

Summary
我们描述了一种方法,自动细胞培养相结合的高含量的成像,可视化和量化多个细胞过程和结构,在高通量的方式。这种方法可以帮助在进一步的基因组功能注释以及确定疾病基因网络和潜在的药物靶点。
Abstract
分子网络的建设和新型药物目标识别,基因组功能注释,作为一个十分紧迫的 1-4的问题需要加以解决的重要挑战。多元互补“组学”方法的遗传危险因素和底层的许多神经退行性疾病的致病机制提供了线索,但大多数研究结果仍然需要功能验证 5 。例如,最近的基因组关联研究帕金森氏病(PD),发现了许多疾病的风险因素的新位点,但潜在的致病变种(S)或致病机制尚不清楚6, 7。由于每个关联区域可以包含多个基因,每个表型与疾病相关的基因,采用传统的细胞生物学技术,功能评价,需要太长时间。
还有一个需要了解的分子网络链接它们所造成的表型的基因突变。据预计,疾病表型已中断的多重相互作用的结果。这些网络使用传统的分子生物学方法的重建工作将耗时。此外,来自独立研究的各个组件,还原论方法,网络预测可能会低估了网络的复杂性8。这低估,部分解释由于不良或有毒副作用的药品批准文的成功率很低。获得疾病相关的途径,使用HT / HC细胞筛选方法,并确定这些途径中的关键节点网络的角度来看,可能导致确定的目标是更适合的干预治疗。
高通量筛选(HTS)是一种理想的方法来解决这些问题9-12。但传统的方法是一维全以及细胞检测,用于简化STIC读数复杂的生物过程。他们无法同时在神经退行性疾病,如轴突运输赤字或改变形态属性13,14中观察到的许多表型量化。这种方法不能用于研究细胞过程或致病事件发生在细胞亚群的动态性质。要量化一个移动到多维表型等特点,被称为高内涵筛选(HCS)4, 15-17。 HCS是几个过程的细胞为基础的量化,同时,它提供了一个更详细的各种扰动的细胞反应的代表性,以高温超导相比。
HCS已超过19高温超导18,有许多优点,但进行高吞吐量(HT)技术含量高(HC)的神经元模型的屏幕是有问题的的,由于成本高,环境变化和人为错误。为了检测细胞反应,在“phenomics”规模使用HC成像之一,以减少偏差和错误,同时提高灵敏度和重复性。
在此我们描述了一个方法,准确,可靠地使用自动细胞培养20和HC成像在神经元细胞模型进行的shRNA屏幕。我们描述了我们如何使用这种方法来确定一个特定的蛋白质,DJ1,当突变导致常染色体隐性遗传性帕金森病 21调制器。
结合HC的影像与羟色胺的方法的多功能性,它可以准确地量化过多的表型。随后可利用的基因组,以推进我们的理解,参与疾病的发病机制的途径,以及找出潜在的治疗靶点。
Protocol
1。自动贴壁细胞培养过程中(图1) 20
- 准备自动细胞培养系统,细胞培养板的输入。负载消耗品(如枪头,细胞培养板,实验板)到系统中使用的图形用户界面(GUI)。确保有足够的细胞培养基,磷酸盐缓冲液(PBS)和胰蛋白酶在机器人系统。
- 手动种子两个2 × 10 6细胞,每盘的SH - SY5Y神经母细胞瘤细 胞株,omnitray板。 10%胎牛血清(FBS),维持在OPTI - MEM细胞。放入细胞培养,使用GUI机器人的板块。将在37 ° C和5%CO 2孵育细胞。
- 选择其中细胞培养协议需要发起20。一是可以选择从22日 ,贴壁细胞培养过程中培养和扩大胚胎干细胞(ES细胞)的小鼠饲养细胞 23或悬浮培养CELLS 22。
- 选择贴壁细胞培养协议(图1),确保贴壁细胞线的具体参数文件进行调整,使合流阈值(包含细胞omnitray板面积)70%。设置总胰酶消化时间为两分钟。
- 指示准备新omnitray板的机器人,一个2 × 10 6细胞每盘播种细胞数目。
- 输入到细胞培养系统,使用自动的贴壁细胞培养协议(图1)omnitray板。该协议包括以下步骤:培养板成像,直到他们达到预先定义的汇合门槛。如果细胞没有到达合流阈值,在5个读数板从系统中删除。到达合流用户定义阈值后,细胞被洗净,胰酶消化并计数。预先确定数量的细胞被添加到新omnitray板,如果有足够数量的细胞,指定麻木ER的检测板被运到甲板上,并配发到每口井的细胞定义。检测板可直接使用作进一步处理系统集成的显微镜或输出成像。
