

Summary
Nous présentons une analyse ex vivo de la migration cellulaire qui permet une quantification précise du potentiel de la migration des cellules de la crête neurale entérique en présence de divers facteurs de croissance.
Abstract
Cellules de la crête neurale (CCN) sont une population de cellules multipotentes transitoire et qui provient du tube neural dorsal et migre intensivement dans l'ensemble du développement de l'embryon des vertébrés. En plus de fournir des cellules gliales et neurones périphériques, CCN générer mélanocytes ainsi que la plupart du squelette cranio-facial. La migration et la différenciation CCN est contrôlé par une combinaison de leur origine axiale le long du tube neural et leur exposition aux signaux extracellulaires distinctes suivant les régions. Un tel apport de ligands extracellulaires est particulièrement évident au cours de la formation du système nerveux entérique (ENS), un réseau complexe interconnecté de ganglions nerveux qui commande localement (entre autres) un mouvement intestin musculaire et la motilité intestinale. La plupart de l'ENS est dérivé d'un petit groupe initial de la CCN qui entreprennent un long voyage afin de coloniser - dans un rostre à la mode caudale - toute la longueur de l'intestin prospective. Parmi plusieurs voies de signalisation connues de l'influencer entérique colonisation CCN, le GDNF / signalisation de RET est reconnu comme le plus important. En effet, la sécrétion spatio-temporelle contrôlée du ligand GDNF RET par le mésenchyme de l'intestin est le principal responsable de l'attraction et de l'orientation de RET exprimant entérique CCN et dans l'intestin embryonnaire. Ici, nous décrivons une analyse ex vivo de la migration des cellules, ce qui rend l'utilisation d'une lignée de souris transgéniques possédant CCN marqué par fluorescence, ce qui permet une quantification précise du potentiel de migration CCN entérique en présence de divers facteurs de croissance, y compris le GDNF.
Introduction
Cellules de la crête neurale (CCN) sont un type de cellule transitoire unique de vertébrés qui forme de nombreux dérivés au cours du développement de l'embryon. Cette population de cellules se présente à la frontière de la plaque neurale, à côté de l'ectoderme neural non-1. Au cours de la neurulation, flexion des plaques lieux neuronaux de la CCN le long de la partie dorsale du tube neural se former. CCN subit alors une transition épithéliale-mésenchymateuse, la ségrégation et la migration à partir du tube neural. CCN coloniser différentes structures embryonnaires, y compris le tube digestif où ils forment l'ensemble du système nerveux entérique (SNE), un réseau interconnecté de ganglions nerveux intégré dans la paroi intestinale. Comme l'a récemment examiné 2,3, de nombreux gènes ont été impliqués dans le développement de cette structure complexe.
La plupart de l'ENS est dérivé d'un petit pool de NCC provenant du tube neural vagal (soit de l'ordre de la limite rhombencéphale / moelle épinière prospective) 4.Ces progéniteurs neuronaux atteignent l'intestin antérieur autour du jour embryonnaire (e) 9.0 chez la souris, puis migrent caudalement dans le mésenchyme de l'intestin jusqu'à environ e15.0 de coloniser l'ensemble des intestins embryonnaires. Un sous-groupe mineur de cellules progénitrices neurales du côlon est également fourni par sacral CCN, qui envahissent l'intestin postérieur dans la direction opposée jusqu'à la caecum 4. Les deux vagal et sacrée CCN exige multiples migration, la prolifération, la survie et de signaux de différenciation promotion pour assurer la formation complète de l'ENS. À cet égard, les modèles animaux - souris en particulier génétiquement modifiés - ont joué un rôle dans l'identification de plusieurs ligands extracellulaires essentielles: GDNF (glial facteur neurotrophique dérivé des cellules), l'endothéline-3, la neurotrophine-3, (protéines morphogénétiques osseuses) MPG, Netrin , ainsi que Sonic et Indian Hedgehog (Shh et Ihh) 5-10. Parmi ceux-ci, la signalisation par le GDNF RET de récepteur transmembranaire de la tyrosine kinase (Rearranged pendant la transfection) est reconnu comme l'ee voie la plus critique pour l'attraction et la direction de la CCN et à l'intérieur de l'intestin embryonnaire. GDNF est sécrétée par le mesenchyme de l'intestin et forme un gradient spatio-temporellement rosrrocaudal contrôlé qui est directement chimio-attractifs pour entérique CCN, qui expriment RET 11,12.
