We are interested in understanding the role specific genes on metabolism, particularly respiration rate and lipid metabolism. Therefore, we treated wild type zebrafish embryos from 26 hpf onwards with a specific pharmacological inhibitor of enzyme encoded by one of these genes. At 50 hpf, half of the vehicle and inhibitor-treated embryos were analysed using the Seahorse for mitochondrial function, while the remaining half were fixed in 4% PFA overnight at 4 °C and subsequently stained for lipid deposition with Oil-Red-O. Treatment with the specific enzyme inhibitor led a ~2 fold increase in basal respiration (Figure 4A), which was associated with an increase in lipid deposition especially in the telencephalon and underneath the otic vesicle (Figure 4B,C).

Figure 1. Schematic representation of chemical treatment of zebrafish embryos. Wildtype, genetically-manipulated, or pharmacologically treated zebrafish embryos are background.
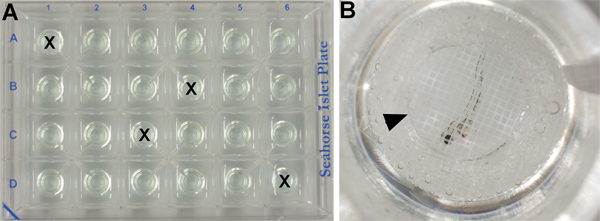
Figure 2. XF24 islet plate set up. (A) Four wells contain embryonic medium only as temperature control (marked by an X) and the others contain one embryo/well. (B) Close-up of one well (C6) showing a 50 hpf wild type embryo treated with vehicle (DMSO) covered by an islet capture screen (arrowhead).
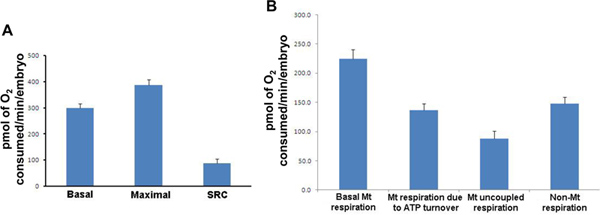
Figure 3. Respiration parameters for wild type 50 hpf zebrafish embryos. (A) For each measurement, the mean of the 20 individual embryos is presented. Basal respiration (Mean: 299.7; SD: 75.7; SEM: 16.9), Maximal respiration (Mean: 387.4; SD: 93.3; SEM: 20.9), Spare Respiratory Capacity (Mean: 87.7; SD: 72.0; SEM: 16.1). (B) For each measurement, the mean of the 20 individual results is presented. Basal mitochondrial respiration; mitochondrial respiration due to ATP turnover; mitochondrial uncoupled respiration and non-mitochondrial respiration. Mt: mitochondrial; SRC: Spare Respiratory Capacity; SD: Standard Deviation, SEM: Standard Error of Mean. Results are presented in pmol of O2 consumed/min/embryo. Click here to view larger figure.
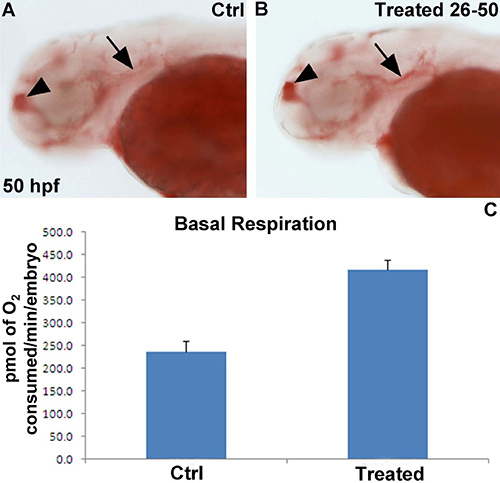
Figure 4. Representative results of chemical exposure on basal respiration and lipid deposition. (A,B) 50 hpf embryo stained with Oil-Red O in lateral view. The embryo incubated with the selective inhibitor (B) displays more Oil-Red-O staining in the telencephalon (arrowhead) and underneath the otic vesicle (arrow) compared to a vehicle-treated control embryo. (C) Basal respiration shows an ~ 2 fold increase in chemically treated embryos (n=10 embryos per group). Results are presented in pmol of O2/min/embryo.
