

Summary
の神経筋接合部(NMJ)
Abstract
ショウジョウバエ幼虫神経筋接合部(NMJ)は、シナプスの構造と機能の研究のための優れたモデルである。 ショウジョウバエは、強力な遺伝子操作の容易さのためによく知られており、幼虫の神経系は正常な機能にも乱れていないだけの研究に特に役立っていますそれは、いくつかの神経疾患(ロイドとテイラー、2010年)同行。 ショウジョウバエに見られる多くの重要なシナプス分子はまた、哺乳類で発見され、ほとんどのCNS のような哺乳動物ではシナプスを興奮している、 ショウジョウバエ NMJはグルタミン酸作動性であり、活性依存性リモデリング(島ら 、2000)を示しています。それらの神経支配のパターンは、ステレオタイプとそれが可能などの運動ニューロンとそれが支配することが体壁筋線維(Keshishianと金、2004)との間のものとして特定されたシナプスの端末を、勉強することの繰り返しなので、さらに、 ショウジョウバエの神経細胞を個別に識別することができます。遺伝子と物理操作の容易さとともに、進化的に保存シナプスのコンポーネントの存在は、シナプス機能(Budnik、1996)基礎となるメカニズムを調べるためのショウジョウバエモデルに最適です。
これらは神経伝達物質の放出の部位であるので、シナプスの端末でアクティブなゾーンは、特に興味深いものである。 NC82は、 ショウジョウバエのタンパク質Bruchpilot(BRP)、アクティブゾーン(Wagh ら 、2006)の重要なコンポーネントであるCAST1/ERC家族を認識するモノクローナル抗体である。 BRPは、直接、アクティブなゾーンのT -バーを形成することが示さと効果的にTバーの密度の下のCa 2 +チャネル(フーケら 、2009)クラスタリングを担当した。 BRPの変異体は、Ca 2を削減しました+チャネルの密度は、落ち込んで誘発小胞放出、および変更された短期可塑性(キッテルら 、2006)。アクティブなゾーンへの変化は、 ショウジョウバエの疾患モデルで観察されている。例えば、NC82抗体を用いた免疫蛍光は、アクティブなゾーンの密度が筋萎縮性側索硬化症とピット-ホプキンス症候群のモデルで減少したことを示した(Ratnaparkhi ら、2008;。。Zweierら 、2009)。したがって、アクティブなゾーン、または他のシナプス蛋白質の評価は、疾患のショウジョウバエの幼虫のモデルでシナプスの欠陥の存在の貴重な初期の手がかりを提供することがあります。
NMJの免疫蛍光分析のための準備ホールマウント解剖ショウジョウバエの幼虫は、いくつかのスキルが必要ですが、少し練習すれば、ほとんどの科学者によって行うことができます。発表は各遺伝子型間の環境の違いを制限し、再現性と統計分析の信頼性のために十分な動物を提供する、解剖と同じ解剖皿の中で免疫染色するために複数の幼虫のために提供する方法です。
Protocol
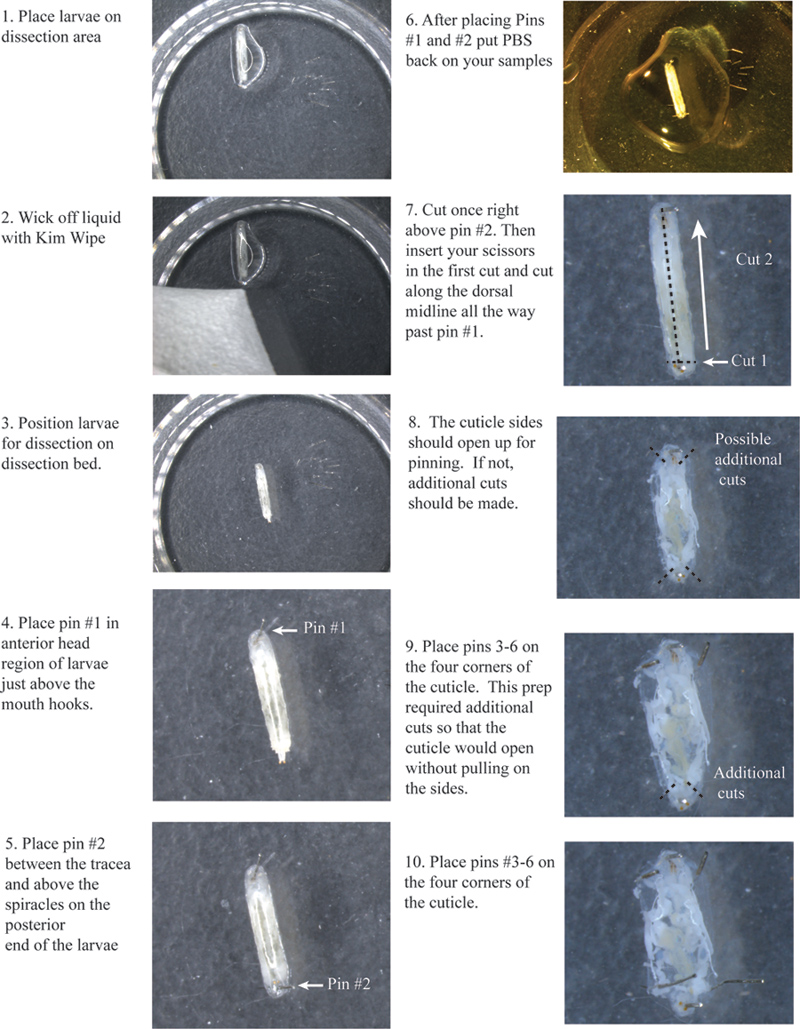
1。免疫蛍光のための準備:
- 解剖表面を作成するには、小規模な組織培養プレートにSylgard 184シリコーンエラストマーベースを注ぐ。解剖領域がリムよりも低くなるように完全にプレートを埋めるためではないことを確認してください。
- 操作の増加を容易にするために約3.5mmの長さに解剖ピンを取り除いて、幼虫ごとに少なくとも6ピンを持っていることを確認してください。
- あなたは、解剖用のピンを把握できるようになります鈍鉗子が必要になります。
2。幼虫の解剖:
- バイアルの側面周囲にクロールされており、蛹になるために開始されていない放浪三齢幼虫を選択してください。
