

Summary
Мы представляем Экс Vivo клеточную миграцию анализа, который позволяет точное количественное определение кишечной нервной миграции гребень клеток потенциала в присутствии различных факторов роста.
Abstract
Клетки нервного гребня (NCC) являются временными и мультипотентны клеточная популяция, которая происходит от дорсальной части нервной трубки и мигрирует широко во всех развивающихся эмбрионов позвоночных. В дополнение к предоставлению периферийные глии и нейронов, NCC генерировать меланоциты, а также большую часть черепно-лицевого скелета. NCC миграция и дифференциация контролируется сочетанием их осевой происхождения вдоль нервной трубки и степень их подверженности региональном различных внеклеточных сигналов. Такой вклад внеклеточных лигандов особенно очевидно во время формирования кишечной нервной системы (ENS), сложной взаимосвязанной сети нейронной ганглиев, что локально управляет (среди прочего) движения кишечника мышц и моторики кишечника. Большая часть ENS происходит от малого начального пула NCC, что предпринять долгое путешествие, чтобы колонизировать - в ростральнее хвостового моды - по всей длине предполагаемого кишечнике. Среди нескольких сигнальных путей, известныхвлиять кишечно NCC колонизации, GDNF / сигнализации РЭТ признан самым важным. Действительно, пространственно-временной контролируется секреция РЭТ лиганда GDNF в кишечнике мезенхимы является главным ответственным за привлечение и руководством RET-выражения кишечной NCC к и в эмбриональной кишки. Здесь мы описываем экс виво анализа миграции клеток, что делает использование трансгенной линии мышей, обладающего флуоресцентно меченого NCC, которая позволяет точную количественную оценку кишечной NCC миграционного потенциала в присутствии различных факторов роста, в том числе GDNF.
Introduction
Клетки нервного гребня (NCC) являются переходным типом клеток уникальным для позвоночных, которая формирует множество производных во время развития эмбриона. Это популяция клеток возникает на границе нервной пластинки, рядом с не-нейронных эктодермы 1. Во время нейруляции, изгиб нервной пластинки мест NCC вдоль верхнего края формирующегося нервной трубки. Затем НКК пройти переход эпителиально-мезенхимальных, разделения и миграции от нервной трубки. NCC колонизируют различные эмбриональные структуры, в том числе желудочно-кишечного тракта, где они образуют весь кишечно нервную систему (ENS), взаимосвязанной сети нейронной ганглиев, встроенного в стенку кишечника. Совсем недавно, отзывы 2,3, многие гены были вовлечены в разработку этой сложной структуры.
Большую часть ENS происходит от небольшой пул NCC происходящих из блуждающего нервной трубки (т.е. вокруг предполагаемой границы заднего мозга / спинного мозга) 4.Эти нервные клетки-предшественники достичь передней кишки вокруг эмбриональный день (е) 9,0 у мышей, а затем мигрируют каудально в кишечнике мезенхимы до примерно e15.0 не колонизировать весь эмбриональных кишечник. Несовершеннолетний подмножество толстой нейронных клеток-предшественников также обеспечивается сакральной NCC, который вторгнуться заднюю кишку в противоположном направлении до слепой кишки 4. Оба блуждающего и крестцового NCC требуют несколько миграция-, нераспространения, выживание-и дифференциация способствующих сигналы, чтобы обеспечить полное формирование ENS. В связи с этим, модели животных - особенно генетически модифицированные мыши - играют важную роль в выявлении нескольких существенных внеклеточных лигандов: GDNF (глиальных клеток нейротрофический фактор), эндотелина-3, нейротрофин-3, БМП (костные морфогенетические белки), Netrin , а также Соник и индийская Ежик (Тсс и Ihh) 5-10. Из них GDNF сигнализации через трансмембранный рецептор RET тирозинкиназы (переставить во трансфекции) признан йэ наиболее важным путем для привлечения и руководством НКК в и в эмбриональной кишки. GDNF секретируется кишечника мезенхимы и образует пространственно-временной контролируемый rosrrocaudal градиент, непосредственно chemoattractive к кишечной NCC, которые выражают РЭТ 11,12.