- 指示系统,准备4%omnitray板实验板,与5000细胞,每孔总。
2。 shRNA的病毒生产到检测板电镀(所需时间:6天)
- 长出甘油菌股票包含的shRNA载体(打开生物系统公司,TRC1)卢里亚- Bertani培养基中含有100微克/毫升,氨苄青霉素(Sigma - Aldrich公司)媒体2毫升过夜。
- 按照制造商的协议(Promega公司向导马格尼西合金TFX)提取质粒。
- 产生病毒的RNAi财团高通量慢病毒生产协议(96孔板)24。慢病毒工作是比较安全的,因为病毒颗粒用于转导复制缺陷和分用于其生产的基因包装策略。然而,与慢病毒工作时,更多的生物安全程序是必要的,以尽量减少对自己和他人 25的风险。所有实验必须在MLII或BSL2安全水平实验室进行。使用漂白剂与慢病毒颗粒接触的所有塑料(移液器,塑料菜,媒体)应孵育24小时前处置。
- 确定使用的pLKO.1 GFP质粒(Sigma - Aldrich公司)的GFP阳性细胞的百分比计算的慢病毒的感染复数(MOI)。
- 板慢病毒检测板,MOI 3。
3。慢病毒转导和神经元分化的SH - SY5Y细胞(所需时间:6天)
- 细胞是检测板(见第1.7步)。装载到自动细胞培养系统中包含的shRNA慢病毒的检测板。
- 24小时后,媒体上检测板将改为OPTI - MEM含0.5%FBS和0.1微米维甲酸开始分化过程。 SH - SY5Y细胞分化允许神经炎结构的可视化和同步细胞分裂。
- 继续在5天的分化媒体的实验板的孵化。这将确保最大击倒靶基因的表达。
- 第5天,24小时添加50微米H 2 O 2的实验板的一半,刺激DJ1易位到线粒体。
- 第6天,添加Mitotracker CmxROS(Invitrogen公司)的细胞,在终浓度为每孔200 nm的37 ° 30分钟。
- 一个可以指示系统的图像板直接使用的HC图像扫描仪或板可作进一步处理系统出口。
4。自动免疫检测板(所需时间:2天)
图像质量是paramounT进行一个敏感而可靠的HCS。由于不准确的移液细胞单层的损害可能会导致图像质量差和复制的结果。为了最大限度地减少细胞层损坏,使用机器人站进行免疫。程序是类似于先前已描述26,但已定制,以提高吞吐量和降低耗材的使用。
- 修复细胞的4%多聚甲醛100μL预热至37 ° C。在室温20分钟孵育。
- 与200μL5分钟,3倍的PBS洗涤细胞。
- 孵育检测板200μL的PBS含0.1%的Triton(PBST)10分钟。
- 与200μL5分钟,3倍的PBS洗涤细胞。
- 块缓冲区在室温下为1小时(含5%FBS PBST)与200μL孵育检测板。
- 与200μL5分钟,3倍的PBS洗涤细胞。
- 孵育的FOLL由于抗体4℃过夜° C时:
- 山羊DJ1的N20(圣克鲁斯,5微克/毫升)
- 兔β-Ⅲ微管蛋白(Sigma - Aldrich公司,1微克/毫升)
- 翌日,200μL的PBS洗涤细胞5分钟,3次。
- 以下的二级抗体1小时在室温下孵育检测板:
- AlexaFluor 488驴抗山羊(Invitrogen公司,2微克/毫升)
- AlexaFluor 647羊抗兔(Invitrogen公司,2微克/毫升
- 与200μL5分钟,3倍的PBS洗涤细胞。
- 用Hoechst(Invitrogen公司; 1微克/毫升)孵育细胞10分钟。
- 200μLPBS清洗5分钟,3次细胞。
- 储存于4板° C直到他们可以成像。
5。含量高,图像采集和图像分析(所需时间:5天)
- 图片共30%以及使用20X物镜领域。可视化的FI DJ1TC过滤器设置,与TRITC的过滤器设置,β-Ⅲ微管蛋白与Cy5的过滤器设置和使用紫外线过滤器设置(图3)的原子核的线粒体。
