

Summary
Vi presenterer en ex vivo-cellemigrering assay som tillater presis kvantifisering av enterisk neural crest celle migrasjon potensiale i nærvær av forskjellige vekstfaktorer.
Abstract
Neural crest cellene (NCC) er en forbigående og multipotent cellepopulasjon som stammer fra rygg nevralrøret og vandrer mye i hele utviklings virveldyr embryo. I tillegg til å gi perifere gliaceller og nevroner, NCC generere melanocytter samt det meste av cranio-ansikts skjelett. NCC migrasjon og differensiering er kontrollert av en kombinasjon av deres aksial opprinnelse langs nevralrøret og deres eksponering for regionalt distinkte ekstracellulære signaler. Slike er bidraget av ekstracellulære ligander spesielt tydelig under dannelse av den enterisk nervesystem (ENS), et komplekst sammenvevd nettverk av nervegangliene som lokalt styrer (blant annet) gut muskelbevegelse og intestinal motilitet. Mesteparten av ENS er avledet fra en liten innledende pool av NCC at foreta en lang reise for å kolonisere - i et rostral til caudal mote - hele lengden av den potensielle tarmen. Blant flere signalveier er kjent for åpåvirke enteric NCC kolonisering, er GDNF / RET signaleanerkjent som den viktigste. Faktisk er spatiotemporally kontrollert sekresjon av RET ligand GDNF av tarmen mesenchyme hovedsakelig ansvarlig for tiltrekning og veiledning av RET-uttrykke enteric NCC til og innenfor den embryonale gut. Her beskriver vi en ex vivo-cellemigrering assay, som gjør bruk av en transgen muselinje som innehar fluorescensmerkede NCC, som tillater presis kvantifisering av enteriske NCC migrasjon potensiale i nærvær av forskjellige vekstfaktorer, inkludert GDNF.
Introduction
Neural crest cellene (NCC) er en forbigående celletype som er unik for virveldyr som danner mange derivater under embryoutvikling. Denne cellen befolkningen oppstår på grensen av det nevrale plate, som grenser til ikke-nevrale ektoderm en. Under neurulation, bøying av neural plate steder NCC langs ryggkanten danner neural røret. NCC deretter gjennomgå en epitelial-mesenkymale overgang, segregerende og migrere bort fra nevralrøret. NCC kolonisere forskjellige embryonale strukturer, inkludert i fordøyelseskanalen, hvor de danner en hel enterisk nervesystem (ENS), et sammenkoplet nettverk av nervegangliene innleiret i tarmveggen. Som nylig anmeldt 2,3, har mange gener er involvert i utviklingen av dette intrikate struktur.
Mesteparten av ENS er avledet fra en liten mengde av NCC stammer fra neural vagal-røret (dvs. rundt den potensielle hindbrain / ryggmarg grense) 4..Disse nevrale stamceller nå forutgående rundt embryonale dag (e) 9,0 hos mus, og deretter vandrer caudally i tarmen mesenchyme inntil ca e15.0 å kolonisere hele embryonale tarmen. En liten undergruppe av colonic nevrale stamceller er også forsynt med sakral NCC, som invaderer den bakre tarm i motsatt retning opp til cecum 4.. Både vagal og sakrale NCC krever flere migrasjon-, spredning-, overlevelses-og differensieringsfremmende signaler for å sikre komplett dannelsen av ENS. I denne forbindelse, dyremodeller - spesielt genmodifiserte muse - har vært medvirkende i identifisering av flere essensielle ekstracellulære ligander: GDNF (glial celle-avledet neurotrophic factor), Endotelin-3, neurotrophin-3, BMP (benmorfogent proteiner), Netrin , samt Sonic og indiske Hedgehog (Shh og IHH) 5-10. Av disse er GDNF signalering gjennom tyrosin kinase transmembrane reseptor RET (Rearranged under transfeksjon) anerkjent som the mest kritisk vei for tiltrekning og veiledning av NCC til og i den embryonale gut. GDNF skilles ut av tarmen mesenchyme og danner en spatiotemporally kontrollert rosrrocaudal gradient som er direkte chemoattractive til ente NCC, som uttrykker RET 11,12.
