alpha-lipoic acid protects retinal pigment epithelial cells from oxidative damage. Invest Ophthalmol Vis Sci. 46, 4302-4310 (2005).

{kind=link}