Среди других функций, ЭНС регулирует движение в желудочно-кишечном тракте через его взаимодействие с гладкой мускулатуры в стенке кишечника. Отсутствие нервной ганглиев в концевой области результатов кишечника при болезни Гиршпрунга: тоническое сокращение пораженного сегмента приводит к закупорке, вверх по течению накопления сброженного материала и массивной живота кишки и брюшной полости. Болезнь Гиршпрунга происходит примерно один из 5000 живорожденных. Ростро-каудальном миграция картина кишечной NCC, как полагают, является основным фактором, способствующим этиологии болезни Гиршпрунга. Толстой кишки, удалена от источника миграции NCC и последнюю часть бOwel подвергшихся колонизации, является наиболее восприимчивы к дефектам в формировании ENS. В соответствии с его решающую роль в кишечной миграции NCC, нарушение GDNF / RET сигнализации является основным известным генетическая причина болезни Гиршпрунга 13.
Чтобы лучше изучить NCC и развитие ENS, мы получили трансгенной линии мыши - по имени Gata4p [5kb]-GFP 14 - в которых миграционный NCC помечены зеленого флуоресцентного белка (GFP). Мы рядом усовершенствовал Экс Vivo миграции клеток анализа, адаптированный из опубликованных работ других групп 11,12,15, что теперь позволяет точное количественное определение кишечной NCC миграционного потенциала в присутствии различных факторов роста, таких как GDNF.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
Заявление Этика
Опыты с мышами проводились следующие Канадского совета руководящих принципов по уходу за животными для ухода и манипуляции животных, используемых в медицинских исследованиях. Протоколы с участием манипуляции животных были утверждены комитетом институциональные этики Университета Квебека в Монреале (Комитет Institutionnel де Защита де Animaux; справочный номер 0512-R3-650-0513).
1. Подготовка коллагена Гели
Работа в стерильных моды, под капот культуры ткани.
- Подготовить всю 5x DMEM (модифицированной, по существу среда Игла), включая стандартные антибиотики. Растворите 3,37 г DMEM порошка и 0,925 г NaHCO 3 в 20 мл воды. Стерилизацию при прохождении через фильтр 0,22 мкм. Добавить 2,5 мл стерильного 100x пенициллина / стрептомицина и 25 мл стерильной инактивированной нагреванием фетальной бычьей сыворотки. Хранить при 4 ° С.
- На льду, смешать 800 мклраствора коллагена I (3,77 мг / мл в 0,02 N уксусной кислоты, фильтр стерилизуют), 600 мкл полной среды DMEM 5-кратным и 17 мкл 1 N NaOH. Развести стерильной водой до конечного объема 3 мл. Включить соответствующие факторы роста в миксе. Обычно мы используем GDNF при 10 нг / мл стимулировать энтеросолюбильного миграции NCC.
- Депозит около 480 мкл в каждую лунку одной строки в 24-луночный планшет. Избегайте пузырей. Остальные строки могут быть использованы для оценки влияния других факторов роста на клеточной миграции.
- Пусть коллаген полимеризации по крайней мере 1 час в стерильном инкубаторе при температуре 37 ° С.
2. Вскрытие животных 16
- Настройка вязки и проверьте вагинальных пробок на следующее утро. День E0.5 быть полдень в день влагалища вилка найти, изолировать самку и не ждать 12 дней, пока E12.5.
- Обезболить беременных мышей изофлураном и усыпить по CO 2 ингаляции.
- Спрей мышь с 70% этанола. Поднимите живота секкин и открыть живот полость с рассекающих ножницами.
- Удаление матки в стеклянную чашку Петри, наполненную ледяной PBS (забуференный фосфатом физиологический раствор). Вырезать матку поперечно между отдельными вздутий deciduum изолировать каждый имплантация эмбриона сайт.
- Работа на каждом участке имплантации отдельно в другой стекл нную чашку Петри, наполненную ледяной PBS. При вскрытии микроскопом, использовать тонкий пинцет для удаления мышечного слоя матки.
- Откройте висцерального желточного мешка и амнион выявить эмбрион. Будьте осторожны при разрыве кровеносных сосудов присоединения эмбриона к плаценты / висцерального желточного мешка, так как они переплетаются с развивающимися кишечника.
- Sever головой эмбриона на шее.
- Включить закрытые щипцы в брюшной полости эмбриона, чуть выше темно-красного цвета печени и пусть щипцы открыты себя (остановка приложения давления), чтобы сделать поперечную отверстие в брюшной полости рта. Потяните открытые щипцы вниз кЗадний конец эмбриона, чтобы открыть живот полностью.