Blant andre funksjoner, ENS regulerer bevegelsen i fordøyelseskanalen gjennom dets interaksjon med glatt muskulatur i tarmveggen. Fravær av nevrale ganglier i terminalen regionen i tarmen resulterer i Hirschsprung sykdom: tonic sammentrekning av de berørte segmentet fører til blokkering, oppstrøms opphopning av fordøyd materiale og massive oppblåsthet i tarmen og magen. Hirschsprung sykdom oppstår omtrent ett i 5000 levendefødte. Den rostro-caudal vandringsmønster av ente NCC antas å være den viktigste medvirkende faktor til etiologi av Hirschsprung sykdom. Tykktarmen, lengst fra kilden til å migrere NCC og siste del av bOwel å bli kolonisert, er mest utsatt for feil i ENS formasjon. I samsvar med sin avgjørende rolle i ente NCC migrasjon, er forstyrrelse av GDNF / RET signale den viktigste kjente genetiske årsaken til Hirschsprung sykdom 13.
For bedre å studere NCC og ENS utvikling, genererte vi en transgen muselinje - oppkalt Gata4p [5kb]-GFP 14 - der vandrende NCC er merket med grønt fluorescerende protein (GFP). Vi neste perfeksjonert en ex vivo-cellemigrering analyse tilpasset fra publiserte arbeide av andre grupper 11,12,15, som nå tillater presis kvantifisering av enteriske NCC migrasjon potensiale i nærvær av forskjellige vekstfaktorer, slik som GDNF.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
Etikk uttalelse
Eksperimenter med mus ble utført følgende Canadian Council of Animal Care retningslinjer for omsorg og manipulering av dyr som brukes i medisinsk forskning. Protokoller som involverer manipulering av dyr ble godkjent av institusjonelle etisk komité ved University of Quebec i Montreal (Comité Institutionnel de Protection des Animaux, referansenummer 0512-R3-650-0513).
En. Utarbeidelse av kollagen Gels
Arbeid i et sterilt mote, under en vevskultur panseret.
- Forbered komplett 5x DMEM (Eagle modifiserte viktig medium) inkludert standard antibiotika. Oppløs 3,37 g av DMEM pulver og 0,925 g NaHCO3 i 20 ml vann. Steriliser ved å passere gjennom et 0,22 mikrometer filter. Til 2,5 ml sterilt 100x penicillin / streptomycin og 25 ml sterilt varme-inaktivert føtalt bovint serum. Oppbevar ved 4 ° C.
- På is, blande 800 mLof Collagen I-løsning (3,77 mg / ml i 0,02 N eddiksyre, filtersterilisert), 600 pl av komplett 5x DMEM og 17 pl av 1 N NaOH. Fortynn med sterilt vann til et sluttvolum på 3 ml. Ta med relevante vekstfaktorer i miksen. Vi bruker vanligvis GDNF på 10 ng / ml for å stimulere enteric NCC migrasjon.
- Innskudd omtrent 480 ul i hver brønn i en enkelt rad i en 24-brønns plate. Unngå bobler. De gjenværende rader kan brukes for å teste effekten av andre vekstfaktorer på cellemigrasjon.
- La kollagen polymerisere minst 1 time i en steril inkubator ved 37 ° C.
2. Dissection of Animals 16
- Sett opp parringer og se etter vaginal plugger neste morgen. Dag e0.5 blir middag på dagen vaginal plug er funnet, isolere den kvinnelige og vente 12 dager før E12.5.
- Anesthetize gravide mus med isofluran og avlive ved CO 2 innånding.
- Spray musen med 70% etanol. Løft magen spårørende og åpne magen hulrom med dissekere saks.
- Fjern livmoren i en glasspetriskål fylt med iskald PBS (fosfatbuffret saltoppløsning). Skjær livmoren tvers mellom individuelle deciduum hevelser å isolere hver embryoimplantasjonen nettstedet.
- Arbeidet med hvert implantasjon nettstedet separat i et annet glass petriskål fylt med iskald PBS. Under et dissekerende mikroskop, bruke fin pinsett for å fjerne muskel lag av uterus.
- Åpne visceral plommesekken og amnion å avsløre embryo. Vær forsiktig når du kutte blodårene bli med embryo til placenta / visceral plommesekken, som de henger sammen med utviklings tarmen.
- Sever embryo hode i nakken.