- 実体顕微鏡下で解剖の表面に幼虫を置きます。離れてキムワイプを使用して余分な湿り気を放出する。
- 背側が上を向くようにそれぞれの幼虫を手配する。あなたは、幼虫の背側に沿って2つの気管、白い線を見ることができます。これは背側正中線をマークするため、可能な限り中央にあるセンターこれらをしてみてください。
- ちょうど口のフックに場所のピン#1(図1)。
- #2はちょうど気管との間の気門の上に尾部にピンを置きます。
- キューティクルの乾燥を防ぐために幼虫に少量のPBSを追加。
- 遺伝子型は各遺伝子型間にピンを配置することにより、空間的に離れていることを確認しながら、すべての幼虫で、上記の手順を繰り返します。
- 右のピンの上に小さな横カットの両方気管間#2(図1)してください。
- 過去のピンまでの気管の間に背側正中線に沿って最初のカットとカットのすべての方法#1であなたのはさみを挿入します。
- あなたは左と右サイドは、キューティクルを引っ張ることなく、それらを突き止めることができるように開放されるように、キューティクルの各側面の非常に上部と下部に他の小さな切り傷を作る必要があるでしょう。
- 場所ピン#の3、4、5、および各コーナーで6。
- 室温で20-25分のために4%PFAでサンプルを修正
- PFAをデカントし、毎回デカント、PBSで3〜4回すすいでください。
3。組織の除去:
- 通常、脳が配置されている領域を保護するため後端で組織を除去することによって開始します。
- steromicroscopeとPBSのドロップの下で幼虫とを使用している間、最初に気管の一つの後端をつかみ、それと脂肪体の一部を削除可能性がある、それを引き出します。第二気管について、この手順を繰り返します。
- 後部脂肪体や腸の一部を取得し、準備から外せない。後部組織のほとんどが削除されると、脳の周囲の小さな組織を取り除くことに焦点を当てる。あなたは筋肉、またはNMJの形態にのみ興味があればあなたも脳を削除することができます。
- 脳の除去は、腹部のセグメントの方向に役立つマーカーである筋肉月31日のよりよい視覚化することができます。
4。アクティブなゾーンのための免疫蛍光:
- 解剖皿の解剖プレップにブロッキング剤(PBS中の5%正常ヤギ血清)を追加します。室温で1時間ブロックします。あなたが唯一のブロッキングと抗体希釈用つのソリューションを可能にすることを希望する場合、0.1%TritonX - 100組織の透過性については、ブロッキング剤に含まれています。
- 0.1%TritonX - 100でのブロッキング試薬で希釈した一次抗体を適用します。アクティブなゾーンの抗体は、発達研究ハイブリドーマバンク(DSHB、http://dshb.biology.uiowa.edu/)によって達成可能である。 NMJ膜と軸索にラベルを付ける(例えば、西洋ワサビペルオキシダーゼ- Cy3の共役)参照抗体を使用してください。
- 4℃で一晩一次抗体をインキュベートする。冷蔵庫の乾燥度は抗体溶液を蒸発しないように、使用されるピペットチップボックスの一番下にある水を入れて湿度のチャンバーを作成します。漂白蛍光を減らすためにアルミホイルで半透明のふたをカバー。湿度チャンバ内の解剖のスライドを置き、一晩4℃インキュベーターで全体のボックスを置く。
- 翌朝、PBST(0.2%トリトン- X - 100を含むPBS)で5〜10分ごとに5〜10倍で洗ってください。
- 対象となる湿度のチャンバー内で2〜4時間室温で0.1%TritonX - 100でのブロッキング試薬で希釈した二次抗体(例えばインビトロジェンAlexaのヤギ抗マウス- 488)でインキュベートする。あなたが筋肉や脳内の核の位置を特定したい場合は、二次的に(例えば、DAPI)染色核が含まれています。