- 使用房室分析Bioapplication(Cellomics,ThermoFisher),以确定内的线粒体Mitotracker信号的平均强度分析图像。 (图4B,F)。
- 要确定平均DJ1和线粒体之间的重叠系数,使用Cellomics Colocalisation bioapplication(Cellomics,ThermoFisher)的图像进行分析。定义如下的感兴趣区域(ROI):投资回报率 - 核(图4A,E),乙的投资回报率 - 线粒体(图4的B,F)。排除乙的投资回报率的投资回报率分析,以确保只有细胞质。定义目标区域我作为目标区域II(图4C,G)的DJ1线粒体。
- 分析图像,使用神经元分析bioapplication(Cellomics,Thermofisher)跟踪突起的平均长度从β-Ⅲ微管蛋白染色(图4D,H)的。
- 使用Opera LX自动焦读者(在Perkin - Elmer)的图像板。图片共30每口井用清水浸泡使用60X物镜领域。可视化线粒体的561 nm激光和紫外光激发的原子核。
- 使用点边缘岭(SER)纹理特征算法的图像分析。在SER岭滤波器传输形成脊状花纹的像素强度。线粒体较为零散,服务岭得分越高(图8)。
6。数据正常化和分析
- 从图像分析软件的数据导入BioConductor CellHTS2 R软件环境的包(版本2.11.1,2.6版BioConductor)。
- 对数的底(2)转换数据之前,每盘的中位数为基础正常化27,28 。不适用方差调整每盘。
- 要确定一个表型的修饰符,使用differe之间的双向方差NT治疗组,即炒感染未经处理的细胞与炒毒素处理的细胞与靶基因的未经处理的细胞与靶基因处理的细胞(图5-7)。
7。代表性的成果
内DJ1突变引起发病早期隐性帕金森病21,但目前还不清楚如何DJ1损失引起的疾病表型。据悉,DJ1缺陷的细胞更容易受到氧化应激诱导的细胞死亡和氧化应激,从细胞质DJ1区转移到线粒体29,30 。通过构建慧聪检测监控这些表型,我们可以找出基因,调节或影响与DJ1相关的表型。这种方法可以帮助破译内可在疾病的发病机制涉及DJ1功能和的途径。
上位DJ1互动(图5)例:DJ1击倒在暴露细胞毒素在细胞活力的更大的损失结果(英美B:图像 - B)相比,炒慢病毒感染的细胞(英美答:图像)。击倒一个靶基因有一个类似的效果观察细胞与DJ1击倒(BAR - C - C的图像)。击倒的DJ1和靶基因的细胞活力明显大于损失或基因(BAR - D图像三维)的损失一个结果。这表明上位DJ1和靶基因答:之间的互动
基因调节DJ1易位(图6):当细胞暴露在毒素,DJ1从细胞质区转移到线粒体,这是由一个更高的重叠系数之间的DJ1和线粒体(英美一量化:图像A与范例英美- C影像C)。在沉默靶基因B已被的细胞,少DJ1区转移到线粒体细胞接触到的毒素。这表明,靶基因B是在参与DJ1运输到线粒体。 (BAR - B:图像B和BAR - D图像D)
击倒目标基因C野生型SH - SY5Y细胞在突起长度显着增加的结果:在神经细胞的生长(图7)有关的基因的一个例子(英美B:图像- B)的表达炒慢病毒感染的细胞相比的shRNA(英美答:图像A)。这种效果是失去了细胞毒素(BAR - C和D)。
线粒体形态(图8)有关的基因的例子:野生型的SH - SY5Y细胞与shRNA的目标在减少线粒体的SER岭分割值(图8,图像C和D)基因研发成果感染相比,炒慢病毒(图8,图像A和B)感染的细胞。
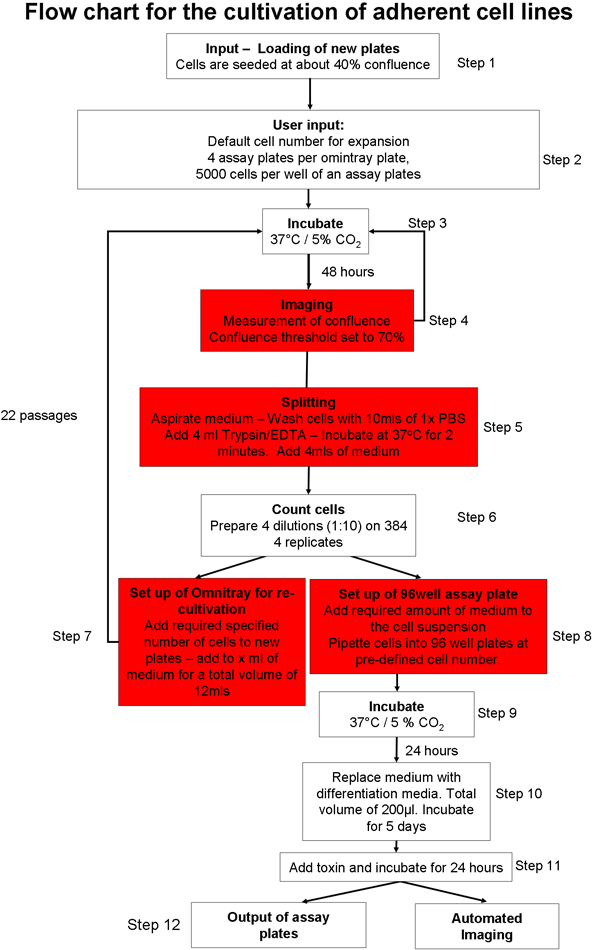
图1:自动细胞培养协议的大纲 。

图2的筛选过程中,图像分析和筛选过程中所使用的统计方法原理概述:I)细胞培养,直到他们汇合,并随后进入包含的shRNA慢病毒检测板镀。细胞是有区别的5天中和毒素,然后添加到24小时的板块。检测板从系统输出和免疫染色。括号内的数字为每个进程所采取的时间。 ii)数据使用HC成像仪(细胞活力,蛋白质易位和神经生长)和一个自动化的共焦成像仪(线粒体形态)获得。数据在R CellHTS2包出口基地(2)数转换和规范化。双因素方差分析是用来识别不同的变量之间的显著的相互作用。