- Захватите соединительную ткань позади печени и тянуть кишки из живота, стараясь не сломать кишечник (двоеточие прилагается к анусу).
- Разрежьте двоеточие, чтобы освободить кишки от остальной части эмбриона. Разрез можно сделать в любом месте вдоль толстой кишки. Забронируйте хвостовой части на потом.
- Дразнить соединительную ткань от слепой кишки, затем остальные кишечнике. Будьте осторожны, чтобы не ранить кишечник, делая так.
- Срежьте печень и желудок (на ростральной конце тонкой кишки), а также мезонефроса и генитальные гребни, если некоторые из них присутствовать.
- Изолировать в тонкий кишечник. Опять же, будьте осторожны, не пресечь кишечник. Отныне rosrrocaudal ориентация ткани кишечника можно отслеживать с помощью резкого кривизну присутствующих на ростральной конце. Оставьте тонкую кишку в PBS при комнатной температуре в течение такого короткого периода, как это возможно, прежде чем EMпостельные принадлежности (см. шаг 3.3).
- Наконец, запишите число хвостовых сомитах для каждого эмбриона точно определить стадию эмбрионального развития.
3. Секционирование эмбриональных кишечнике
- Прежде чем приступить к эмбриона вскрытий, растопить 1,5 г агарозы в 100 мл PBS (фосфатно-солевой буфер) и держать при температуре 50 ° С.
- Налейте расплавленную агарозу в включени пресс-формы (например, закрыты 2 трубки микроцентрифужных мл, разрезанные в продольном направлении, чтобы вырезать приблизительно 1/4 от стенки трубы). Пусть агарозы остыть примерно до 42-45 ° С, она должна быть лишь слегка теплой на ощупь.
- Вставить эмбрионального кишечника как раз перед агарозном затвердевает (это могут быть оценены с щипцы советы и произойдет вокруг 36-38 ° С). Проведение кишечника с помощью щипцов сложенном ростральной конце концов, вытянуть его очень медленно через агарозы по длине формы. Это помогает держать прямой кишки а множества агарозы. Отпустите твыдавать, как только он начинает сопротивляться перемещается. Следите за ростральной-каудальной ориентации кишечнике.
- Поставьте форму в холодильник 2-3 минут, чтобы обеспечить агарозном полностью установлен.
- Выньте агарозы из формы (по сдвинув ее из открытой трубки Эппендорф). С лезвием, вынуть лишний агарозы на обоих концах, делая сокращения, перпендикулярные кишечника.
- Приклейте ростральной конец кишки / агарозы блокировать вниз на сцене металла микротоме с вибрирующей лезвия. Обрежьте излишки агарозы по бокам кишечника / агарозном блока.
- Установите этап металла на вибрирующую микротома камеры. При необходимости отрегулировать угол сцены так кишечник как можно более вертикальным (отсюда перпендикулярно к лезвию). Обложка образец с ледяным PBS. Принесите микротоме лезвие до нескольких миллиметров под поверхностью буфера.
- Сделать 200 мкм Vibratome поперечные куски хвостового-самый маленький кишечника, обеспечивая, что еACH агарозном ломтик содержит полный кишечного сечение.
4. Культура кишечных эксплантов
- Аккуратно депозит свежесрезанные ломтики кишки / агарозные плоский удар коллагеновых гелей с пинцетом, помещая одну часть к середине каждую лунку.
- Выдержите 3 дня при 37 ° С, в условиях повышенной влажности 5% СО 2 атмосферы, чтобы позволить миграцию NCC из эксплантата.
- Возьмите ломтики кишки / агарозные от коллагеновый гель очень осторожно пинцетом. Позаботьтесь, чтобы не повредить гель ниже.
- Старайтесь не прикасаться к коллагена гель непосредственно во время последующих инкубации и промывки шагов в целях обеспечения того, картина миграции клеток не нарушается. Исправить с 500 мкл 4% PFA (параформальдегид) (в PBS) на лунку 1 ч при комнатной температуре.
- Заменить фиксатор 500 мкл DAPI (4 ',6-диамидино-2-фенилиндол) Раствор (5 мкг / мл в PBS) на лунку и инкубировали 10 мин при комнатной температуре.
- Вымойте каждую лунку 3x500 мкл PBS в течение 5 мин.
- Сфотографируйте флуоресцентные клетки (GFP и DAPI каналов), встроенные в коллагеновый гель в каждую лунку.