- PBST(0.02%のTriton - X - 100を含むPBS)で5-10分ごとに5-10回洗浄する。
5。スライド上の幼虫のサンプルをマウントする:
- 最後のPBSTで洗浄をデカントし、オフウィク。
- ピンダウン幼虫の毛皮をカバーするためにピペットで約1mLの80%グリセロール。
- すべての幼虫の6ピンの5を取り外します。
- あなたは、スライドの最初の遺伝子型を置くことgenotyのすべての幼虫の最後のピンを削除する準備ができたら、PE。
- 洗浄したスライド上にマウントするメディアの2滴を入れて"洗浄"のスライドを準備。このステップでは、マウントと明確なイメージングのための幼虫オフ水溶液を洗浄することが重要です。
- あなたの鈍ピンセットで尾して、1つの幼虫をつかむ、グリセロールの大部分を削除するには、周りのグリセロールにして、解剖皿の乾燥したエッジに沿って幼虫をドラッグします。第一封入剤のドロップで幼虫を置きます。
- 同じ遺伝子型の幼虫のすべてについて、ステップ6を繰り返します。
- 一度、すべての幼虫は、最初の封入剤のドロップで、尾による最初の幼虫を取得し、媒体の大部分を除去するために皿の乾燥したエッジに沿って、ドロップとを介してプルです。その後、2つめの固定培地のドロップに幼虫を置く。
- 同じ遺伝子型を持つすべての幼虫のための手順8を繰り返します。
- それをラベリングし、スライド上にマウントするメディアの小さな"T"の形を置くことによってあなたの最終的なスライドを準備します。
- ステップ5.8のように、尾部で幼虫をつかみ、2つめの固定培地のドロップで、その後、"ウォッシュ"スライドの乾燥したの側面に沿って毛皮を引き出します。メディアの大多数が脱落した後、封入剤の最後のスライドに幼虫の毛皮を置きます。
- すべての幼虫の毛皮のために繰り返します。
- すべての幼虫の毛皮がようにキューティクルの側がダウンしていると暴露した筋肉が上を向いていることを確認して、目的の配置されていることを確認してください。
- "T."の上のラインの開始エッジで、解剖幼虫をカバーするためにスライド上にカバースリップを置き、
- 封入剤は、カバースリップの下に完全に広がることを確認し、カバースリップ上の小さな量(例えば9ボルト電池を)置く。
- 封入剤が硬化するまで、可能な蛍光消光を減らすために、引き出しや暗い空間でスライドを配置。
- カバースリップの研がをオーバーフローマウンティング培地をクリーンアップします。 70%エタノールでカバースリップまたはスライド上の過剰な乾燥した封入剤を拭き取ります。
- 最終的なシールを作成するには、指の爪磨きで封印。
6。代表的な結果:
解剖と免疫染色幼虫の例のスライドを図2に示されています。幼虫の解剖、汚れ、および取り付けが適切に行われている場合、ユーザーはセグメントA2 - A6に関心の幼虫の筋肉(図3)を識別することができます。ホアンらは詳細な筋肉の位置の説明だけでなく、特定のシナプスboutons(ホアン、千葉、2001)の特性を提供します。ハイパワーの目標を使用してユーザーが単一のシナプスに注目し、Zスタック画像を取ることができます。シナプス6月7日の部分の代表的な最大値投影画像(図4)示されています。イメージングソフトウェアを使用し、アクティブなゾーンを定量的にあなたの突然変異の表現型に特有の特性を評価することができます。特定の突然変異の表現型に変更される可能性のある機能の例としては、総数、flourescence強度、およびアクティブなゾーン間の間隔が含まれています。
潜在的な落とし穴:
幼虫神経筋接合部の免疫蛍光の評価は、このシステムの有用性を制限することができる潜在的な落とし穴があるが、シナプス生物学に貴重な洞察を提供することができる貴重なシステムです。例えば、頻繁にそれは、同じサンプルで複数の一次抗体の染色パターン画像に望ましい。適切な一次抗体、適切な蛍光物質、および適切な陰性対照の選択は、成功に不可欠です。
あなたの実験でイメージング二つの抗原の場合、一次抗体は、目的の各蛋白質が一意に蛍光標識二次抗体で識別できるように、異なる種に由来していることを確認して。バックグラウンドを低減するための推奨方法は、二次抗体(ヤギ抗ウサギまたは抗マウス二次抗体を用いてブロッキング剤として、例えば使用する正常ヤギ血清)の派生元である種の正常血清を含むブロッキング剤を使用することです。
あなたが実験に使用する一次および二次抗体を計画した後、また、あなたの染色のコントロールを含め検討する必要がありますすることで、見ている蛍光シグナルは本物だけでなく、バックグラウンドであることを保証するために設定します。可能であれば、目的のタンパク質が含まれていないネガティブコントロールサンプルが含まれています。これは簡単に野生型の動物を含めることにより、外因性遺伝子の過剰発現の研究で行われているが、いくつかのケースでは目的タンパク質のホモ接合変異体を使用することで実現できます。別の本質的なネガティブコントロール、あなたの蛍光シグナルのバックグラウンドレベルの測定に有用な、主を含むことなく、二次抗体を使用することです。するときには、イメージングして、その結果プレップ、それをあなたが見ていることを蛍光シグナルのすべてが本物ではないかもしれないことを理解しておくことが重要である、とよからの背景かもしれないUR二次抗体または内因性の組織の背景から。あなたは、蛍光強度の間の変更をアッセイする際にバックグラウンドの蛍光を考慮して画像を正規化する必要がある場合があります。
蛍光体の選択は慎重に行う必要があります。それぞれの蛍光体と一意に各信号を検出する能力のための励起および発光スペクトルの考慮はを通じてチャンネルの出血を避けるために不可欠です。不確実性、裏写りの程度一度に1つのフルオロフォア標識抗体と免疫染色、あなたの最後の実験で使用することを各チャネルを持つイメージを確認するかどうかを判断する簡単な方法がある場合。経由でブリードがあなたの顕微鏡フィルターセットで発生している場合、これは明らかにする必要があります。

図1:キューティクルカットし、端子1-6の配置のフローチャートは、 より大きな画像を表示するにはここをクリック

図2。最後のスライド上の幼虫の配置。幼虫プレップがダウン筋肉の面を上にしてキューティクルの側面を露出していることを確認します。各サンプルの間に少なくとも幼虫の体の幅のままにしておきます。

図3。ときイメージング、関心の筋肉とセグメントを探します。一般的な神経筋接合部では、支配する筋肉の6 / 7、筋肉13、筋肉12、または筋肉4を特徴とする。腹側正中線の反対側に、示されて半セグメントの鏡像の筋肉の構造があります。あなたが撮影であるセグメントを識別するマーカーとして筋31を使用してください。定量化のために使用することができる遺伝子型ごとに多数のシナプスの画像を収集してサンプルの残りの画像と同じシナプスとセグメント、。

図4。NMJ神経支配筋6月7日の部分の代表的な最大の投影画像が表示されます。 NC82(Bruchpilot)染色は緑で表示されます。 HRPシナプス前染色は赤色で表示されます。さらに画像処理ソフトウェアによって定量することができるアクティブゾーンpunctaeに注意してください。
Subscription Required. Please recommend JoVE to your librarian.