Entre autres fonctions, l'ENS règle le mouvement dans le tube digestif par son interaction avec le muscle lisse de la paroi intestinale. Absence de ganglions nerveux dans la région terminale des résultats de l'intestin dans la maladie de Hirschsprung: contraction tonique du segment affecté conduit à un blocage, l'accumulation en amont de la matière digérée et la distension massive de l'intestin et de l'abdomen. La maladie de Hirschsprung se produit environ un sur 5000 naissances vivantes. Le motif de la migration rostro-caudale de entérique CCN est considéré comme le principal facteur contribuant à l'étiologie de la maladie de Hirschsprung. Le côlon, le plus éloigné de la source de migration NCC et dernière partie de bowel être colonisé, est le plus sensible à des défauts dans la formation ENS. Conformément à son rôle crucial dans la migration CCN entérique, la perturbation de la signalisation GDNF / RET est la principale cause génétique connue de la maladie de Hirschsprung 13.
Pour mieux étudier la CCN et le développement de l'ENS, nous avons généré une lignée de souris transgénique - nommé Gata4p [5 kb]-GFP 14 - dans laquelle migratoire CCN sont marquées avec la protéine fluorescente verte (GFP). On a ensuite mis au point un test ex vivo de la migration cellulaire, adapté de travaux publiés par d'autres groupes 11,12,15, qui permet une quantification précise maintenant le potentiel de migration CCN entérique en présence de divers facteurs de croissance, tels que le GDNF.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
Déclaration éthique
Des expériences sur des souris ont été effectuées après les lignes directrices du Conseil canadien de protection des animaux pour le soin et la manipulation des animaux utilisés dans la recherche médicale. Protocoles impliquant la manipulation d'animaux ont été approuvés par le comité d'éthique institutionnels de l'Université du Québec à Montréal (Comité Institutionnel de Protection des Animaux; numéro de référence 0512-R3-650-0513).
Une. Préparation de collagène Gels
Travailler de manière stérile, sous une hotte de culture de tissus.
- Préparer complète 5x DMEM (milieu essentiel modifié Eagle) y compris les antibiotiques standard. Dissoudre 3,37 g de poudre DMEM et 0,925 g de NaHCO 3 dans 20 ml d'eau. Stériliser par passage à travers un filtre de 0,22 um. Ajouter 2,5 ml de 100x stérile de pénicilline / streptomycine et de 25 ml de sérum bovin foetal inactivé par la chaleur stérile. Stocker à 4 ° C.
- Sur la glace, mélanger 800 pisolution de collagène I (3,77 mg / ml dans 0,02 N d'acide acétique, stérilisée par filtration), 600 ul de DMEM complet 5x et 17 ul de 1 N NaOH. Diluer avec de l'eau stérile jusqu'à un volume final de 3 ml. Inclure des facteurs de croissance pertinentes dans le mélange. Nous utilisons généralement GDNF à 10 ng / ml pour stimuler la migration CCN entérique.
- Dépôt d'environ 480 pi dans chaque puits d'une seule ligne dans une plaque de 24 puits. Éviter les bulles. Les lignes restantes peuvent être utilisées pour tester l'effet d'autres facteurs de croissance sur la migration cellulaire.
- Laissez le collagène polymériser au moins 1 h dans un incubateur stérile à 37 ° C.
2. Dissection des animaux 16
- Mettre en place des accouplements et vérifier bouchon vaginal le lendemain matin. 0,5 ME de jour étant midi le jour du bouchon vaginal est trouvé, isoler la femelle et attendre 12 jours jusqu'à ce que E12.5.