5. Анализ изображений
Мы широко использовали ImageJ 17 для обработки и количественной оценки изображения, созданные после эксплантата культуры.
- Начните с визуализации микрометра слайд при том же увеличении, как на фотографиях люминесцентных клеток. Измерьте длину одного микрона в пикселях (с использованием линейного инструмент Линия).
- Установите масштаб (Анализ / Задать шкалу; вход число пикселей / микрона).
- Для каждого флуоресценции GFP фотографии, изменить формат изображения в 8-битный оттенках серого (Image/Type/8-bit).
- Отрегулируйте интенсивность сигнала (изображения / Настройка / Яркость / Контрастность).
- Вычтите фоновый шум при необходимости (процесс / вычитание фона; настроить радиус катящийся шар).
- Установить порог, чтобы выделить клеток и клеточных скоплений (Изображение / Настройка / Threshold),н применить водораздел разделить сгустки (процесс / Binary / водораздела).
- Анализ частиц указать интересующие участки (ROI) и генерировать статистику количества клеток (Анализ / Анализ частиц, набор минимальный размер, чтобы исключить оставшиеся пикселей).
- Установите параметры измерения включить диаметр Фере (Analyze / Set измерений).
- Группа трансформирования и измерить в целом, чтобы определить диаметр в Фере, указание распространения клеток (Анализ / Инструменты / Диспетчер ROI / Подробнее / ИЛИ, то менеджер ROI / Добавить и менеджер ROI / Measure).
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
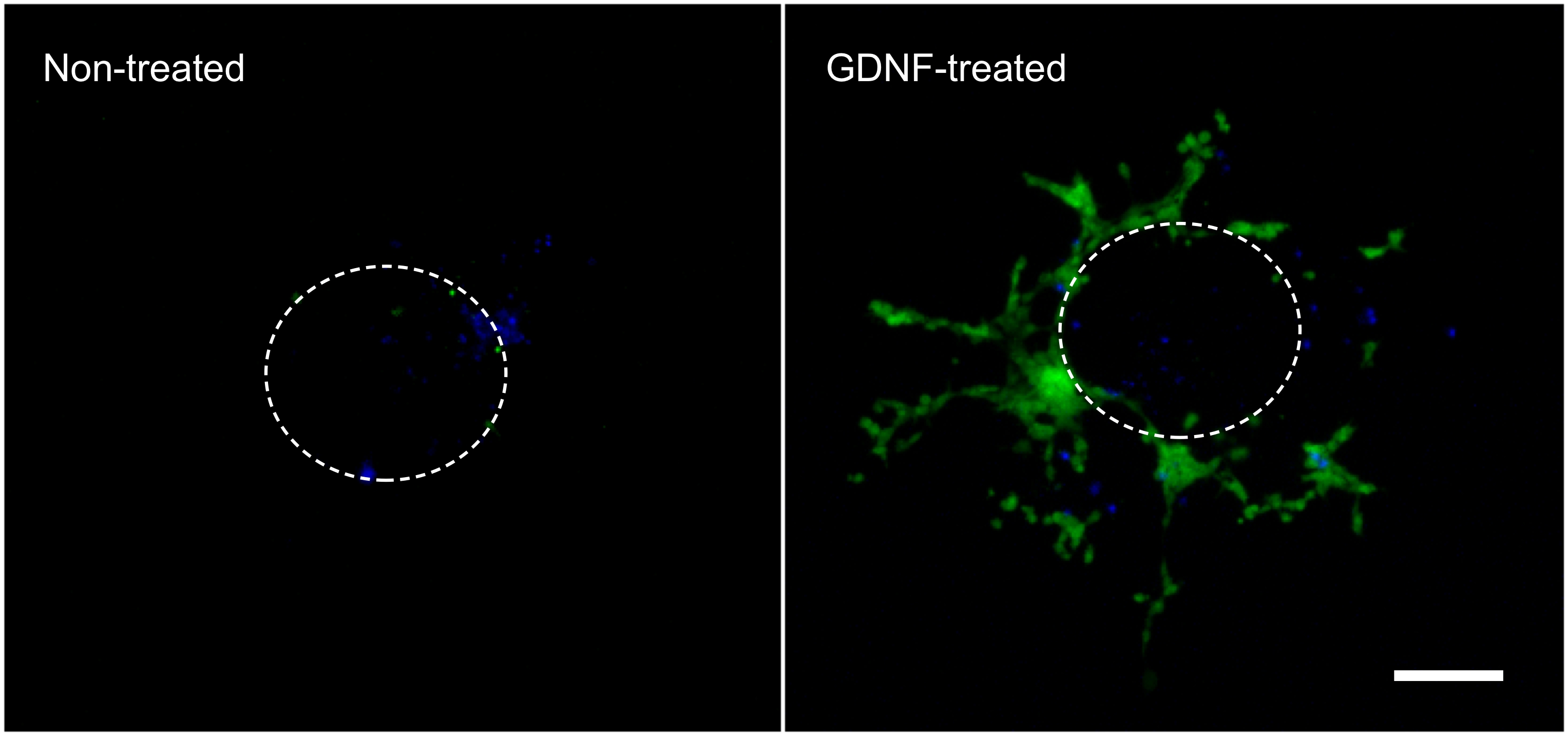
Следующие результаты являются репрезентативными что могут быть получены с методикой, описанной здесь (рис. 1). Использование факторов роста (т.е. GDNF) стимулирует миграцию GFP-экспрессирующих кишечно NCC из кишечника эксплантата и в гель коллагена (рис. 2). Хотя некоторые клетки выходят из эксплантата в отсутствие факторов роста, в основном это не GFP-меченых и представляют пассивную запись. Надо удалить кишечную кусочек от коллагеновый гель для записи результатов, как эта ткань по-прежнему сильно населена флуоресцирующих клеток и в противном случае скрыть клетки, лежащие в коллагена под ним. Количественная оценка этих результатов показывает, что многие другие клетки находятся в коллагеновый гель, когда GDNF присутствует, и что активная миграция имеет место (рис. 3). Действительно, пассивные клетки находятся непосредственно под эксплантов, в пределах диаметра кишечной ломтик, порога EAS клетки, которые активно вторгаются коллагена гель отойти от их места происхождения и распространяться дальше.