图3细胞收购慧聪成像的合成图像。一)未经处理的细胞,B与H 2 O 2)细胞治疗。 DJ1标记为绿色,红色和蓝色的细胞核线粒体。突起染色不突出。

图4:从HCS几个细胞功能的定量。 AD)未经处理的SH - SY5Y细胞。 EH)的H 2 O 2处理的SH - SY5Y细胞。 A,E)核分割和投资回报率的定义;的B,F)的识别和量化的线粒体,投资回报率乙,C,G)的DJ1鉴定,第二通道目标; D,G)的鉴定和平均突起长度计算。靠近图像边缘的细胞,是从分析中排除。插入图像,图像分析前。

图6。调节DJ1易位(酒吧的字母对应图像刻字)基因的鉴定。图片A至D DJ1(绿色),线粒体(红色)和细胞核(蓝色),用于DJ1易位到线粒体定量标示的SH - SY5Y细胞。

在神经细胞生长有关的基因的鉴定(酒吧字母对应图像刻字)图7 。。 A至D SH - SY5Y细胞β-Ⅲ微管蛋白(绿色)和细胞核(蓝色),突起长度定量使用标记的图像。

图8。识别有关的基因在调节线粒体形态。图片A和C是炒的shRNA或shRNA的基因打靶D分别感染SH - SY5Y细胞的合成图像。线粒体是红色的,而核以蓝色彩色。图片B和D是可视化的服务岭量化。
Discussion
半场/ HC细胞筛选系统,降低成本,结合强大的全基因组的工具来修改基因功能的可用性,半场/ HC屏幕成为在学术界司空见惯。该方法已被成功地应用于不同的领域,如确定31-33 9日 ,在癌症和胚胎发育 34-36的药物靶标研究,甚至已经破译37,38神经精神障碍的途径申请的潜力。但是这种制度的实施,需要经常服用至少6个月的时间和努力与流程优化的显著投资。所有步骤,如胰蛋白酶消化时间,移液速度和播种密度有所调整,以确保细胞健康和成长始终。防止细菌污染是每周清洁服务协议,在C自动细胞培养面临的最困难的挑战之一ombination与不断冲洗,用70%乙醇,无污染文化所必需的所有液态轴承线。这将是还需要改进的机器人,让其他文书,如共焦显微镜更高的分辨率和-80 ° C的复合存储冰柜可以集成,
也有需要解决的限制,提高灵敏,速度和实用工具,这种方法来研究基因网络和确定有关的基因,致病性的分子途径。
要进行半场/ HC屏幕,并确保可靠的数据收集,几个方面进行优化。首先,测量的可靠性是至关重要的,是依赖时的稳定性和检测的灵敏度。例如,上面所述的检测是适合小屏幕,但难以落实,在基因组范围内,由于图像采集前的数字处理所需的步骤。因此,人们将不得不构造稳定的细胞系,表达报告基因,这将允许直接成像,导致下降的变化,由于数量减少处理步骤。目前,设计,检测,准确地描绘和可靠的量化利益的表型是在HC筛选过程的主要瓶颈。
许多屏幕在使用不同的RNAi库,所有这一切都从脱靶效应,有限的基因沉默效率和不完整的基因组覆盖率遭受哺乳动物细胞进行。因此,图书馆需要作出更具体,更有力和有更好的覆盖。目前正在努力创造这样的它是希望这些努力会改善的HT / HC SCRE重复性 EN命中。
许多大型细胞屏幕的限制,他们是在神经母细胞瘤细胞进行的,因为他们可以通过基因操纵和培养大批相对容易。然而,在体外细胞培养模型的体内功能确定的“老人与海”的相关性值得商榷,尤其是大脑的高度专业化的细胞类型,形成了突触连接功能密集和复杂的网络,作为一个高度集成的单位组成。因此,它是常见的,点击确定使用上面描述的筛选方法,在使用其他技术的二次屏幕,并在更多的生理39个相关模型验证。为了提高翻译过程中的HCS识别点击,更具代表性和更复杂的模型,如原代细胞和分化的干细胞共培养系统,需要开发和适应半场/ HC方法。
ntent“>随着自动化的细胞培养和HC成像之一相结合可以迅速获得新的见解和神经元的功能如何确定哪些途径疾病的发展是重要的,从其他”组学“方法中生成的信息相结合的HCS / HTS数据,它会然后才有可能兴建的脑部疾病的系统生物学概述,从而有利于治疗的发展。Disclosures