- Anesthésier les souris enceinte avec de l'isoflurane et l'euthanasie par inhalation de CO 2.
- Pulvériser la souris avec 70% d'éthanol. Soulevez les s abdomenparents et ouvrir la cavité de l'abdomen avec des ciseaux de dissection.
- Retirer l'utérus dans une boîte de Pétri en verre rempli avec du PBS glacé (solution saline tamponnée au phosphate). Couper l'utérus transversalement entre gonflements deciduum individuels d'isoler chaque site d'implantation de l'embryon.
- Les travaux sur chaque site d'implantation séparément dans une autre boîte de Pétri en verre rempli de PBS glacé. Sous un microscope de dissection, utiliser une pince fine pour enlever la couche musculaire de l'utérus.
- Ouvrez le sac jaune viscérale et l'amnios de révéler l'embryon. Faites attention lorsque vous couper les vaisseaux sanguins se joindre à l'embryon au placenta / viscérale jaune sac, car ils sont étroitement liés avec les intestins en développement.
- Couper la tête de l'embryon au niveau du col.
- Insérez une pince fermés dans la cavité abdominale d'un embryon, juste au-dessus du foie de couleur rouge foncé et laisser la pince ouverte d'elle-même (cesser d'appliquer la pression) pour faire une ouverture transversale dans la cavité de l'abdomen. Tirez les pinces ouvertes vers le basl'extrémité postérieure de l'embryon pour ouvrir l'abdomen complètement.
- Prenez le tissu conjonctif derrière le foie et tirer les tripes de l'abdomen, en faisant attention de ne pas casser les intestins (colon est attaché à l'anus).
- Couper le côlon pour libérer les entrailles du reste de l'embryon. La coupe peut être effectué à n'importe quel endroit le long du côlon. Réserver la partie de queue de côté.
- Démêler le tissu conjonctif du caecum, puis le reste de l'intestin. Faites attention de ne pas blesser les intestins en le faisant.
- Couper le foie et l'estomac (à l'extrémité rostrale de l'intestin grêle), ainsi que le mésonéphros et crêtes génitales si certains sont présents.
- Isoler l'intestin grêle. Encore une fois, attention à ne pas étouffer l'intestin. A partir de maintenant, l'orientation rosrrocaudal du tissu intestinal peut être suivi à l'aide de la forte courbure présente sur l'extrémité rostrale. Laissez l'intestin grêle dans du PBS à température ambiante pendant un temps aussi court que possible avant de luiliterie (voir l'étape 3.3).
- Enfin, notez le numéro de la queue somites pour chaque embryon de déterminer avec précision le stade de développement embryonnaire.
3. Sectionnement de embryonnaires intestins
- Avant de procéder à la dissection de l'embryon, faire fondre 1,5 g d'agarose dans 100 ml de PBS (solution saline tamponnée au phosphate) et maintenir à 50 ° C.
- Verser fondu agarose dans un moule intégration (par exemple, un fermé 2 ml microtube qui a été coupé sur la longueur d'exciser environ 1/4 de la paroi du tube). Laissez refroidir l'agarose à environ 42-45 ° C, il devrait être que légèrement chaude au toucher.
- Incluez l'intestin embryonnaire juste avant la solidification d'agarose (ce qui peut être évaluée à l'aide de forceps conseils et se fera autour de 36-38 ° C). La tenue de l'intestin avec une pince par l'extrémité rostrale pliée, le tirer très lentement à travers l'agarose long de la longueur du moule. Cela aide à garder l'intestin droite tandis que les ensembles d'agarose. Relâchez le témettre dès qu'elle commence à résister à être déplacé. Gardez une trace de l'orientation de la rostrale-caudale de l'intestin.
- Mettez le moule au réfrigérateur 2-3 minutes, afin d'assurer la gélose a complètement réglée.
- Sortir le agarose à partir du moule (en le faisant glisser hors du tube Eppendorf ouvert). Avec une lame, retirer l'excédent d'agarose à ses deux extrémités, des coupes perpendiculaires à l'intestin.