Рисунок 1. Обзор техники эксплант культуры. A) Равномерное населения флуоресцентного кишечной NCC в хвостовой тонкой кишки регионе используется, чтобы сделать 200 μ-толстые эксплантов. Шкала бар. 200 мкм B) культуральную среду, содержащую коллаген осаждается в 24-луночных планшетах и оставили твердеть в течение 1 часа. Vibratome ломтики геле встраиваемый эмбриональных кишечника оседают на гелей (один ломтик на лунку), и люминесцентные кишечно NCC разрешается мигрировать из эксплантов в течение 3 дней. Ломтики затем вылетел до визуализации ячейки, которые вторглись в гели коллагена.s/ftp_upload/50709/50709fig1highres.jpg "целевых =" _blank "> Нажмите здесь, чтобы посмотреть увеличенное изображение.

Рисунок 2. Миграция клеток из кишечника эксплантов и в коллагеновый гель. Клеток, которые мигрировали из кишечной эксплантов в течение 3 дней инкубации в отсутствии или в присутствии 10 нг / мл GDNF фиксировали, окрашивали с DAPI (синий) и фотографировали в показать GFP-меченых кишечно NCC (зеленый). 70X увеличением. Шкала бар: 100 мкм. Пунктирная линия представляет приблизительный размер и расположение эксплантата прежде чем она была снята с гелем. Нажмите здесь, чтобы увеличить рисунок. Нажмите чпрежде чем, чтобы увеличить изображение.

Рисунок 3. И распространение (диаметр Фере) из GFP-экспрессирующих клеток, мигрирующих из кишечника эксплантатах через 3 дня Количественная оценка кишечной NCC миграционного потенциала. Число количественно с помощью ImageJ программное обеспечение 17. В обоих случаях существует значительная разница между необработанными и GDNF-обработанных условиях в соответствии с т теста Стьюдента (р <0,001; *). Средний диаметр кишечных ломтиками (пунктирная линия черный: 260 мкм) был включен различать пассивной и активной записи миграции. НТ: необработанные, п:. Количество эксплантатах обработанных Нажмите здесь, чтобы увеличить изображение .