我们什么都没有披露
Acknowledgments
我们感谢继续支持和技术援助的伊娃Blaas汉密尔顿程序员和专家。这项工作得到了两个苏国投资补助(911-07-031和40-00506-98-10011),Prinses贝娅特丽克丝全宗Wetenschapsprijs 2009年和神经校园阿姆斯特丹,律政司司长是由TI制药支持:T5 - 207。
Materials
| Name | Company | Catalog Number | Comments |
| AI.CELLHOST | Hamilton Co | http://www.hamiltonrobotics.com/en-uk/applications/cellomics/ | |
| OPTI-MEM | Invitrogen | 31985-054 | |
| RETINOIC ACID | Sigma-Aldrich | R2625 | |
| OMNITRAY PLATES | Nalge Nunc international | 465219 | |
| 96 WELL CULTURE PLATES | Greiner Bio-One | 655086 | |
| DJ1 N20 ANTIBODY | Santa Cruz Biotechnology, Inc. | SC27004 | |
| BETA-III TUBULIN ANTIBODY | Sigma-Aldrich | T3952 | |
| MITOTRACKER CMXROS | Invitrogen | M-7512 | |
| H–CHST-33342 | Invitrogen | H1399 | |
| HYDROGEN PEROXIDE | Sigma-Aldrich | 216763-100ML | |
| TRYPSIN | Invitrogen | 25050014 | |
| DULBECCO’S PHOSPHATE BUFFERED SALINE | Invitrogen | 14190086 | |
| PROMEGA WIZARD MAGNESIL TFX | Promega Corp. | A2380 | |
| SHRNA CLONES | Open Biosystems | http://www.openbiosystems.com/RNAi/shRNALibraries/ TRCLibraryDetails/ | |
| CELLOMICS BIOAPPLICATIONS | Thermo Fisher Scientific, Inc. | http://www.thermo.com/hcs |
References
- Geschwind, D. H., Konopka, G. Neuroscience in the era of functional genomics and systems biology. Nature. 461, 908-915 (2009).
- Ge, H., Walhout, A. J., Vidal, M. Integrating 'omic' information: a bridge between genomics and systems biology. Trends. Genet. 19, 551-560 (2003).
- Zhu, H., Snyder, M. 'Omic' approaches for unraveling signaling networks. Curr. Opin. Cell. Biol. 14, 173-179 (2002).
- Jain, S., Heutink, P. From single genes to gene networks: high-throughput-high-content screening for neurological disease. Neuron. 68, 207-217 (2010).