- Collez l'extrémité rostrale de l'intestin / agarose bloquer sur le stade du métal d'un microtome à lame vibrante. Couper l'excédent d'agarose sur les côtés du bloc intestin / d'agarose.
- Monter la phase métallique sur la chambre de microtome vibrant. Si nécessaire, ajuster l'angle de la scène si l'intestin est aussi vertical que possible (donc perpendiculaire à la lame). Couvrez le spécimen avec PBS glacé. Amener la lame de microtome vers le bas à quelques millimètres sous la surface du tampon.
- Faire 200 um vibratome coupes transversales de l'intestin caudal le plus petit, veiller à ce que ehaque tranche d'agarose contient une section complète intestinale.
4. Culture de Intestinal explants
- Déposer délicatement les tranches intestin / d'agarose fraîchement coupées à plat sur les gels de collagène avec une pince, placer une tranche vers le milieu de chaque puits.
- Incuber 3 jours à 37 ° C, dans un environnement humide de 5% de CO 2, atmosphère, pour permettre la migration du CCN sur l'explant.
- Prendre les tranches intestin / d'agarose le gel de collagène très doucement avec une pince. Prenez soin de ne pas endommager le gel ci-dessous.
- Éviter de toucher le gel de collagène directement au cours de l'incubation et de lavage des étapes ultérieures, afin de s'assurer que le motif de la migration des cellules ne soit pas perturbé. Fixer avec 500 pl de 4% de PFA (paraformaldéhyde) (dans du PBS) par puits 1 heure à température ambiante.
- Remplacer le fixateur avec 500 ul de DAPI (4 ',6-diamidino-2-phénylindole) solution (5 ug / ml dans du PBS) par puits et incuber 10 min à température ambiante.
- Laver chaque puits 3xavec 500 ul de PBS pendant 5 min.
- Photographier les cellules fluorescentes (canaux de GFP et DAPI) incorporés dans le gel de collagène à l'intérieur de chaque puits.
5. Analyse de l'image
Nous avons fait un usage intensif de ImageJ 17 pour traiter et quantifier les images générées après culture des explants.
- Commencer par l'imagerie d'un coulisseau de micromètre au même grossissement que les photographies de cellules fluorescentes. Mesurer la longueur du micron en pixels (en utilisant l'outil de ligne droite).
- Définissez l'échelle (Analyser / Set échelle, le nombre d'entrée de pixels / micron).
- Pour chaque photographie de fluorescence de la GFP, changer le format de l'image en niveaux de gris de 8 bits (Image/Type/8-bit).
- Réglez l'intensité du signal (image / Réglage / Luminosité / Contraste).
- Soustraire le bruit de fond si nécessaire (processus / Soustraire fond; ajuster le rayon de roulement à billes).
- Fixer un seuil de mettre en évidence les cellules et les amas de cellules (image / Réglage / Seuil), len appliquer un bassin versant à diviser les touffes (Process / binaire / bassins versants).
- Analyser des particules de préciser les régions d'intérêt (ROI) et de générer des statistiques de nombre de cellules (Analyser / analyser des particules; ensemble taille minimale pour exclure les pixels restants).
- Définir les options de mesure d'inclure le diamètre de Feret (Analyser / Mesures Set).
- Groupe ROI et mesurer dans son ensemble pour déterminer le diamètre de la Feret, une indication de la propagation des cellules (Analyser / Outils / Gestionnaire de ROI / Plus / OU, puis Directeur ROI / Ajouter, et directeur de ROI / Mesure).