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Мы покажем, как наш бывший естественных техника культура эксплант можно использовать для точного количественного кишечно NCC миграционный потенциал в присутствии GDNF. Такое точное количественное значительно облегчается с помощью 200 мкм-толстые секции Vibratome кишки вместо большие куски размером приблизительно, как было описано ранее 11,12,15. В самом деле, это позволяет нам работать с разумным количеством клеток в высокой воспроизводимостью обстановке. Следует отметить, что равномерное распределение флуоресцентной кишечной NCC в хвостовом-наиболее области тонкой кишки, из которой кусочки нарезают эксплантов также позволяет анализировать несколько секций из одного кишки (рис. 1А). Кроме того, учитывая, что и кишечно-NCC и аксоны могут выйти из ткани в таких анализах 11, вывод эксплантов в конце периода культивирования позволяет нам сосредоточиться исключительно на миграционный NCC.
Большинство важных шагов были изложены в тексте протокола, однако, как шelfare кишечного эксплантата имеет первостепенное значение для получения здорового мигрирующих NCC, особое внимание должны быть приняты. Не подвергайте эмбриональных тканей к резким изменениям температуры, особенно когда кишечник вкладывается в агарозы (шаг 3.3). Убедитесь, что кишечник при комнатной температуре и агарозы так круто, как возможно (но все еще таял), чтобы избежать "приготовление" ткани. Кишечная эксплант должны процветать на питательной среды заполненные коллагеновый гель, часто увеличиваются в размерах и, разлив из агарозы среза. Если эксплантат появляется нездоровый или хуже, как правило, погибают во время инкубации, попробуйте заменить PBS с культуральной среде при комнатной температуре (например, HEPES буфером М2 или DMEM с добавлением 10% FBS), чтобы помочь поддерживать его во время вскрытия.
Главным ограничением для нашего подхода в том, что она опирается на наличие линии мыши, придающего флуоресцентную метку, чтобы миграции NCC. В отсутствие такого ресурса, антитело против мigrating NCC (например анти-Ret или анти-Sox10) может быть использован для обозначения клеток, которые вторглись в геля коллагена. Кроме того, учитывая, что кишка микросреда является гораздо более сложным, чем простой коллагеновый гель, результаты, полученные с этого анализа в пробирке не может полностью отражают поведение кишечной NCC в естественных условиях. Дополнительные эксперименты с участием изображений живых клеток рекомендуется оценивать это поведение. Следует также отметить, что в дополнение к своей роли в качестве хемоаттрактанта, GDNF, как известно, способствуют распространению миграции кишечно NCC 4. Наша мера кишечной NCC миграционного потенциала в присутствии GDNF, таким образом, вероятно смесь истинной клеточной миграции и пролиферации клеток, сродни механизмов в естественных условиях, приводящих к NCC колонизации кишечника. Если четкое различие между этими двумя процессами желательно, добавление блокатора клеточного цикла (например AZD 5438 18) в культуральной среде может ограничить анализ на мобильный migratioн.
Эта техника может быть расширена, чтобы проверить различные другие внеклеточные лиганды, а также ингибиторы специфических сигнальных путей и любую комбинацию в нем. Другие ткани также может потенциально быть расчленены и секционные, позволяя изучение NCC миграции во многих эмбриональных структур. В сочетании с романом и / или неохарактеризованных мутантных линий мышей с возможными дефектами развития NCC, наш метод может быть применен в целях быстрого обнаружения недостатков в миграционного поведения в ответ на определенные события сигнализации.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Авторы не имеют ничего раскрывать.
Acknowledgments
Мы благодарим Дениса Флипо для обработки и анализа изображений советом, и Дэвид У. Silversides в лаборатории которого был создан Gata4p [5kb]-GFP мыши линии. Исследования в лаборатории Пилон финансируется CIHR, NSERC, FRQS и FRQNT.