- Manolio, T. A. Finding the missing heritability of complex diseases. Nature. 461, 747-753 (2009).
- Nalls, M. A. Imputation of sequence variants for identification of genetic risks for Parkinson's disease: a meta-analysis of genome-wide association studies. Lancet. 377, 641-649 (2011).
- Simon-Sanchez, J. Genome-wide association study confirms extant PD risk loci among the Dutch. Eur. J. Hum. Genet. 19 (6), 655-661 (2011).
- Van Regenmortel, M. H. Reductionism and complexity in molecular biology. Scientists now have the tools to unravel biological and overcome the limitations of reductionism. EMBO. Rep. 5, 1016-1020 (2004).
- Aherne, G. W., McDonald, E., Workman, P. Finding the needle in the haystack: why high-throughput screening is good for your health. Breast. Cancer. Res. 4, 148-154 (2002).
- An, W. F., Tolliday, N. J. Introduction: cell-based assays for high-throughput screening. Methods. Mol. Biol. 486, 1-12 (2009).
- Mayr, L. M., Bojanic, D. Novel trends in high-throughput screening. Curr. Opin. Pharmacol. 9, 580-588 (2009).
- Hertzberg, R. P., Pope, A. J. High-throughput screening: new technology for the 21st century. Curr. Opin. Chem. Biol. 4, 445-451 (2000).
- Conrad, C., Gerlich, D. W. Automated microscopy for high-content RNAi screening. J. Cell. Biol. 188, 453-461 (2010).
- Thomas, N. High-content screening: a decade of evolution. J. Biomol. Screen. 15, 1-9 (2010).
- Arrasate, M., Finkbeiner, S. Automated microscope system for determining factors that predict neuronal fate. Proc. Natl. Acad. Sci. U. S. A. 102, 3840-3845 (2005).
- Dragunow, M. High-content analysis in neuroscience. Nat. Rev. Neurosci. 9, 779-788 (2008).