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
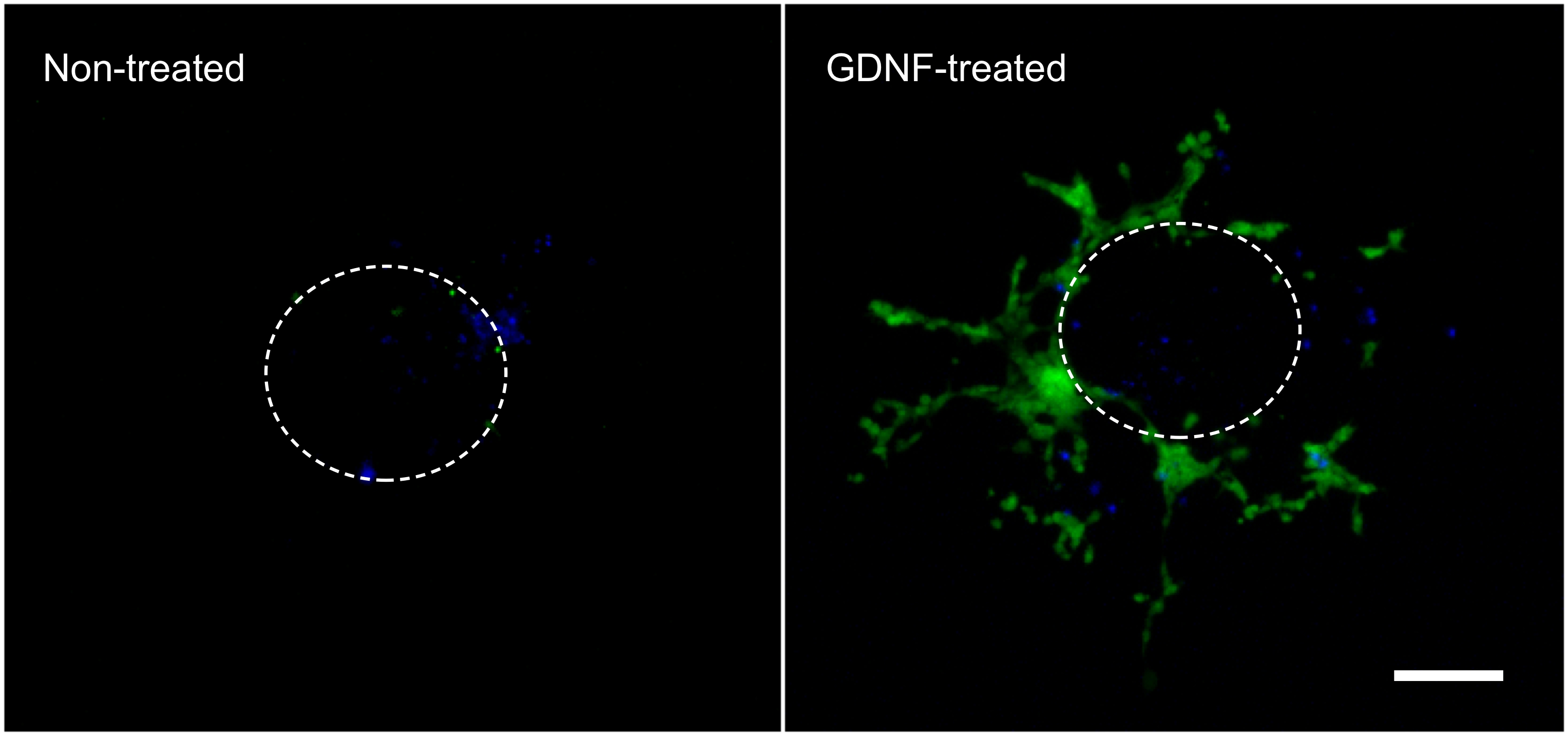
Les résultats suivants sont représentatifs de ce qui peut être obtenu avec la technique décrite ici (figure 1). L'utilisation de facteurs de croissance (c.-à-GDNF) stimule la migration de la GFP exprimant entérique CCN rupture de l'explant intestinal et dans le gel de collagène (figure 2). Bien que certaines cellules sortent de l'explant en l'absence de facteurs de croissance, ceux-ci sont pour la plupart pas GFP-étiqueté et représentent entrée passive. Il est nécessaire d'enlever la tranche intestinale du gel de collagène afin d'enregistrer les résultats, que ce tissu est encore peuplée par les cellules fluorescentes et serait contraire masquer les cellules se trouvant dans le collagène dessous. La quantification de ces résultats montre que beaucoup plus de cellules se trouvent dans le gel de collagène quand GDNF est présent, et que la migration a lieu actif (Figure 3). En effet, les cellules passives se trouvent immédiatement sous les explants, à l'intérieur du diamètre d'une tranche intestinal, wher EAS cellules qui envahissent activement le gel de collagène se déplacent loin de leur point d'origine et la propagation plus loin.