Materials
| Name | Company | Catalog Number | Comments |
| DMEM powder | Wisent | 219-010-XK | |
| NaHCO3 | Bioshop | SOB999 | Biotechnology grade |
| Steriflip vacuum filtration system (0.22 micron) | EMD Millipore | SCGP00525 | |
| Penicilin-Streptomycin solution, 100x | Wisent | 450-201-EL | |
| Fetal bovine serum | Wisent | 095-150 | High quality grade |
| Collagen I | BD biosciences | 354236 | |
| NaOH | Bioshop | SHY700 | Diluted from 10 N stock then sterile-filtered |
| GDNF | Cedarlane | CLCYT305 | |
| Falcon 24-well Plate | BD biosciences | 353047 | |
| Dissecting scissors | Fisher Scientific | 089515 | |
| Glass Petri dish | VWR | 89000-306 | |
| PBS | Sigma | P5493 | Cell culture grade |
| Dissecting microscope | Leica | M125 | |
| Dumont #5 Forceps | Fine Science Tools | 11251-20 | |
| Agarose | Bioshop | AGA001 | Biotechnology grade |
| Surgical blade | Feather | 21 | |
| All Purpose Instant Krazy Glue Pen | Krazy Glue | KG824 | |
| HM 650V Vibrating-Blade Microtome | Thermo Scientific | 920110 | |
| Paraformaldehyde | Sigma-Aldrich | P6148 | |
| DAPI | Sigma-Aldrich | D9564 | |
Table of specific reagents and equipment. |
|||
References
- Bronner, M. E., Le Douarin, N. M. Development and evolution of the neural crest: An overview. Dev. Biol. 366 (1), 2-9 (2012).
- Bergeron, K. F., Silversides, D. W., Pilon, N. The developmental genetics of Hirschsprung's disease. Clin. Genet. 83 (1), 15-22 (2013).
- Obermayr, F., Hotta, R., Enomoto, H., Young, H. M. Development and developmental disorders of the enteric nervous system. Nat. Rev. Gastroenterol. Hepatol. 10 (1), 43-57 (2012).
- Sasselli, V., Pachnis, V., Bursn, A. J.
- Sanchez, M. P., Silos-Santiago, I., et al. Renal agenesis and the absence of enteric neurons in mice lacking GDNF. Nature. 382 (6586), 70-73 (1996).
- Baynash, A. G., Hosoda, K., et al. Interaction of endothelin-3 with endothelin-B receptor is essential for development of epidermal melanocytes and enteric neurons. Cell. 79 (7), 1277-1285 (1994).
- Chalazonitis, A., Pham, T. D., et al. Neurotrophin-3 is required for the survival-differentiation of subsets of developing enteric neurons. J. Neurosci. 21 (15), 5620-5636 (2001).
- Goldstein, A. M., Brewer, K. C., Doyle, A. M., Nagy, N., Roberts, D. J. BMP signaling is necessary for neural crest cell migration and ganglion formation in the enteric nervous system. Mech. Dev. 122 (6), 821-833 (2005).
- Jiang, Y., Liu, M. T., Gershon, M. D. Netrins and DCC in the guidance of migrating neural crest-derived cells in the developing bowel and pancreas. Dev. Biol. 258 (2), 364-384 (2003).
- Ramalho-Santos, M., Melton, D. A., McMahon, A. P. Hedgehog signals regulate multiple aspects of gastrointestinal development. Development. 127 (12), 2763-2772 (2000).
- Natarajan, D., Marcos-Gutierrez, C., Pachnis, V., de Graaf, E. Requirement of signaling by receptor tyrosine kinase RET for the directed migration of enteric nervous system progenitor cells during mammalian embryogenesis. Development. 129 (22), 5151-5160 (2002).
- Young, H. M., Hearn, C. J., et al. GDNF Is a chemoattractant for enteric neural cells. Dev. Biol. 229 (2), 503-516 (2001).
- Amiel, J., Sproat-Emison, E., et al. Hirschsprung disease, associated syndromes and genetics: a review. J. Med. Genet. 45 (1), 1-14 (2008).
- Pilon, N., Raiwet, D., Viger, R. S., Silversides, D. W. Novel pre- and post-gastrulation expression of Gata4 within cells of the inner cell mass and migratory neural crest cells. Dev. Dyn. 237 (4), 1133-1143 (2008).
- Nagy, N., Goldstein, A. M. Endothelin-3 regulates neural crest cell proliferation and differentiation in the hindgut enteric nervous system. Dev. Biol. 293 (1), 203-217 (2006).
- Nagy, A., Gertsenstein, M., Vintersen, K., Behringer, R. Manipulating the mouse embryo: a laboratory manual. , 3rd ed, CSH Press. Cold Spring Harbor. 209-250 (2003).
- Schneider, C. A., Rasband, W. S., Eliceiri, K. W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods. 9, 671-675 (2012).
- Byth, K. F., Thomas, A., et al. AZD5438, a potent oral inhibitor of cyclin-dependent kinases 1, 2, and 9, leads to pharmacodynamic changes and potent antitumor effects in human tumor xenografts. Mol. Cancer Ther. 8 (7), 1856-1866 (2009).

{kind=link}