Discussion
ニューロンは、シナプス終末の面積は非常に重要であり、そしてポストとシナプス前細胞との間の適切な通信のためのブリッジです。疾患モデルにおけるニューロンの健康を調査する強力な方法は、免疫蛍光法によりシナプス末端のタンパク質を分析することです。ここで紹介する免疫蛍光法では、グループ間の環境の違いを制限しながら、研究者は、同時に多くの幼虫を調べることができます。 ショウジョウバエ 3齢幼虫の中枢神経系ではグルタミン酸作動性シナプス、マッピングされ、繰り返されるシナプス端末、遺伝子操作のアクセシビリティ、再現性とパワーを含む多くの利点があります。特にショウジョウバエ神経疾患モデルで、アクティブゾーン蛋白質Bruchpilotのレベルが変更されています。幼虫の多数が同時にアッセイするための、提示の方法を使用してアクティブゾーン分析は、ニューロンの健康の根本的な欠陥を反映することができるグループ間の微妙な違いを検出するための研究が可能になります。
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
利害の衝突は宣言されません。
Acknowledgments
我々は、この原稿についての有益なコメントのために博士Nael Alamiと博士ナムチョルキムに感謝。
Materials
| Name | Company | Catalog Number | Comments |
| Sylgard 184 Silicone Elastomer Base | Dow Corning | 68037-59-2 | After mixing allow for bubbles to rise slowly out by putting on slow rotator or allowing to sit for 30 minutes or more. |
| Stainless Steel Minutien PIns | Fine Science Tools | 26002-10 | Trim to approx. 3-4mm in length with regular scissors |
| Laminectomy Forceps (Blunt- Used for grasping pins) | Fine Science Tools | 11223-20 | Use as blunt forceps for grasping pins |
| Dissection Forceps | World Precision Instruments, Inc. | 501985 | |
| SuperFine Vannas Scissors, 8cm long | World Precision Instruments, Inc. | 501778 | |
| Mouse anti-Brp antibody | DSHB | NC82 | Use 1:50 dilution |
| Cy3 Affinipure Goat Anti-Horseradish Peroxidase | Jackson ImmunoResearch | 123-165-021 | Use at 1:200 dilution |
| Alexa Fluor 488 Goat anti-Mouse IgG | Invitrogen | A11001 | Use approx. 1:200 dilution |
References
- Budnik, V. Synapse maturation and structural plasticity at Drosophila neuromuscular junctions. Curr Opin Neurobiol. 6, 858-867 (1996).
- Fouquet, W., Owald, D., Wichmann, C., Mertel, S., Depner, H., Dyba, M., Hallermann, S., Kittel, R. J., Eimer, S., Sigrist, S. J. Maturation of active zone assembly by Drosophila Bruchpilot. J Cell Biol. 186, 129-145 (2009).
- Hoang, B., Chiba, A. Single-Cell Analysis of Drosophila Larval Neuromuscular Synapses. Developmental Biology. 229, 55-70 (2001).
- Keshishian, H., Kim, Y. -S. Orchestrating development and function: retrograde BMP signaling in the Drosophila nervous system. Trends in Neurosciences. 27, 143-147 (2004).
- Kittel, R. J., Wichmann, C., Rasse, T. M., Fouquet, W., Schmidt, M., Schmid, A., Wagh, D. A., Pawlu, C., Kellner, R. R., Willig, K. I., Hell, S. W., Buchner, E., Heckmann, M., Sigrist, S. J. Bruchpilot Promotes Active Zone Assembly, Ca2+ Channel Clustering, and Vesicle Release. Science. 312, 1051-1054 (2006).
- Koh, Y. H., Gramates, L. S., Budnik, V. Drosophila larval neuromuscular junction: Molecular components and mechanisms underlying synaptic plasticity. Microscopy Research and Technique. 49, 14-25 (2000).
- Lloyd, T. E., Taylor, J. P. Flightless flies: Drosophila models of neuromuscular disease. Ann N Y Acad Sci. 1184, e1-e20 (2010).
- Ratnaparkhi, A., Lawless, G. M., Schweizer, F. E., Golshani, P., Jackson, G. R. A Drosophila model of ALS: human ALS-associated mutation in VAP33A suggests a dominant negative mechanism. PLoS One. 3, e2334-e2334 (2008).
- Wagh, D. A., Rasse, T. M., Asan, E., Hofbauer, A., Schwenkert, I., Dürrbeck, H., Buchner, S., Dabauvalle, M. -C., Schmidt, M., Qin, G., Wichmann, C., Kittel, R., Sigrist, S. J., Buchner, E. Bruchpilot, a Protein with Homology to ELKS/CAST, Is Required for Structural Integrity and Function of Synaptic Active Zones in Drosophila. Neuron. 49, 833-844 (2006).
- Zweier, C., Jong, E. K. de, Zweier, M., Orrico, A., Ousager, L. B., Collins, A. L., Bijlsma, E. K., Oortveld, M. A., Ekici, A. B., Reis, A., Schenck, A., Rauch, A. CNTNAP2 and NRXN1 are mutated in autosomal-recessive Pitt-Hopkins-like mental retardation and determine the level of a common synaptic protein in Drosophila. Am J Hum Genet. 85, 655-666 (2009).

{kind=link}