Figure 1. Vue d'ensemble de la technique de culture d'explant. A) de la population uniforme de fluorescence entérique CCN dans le petit intestin région caudale utilisé pour fabriquer des explants 200 μ d'épaisseur. Barre d'échelle:. 200 um B) Le milieu de culture contenant du collagène est déposée dans des plaques à 24 puits et on laisse durcir pendant 1 heure. Vibratome tranches de intestins embryonnaires agarose-intégré sont déposées sur les gels (une tranche par puits), et fluorescent entérique CCN sont autorisés à migrer hors des explants pendant 3 jours. Les tranches sont ensuite retirés avant l'imagerie des cellules qui ont envahi les gels de collagène.s/ftp_upload/50709/50709fig1highres.jpg "target =" _blank "> Cliquez ici pour agrandir l'image.

Figure 2. La migration des cellules de l'explant intestinal et dans le gel de collagène. cellules qui ont migré à partir d'un explant intestinal au cours de l'incubation de 3 jours en l'absence ou en présence de 10 ng / ml GDNF ont été fixées, colorées avec du DAPI (bleu) et photographié à montrer GFP marqué entérique CCN (vert). Grossissement 70X. Barre d'échelle: 100 um. La ligne pointillée représente la taille et l'emplacement approximatifs de l'expiant avant qu'il ne soit retiré du gel. Cliquez ici pour agrandir la figure. Cliquez hERE pour agrandir l'image.

Figure 3. Quantification de potentiel entérique de migration CCN. Le nombre et la propagation (diamètre de Feret) de cellules exprimant la GFP qui migrent sur des explants intestinaux après 3 jours a été quantifiée en utilisant le logiciel ImageJ 17. Dans les deux cas, il existe une différence significative entre les conditions non traitées et GDNF traités selon le test t de Student (p <0,001; *). Le diamètre moyen des tranches intestinaux (pointillé ligne noire: 260 um) a été inclus pour distinguer entre l'entrée passive et migration active. Nt: non traitée, n. Nombre d'explants transformés Cliquez ici pour agrandir l'image .
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Nous montrons comment notre ex vivo technique de culture des explants peut être utilisé pour quantifier précisément le potentiel de migration entérique CCN en présence de GDNF. Une telle quantification précise est grandement facilitée par l'utilisation de 200 sections vibratome intestinale um d'épaisseur à la place de grands morceaux de la taille approximative, comme décrit précédemment 11,12,15. En effet, cela nous permet de travailler avec un nombre raisonnable de cellules dans un cadre hautement reproductible. Il faut noter que la distribution uniforme du CCN fluorescent entérique à l'intérieur de la région caudale-la majeure partie de l'intestin grêle à partir de laquelle les tranches d'explants sont coupés permet également l'analyse de plusieurs sections à partir d'un seul tube digestif (Figure 1A). En outre, étant donné que les deux CCN et axones entérique peuvent quitter le tissu dans de tels dosages 11, retrait des explants à la fin de la période de culture nous permet de se concentrer exclusivement sur migratoire CCN.