- Varma, H., Lo, D. C., Stockwell, B. R. High throughput screening for neurodegeneration and complex disease phenotypes. Comb. Chem. High. Throughput. Screen. 11, 238-248 (2008).
- Durr, O. Robust hit identification by quality assurance and multivariate data analysis of a high-content, cell-based assay. J. Biomol. Screen. 12, 1042-1049 (2007).
- Miller, J. Quantitative relationships between huntingtin levels, polyglutamine length, inclusion body formation, and neuronal death provide novel insight into huntington's disease molecular pathogenesis. J. Neurosci. 30, 10541-10550 (2010).
- Jain, S., Sondervan, D., Rizzu, P., Bochdanovits, Z., Caminada, D., Heutink, P. The Complete Automation of Cell Culture. Journal of Biomolecular Screening. 16 (8), (2011).
- Bonifati, V. Mutations in the DJ-1 gene associated with autosomal recessive early-onset parkinsonism. Science. 299, 256-259 (2003).
- Ricardo, R., Phelan, K. Trypsinizing and Subculturing Mammalian Cells. J. Vis. Exp. (16), e755-e755 (2008).
- Kent, L. Culture and Maintenance of Human Embryonic Stem Cells. J. Vis. Exp. (34), e1427-e1427 (2009).
- Moffat, J. A lentiviral RNAi library for human and mouse genes applied to an arrayed viral high-content screen. Cell. 124, 1283-1298 (2006).
- Volksgezondheid, M. V. Integrale versie van de Regeling genetisch gemodificeerde organismen en het Besluit genetische gemodificeerde. Organismen. , (2004).
- Anderl, J. L., Redpath, S., Ball, A. J. A Neuronal and Astrocyte Co-Culture Assay for High Content Analysis of Neurotoxicity. J. Vis. Exp. (27), e1173-e1173 (2009).
- Wiles, A. M., Ravi, D., Bhavani, S., Bishop, A. J. An analysis of normalization methods for Drosophila RNAi genomic screens and development of a robust validation scheme. J. Biomol. Screen. 13, 777-784 (2008).
- Canet-Aviles, R. M. The Parkinson's disease protein DJ-1 is neuroprotective due to cysteine-sulfinic acid-driven mitochondrial localization. Proc. Natl. Acad. Sci. U. S. A. 101, 9103-9108 (2004).
- Blackinton, J. Formation of a stabilized cysteine sulfinic acid is critical for the mitochondrial function of the parkinsonism protein DJ-1. J. Biol. Chem. 284, 6476-6485 (2009).
- Gupta, P. B. Identification of selective inhibitors of cancer stem cells by high-throughput screening. Cell. 138, 645-659 (2009).
- Luo, J. A genome-wide RNAi screen identifies multiple synthetic lethal interactions with the Ras oncogene. Cell. 137, 835-848 (2009).
- Mouchet, E. H., Simpson, P. B. High-content assays in oncology drug discovery: opportunities and challenges. IDrugs. 11, 422-427 (2008).
- Vogt, A. Automated image-based phenotypic analysis in zebrafish embryos. Dev. Dyn. 238, 656-663 (2009).
- Pardo-Martin, C. High-throughput in vivo vertebrate screening. Nat. Methods. 7, 634-636 (2010).
- Vogt, A., Codore, H., Day, B. W., Hukriede, N. A., Tsang, M. Development of automated imaging and analysis for zebrafish chemical screens. J. Vis. Exp. (40), e1900-e1900 (2010).
- Ross, P. J., Ellis, J. Modeling complex neuropsychiatric disease with induced pluripotent stem cells. F1000. Biol. Rep. 2, 84-84 (2010).
- Ebert, A. D., Svendsen, C. N. Human stem cells and drug screening: opportunities and challenges. Nat. Rev. Drug. Discov. 9, 367-372 (2010).
- An, W. F., Tolliday, N. Cell-based assays for high-throughput screening. Mol. Biotechnol. 45, 180-186 (2010).