La plupart des étapes critiques ont été décrites dans le texte du protocole, mais comme le wBien-être social de l'expiant intestinale est primordiale pour obtenir sain CCN migration, un soin particulier doit être pris. Eviter de soumettre les tissus embryonnaires à des changements brusques de température, en particulier lorsque l'intestin est intégré dans agarose (étape 3.3). Assurez-vous que l'intestin est à la température ambiante et l'agarose aussi frais que possible (mais encore fondu) pour éviter «cuisson» le tissu. L'explant intestinal doit se développer sur le gel de collagène du milieu de culture rempli, souvent augmenter en taille et en débordant de la tranche d'agarose. Si l'expiant semble malsain ou pire, tend à périr pendant l'incubation, essayez de remplacer le PBS avec des milieux de culture à la température ambiante (par exemple M2 tampon HEPES ou DMEM supplémenté avec 10% de FBS) pour aider à maintenir pendant la dissection.
Une limitation importante de notre approche est qu'elle repose sur la disponibilité d'une lignée de souris conférant un marqueur fluorescent à la migration de la CCN. En l'absence d'une telle ressource, un anticorps contre le migrating CCN (par exemple, anti-Ret ou anti-Sox10) peut être utilisé pour marquer les cellules qui ont envahi le gel de collagène. En outre, étant donné que l'intestin micro-environnement est beaucoup plus complexe que d'un gel de collagène simple, les résultats obtenus avec ce dosage in vitro pourraient ne pas refléter entièrement le comportement de entérique CCN in vivo. Des expériences supplémentaires impliquant imagerie des cellules vivantes sont recommandés pour évaluer ce comportement. Il est également intéressant de noter que, en plus de son rôle de chemoattractant, le GDNF est connue pour favoriser la prolifération de la migration entérique CCN 4. Notre mesure du potentiel de migration CCN entérique, en présence de GDNF est donc probablement un mélange de véritable migration cellulaire et la prolifération cellulaire, semblable aux mécanismes in vivo conduisant à NCC colonisation de l'intestin. Si une distinction claire entre ces deux processus est souhaitée, l'addition d'un inhibiteur du cycle cellulaire (par exemple AZD 5438 18) dans le milieu de culture peut limiter l'analyse à migratio cellulairen.
Cette technique peut être étendue à l'essai de divers autres ligands extracellulaires, ainsi que des inhibiteurs de voies de signalisation spécifiques, et toute combinaison de celui-ci. D'autres tissus peuvent aussi éventuellement être disséqués et sectionnés, ce qui permet l'étude de la migration de la CCN dans de nombreuses structures embryonnaires. Combiné avec roman et / ou souches de souris mutantes non caractérisés avec d'éventuels défauts dans le développement de la CCN, notre technique peut être appliquée à déceler rapidement les lacunes dans le comportement de migration en réponse à des événements spécifiques de signalisation.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Les auteurs n'ont rien à révéler.
Acknowledgments
Nous remercions Denis Flipo pour obtenir des conseils de traitement et d'analyse d'images, et David W. Silversides en laboratoire dont la ligne de la souris Gata4p [5 kb]-GFP a été générée. La recherche dans le laboratoire Pilon est financé par les IRSC, le CRSNG, FRQS et FRQNT.
Materials
| Name | Company | Catalog Number | Comments |
| DMEM powder | Wisent | 219-010-XK | |
| NaHCO3 | Bioshop | SOB999 | Biotechnology grade |
| Steriflip vacuum filtration system (0.22 micron) | EMD Millipore | SCGP00525 | |
| Penicilin-Streptomycin solution, 100x | Wisent | 450-201-EL | |
| Fetal bovine serum | Wisent | 095-150 | High quality grade |
| Collagen I | BD biosciences | 354236 | |
| NaOH | Bioshop | SHY700 | Diluted from 10 N stock then sterile-filtered |
| GDNF | Cedarlane | CLCYT305 | |
| Falcon 24-well Plate | BD biosciences | 353047 | |
| Dissecting scissors | Fisher Scientific | 089515 | |
| Glass Petri dish | VWR | 89000-306 | |
| PBS | Sigma | P5493 | Cell culture grade |
| Dissecting microscope | Leica | M125 | |
| Dumont #5 Forceps | Fine Science Tools | 11251-20 | |
| Agarose | Bioshop | AGA001 | Biotechnology grade |
| Surgical blade | Feather | 21 | |
| All Purpose Instant Krazy Glue Pen | Krazy Glue | KG824 | |
| HM 650V Vibrating-Blade Microtome | Thermo Scientific | 920110 | |
| Paraformaldehyde | Sigma-Aldrich | P6148 | |
| DAPI | Sigma-Aldrich | D9564 | |
Table of specific reagents and equipment. |
|||
References
- Bronner, M. E., Le Douarin, N. M. Development and evolution of the neural crest: An overview. Dev. Biol. 366 (1), 2-9 (2012).
- Bergeron, K. F., Silversides, D. W., Pilon, N. The developmental genetics of Hirschsprung's disease. Clin. Genet. 83 (1), 15-22 (2013).
- Obermayr, F., Hotta, R., Enomoto, H., Young, H. M. Development and developmental disorders of the enteric nervous system. Nat. Rev. Gastroenterol. Hepatol. 10 (1), 43-57 (2012).
- Sasselli, V., Pachnis, V., Bursn, A. J.
- Sanchez, M. P., Silos-Santiago, I., et al. Renal agenesis and the absence of enteric neurons in mice lacking GDNF. Nature. 382 (6586), 70-73 (1996).
- Baynash, A. G., Hosoda, K., et al. Interaction of endothelin-3 with endothelin-B receptor is essential for development of epidermal melanocytes and enteric neurons. Cell. 79 (7), 1277-1285 (1994).
- Chalazonitis, A., Pham, T. D., et al. Neurotrophin-3 is required for the survival-differentiation of subsets of developing enteric neurons. J. Neurosci. 21 (15), 5620-5636 (2001).
- Goldstein, A. M., Brewer, K. C., Doyle, A. M., Nagy, N., Roberts, D. J. BMP signaling is necessary for neural crest cell migration and ganglion formation in the enteric nervous system. Mech. Dev. 122 (6), 821-833 (2005).
- Jiang, Y., Liu, M. T., Gershon, M. D. Netrins and DCC in the guidance of migrating neural crest-derived cells in the developing bowel and pancreas. Dev. Biol. 258 (2), 364-384 (2003).
- Ramalho-Santos, M., Melton, D. A., McMahon, A. P. Hedgehog signals regulate multiple aspects of gastrointestinal development. Development. 127 (12), 2763-2772 (2000).
- Natarajan, D., Marcos-Gutierrez, C., Pachnis, V., de Graaf, E. Requirement of signaling by receptor tyrosine kinase RET for the directed migration of enteric nervous system progenitor cells during mammalian embryogenesis. Development. 129 (22), 5151-5160 (2002).
- Young, H. M., Hearn, C. J., et al. GDNF Is a chemoattractant for enteric neural cells. Dev. Biol. 229 (2), 503-516 (2001).
- Amiel, J., Sproat-Emison, E., et al. Hirschsprung disease, associated syndromes and genetics: a review. J. Med. Genet. 45 (1), 1-14 (2008).
- Pilon, N., Raiwet, D., Viger, R. S., Silversides, D. W. Novel pre- and post-gastrulation expression of Gata4 within cells of the inner cell mass and migratory neural crest cells. Dev. Dyn. 237 (4), 1133-1143 (2008).
- Nagy, N., Goldstein, A. M. Endothelin-3 regulates neural crest cell proliferation and differentiation in the hindgut enteric nervous system. Dev. Biol. 293 (1), 203-217 (2006).
- Nagy, A., Gertsenstein, M., Vintersen, K., Behringer, R. Manipulating the mouse embryo: a laboratory manual. , 3rd ed, CSH Press. Cold Spring Harbor. 209-250 (2003).
- Schneider, C. A., Rasband, W. S., Eliceiri, K. W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods. 9, 671-675 (2012).
- Byth, K. F., Thomas, A., et al. AZD5438, a potent oral inhibitor of cyclin-dependent kinases 1, 2, and 9, leads to pharmacodynamic changes and potent antitumor effects in human tumor xenografts. Mol. Cancer Ther. 8 (7), 1856-1866 (2009).

{kind=link}