- Sett en lukket tang i bukhulen av et embryo, like over den mørke rød-farget leveren og la tang åpen av seg selv (slutter å bruke press) for å lage en tverrgående åpning i magen hulrom. Trekk de åpnede tang ned motden bakre ende av embryoet for å åpne buken helt.
- Ta tak i bindevevet bak leveren og trekk guts ut av magen, være forsiktig med å bryte tarmen (tykktarmen er festet til anus).
- Kutt kolon å frigjøre guts fra resten av embryoet. Kan gjøres kutt på ethvert sted langs kolon. Forbeholder halen del for senere.
- Erte ut bindevev fra cecum, da resten av tarmen. Vær forsiktig så du ikke å såre tarmen mens de gjør det.
- Skjær bort lever og mage (på rostral enden av tynntarmen), så vel som de mesonephros og genital rygger hvis noen er til stede.
- Isoler tynntarmen. Igjen, vær forsiktig med å nappe tarmen. Fra nå av kan det rosrrocaudal orientering i tarmvevet spores ved hjelp av skarpe kurver stede på rostral enden. La tynntarmen i PBS ved romtemperatur i så kort tid som mulig før emsengetøy (se trinn 3.3).
- Til slutt, registrere antall halen somites for hvert embryo å nøyaktig fastslå stadium av fosterutviklingen.
Tre. Seksjonering av Embryonale Innvoller
- Før fortsetter med embryo dissections, smelte 1,5 g agarose i 100 ml PBS (fosfatbufret saltoppløsning) og holdes i 50 ° C.
- Hell smeltet agarose innebygging i en form (for eksempel en lukket 2 ml mikrosentrifugerør som har blitt kuttet i lengderetningen for å skjære ca 1/4 av rørveggen). La agarose kjøles ned til ca 42-45 ° C, bør det være bare litt varm å ta på.
- Legge den embryonale tarmen like før agarose stivner (dette kan evalueres med tang tips og vil skje rundt 36-38 ° C). Holde tarmen med tang ved den brettede rostral ende, trekke det meget langsomt gjennom agarose langs lengden av formen. Dette bidrar til å holde den rett mens tarmen agarose-apparater. Slipp tutstede så snart det begynner å motstå å bli flyttet. Hold orden på rostral-caudal retning av tarmen.
- Sett formen i kjøleskap i 2-3 minutter, for å sikre agarose har helt satt.
- Ta ut agarose fra formen (ved å skyve den ut av det åpnede Eppendorf-rør). Med et blad, ta ut det overskytende agarose i begge ender, slik at kutt perpendikulært til tarmen.
- Lim rostral ende av tarmen / agarose blokkere ned på metall stadium av en mikrotom med vibrerende blad. Reduser det overskytende agarose på sidene av tarmen / agarose-blokk.
- Monter metall scenen på vibrerende mikrotom kammeret. Hvis det er nødvendig, justere vinkelen på scenen slik at tarmen er så vertikal som mulig (derav vinkelrett på bladet). Dekk prøven med iskald PBS. Bring mikrotomen bladet ned til noen få millimeter under buffer overflate.
- Gjør 200 mikrometer vibratome tverrgående kutt av hale-mest tynntarmen, slik at each agarose skive inneholder en full intestinal-delen.
4. Culture of Intestinal explants
- Forsiktig deponere nyklipt tarmen / agarose skiver flatt på kollagen gels med tang, plassere en skive mot midten av hver brønn.
- Inkuber 3 dager ved 37 ° C, i en fuktig 5% CO2 atmosfære, for å tillate migrering av NCC ut av eksplantering.
- Ta tarmen / agarose skiver av kollagen gel veldig forsiktig med pinsett. Vær forsiktig så du ikke skader gel nedenfor.
- Unngå å berøre den kollagen-gel direkte i de etterfølgende inkubering og vasketrinn, for å sikre at cellemigreringsmønsteret ikke blir forstyrret. Fiks med 500 pl av 4% PFA (paraformaldehyd) (i PBS) per brønn i 1 time ved romtemperatur.
- Sett på fikser med 500 mL av DAPI (4 ',6-diamidino-2-phenylindole) løsning (5 ug / ml i PBS) per brønn og inkuber i 10 min ved romtemperatur.
- Vask hver brønn 3xmed 500 ul PBS i 5 min.
- Fotografere fluorescerende celler (GFP og DAPI kanaler) innebygd i kollagen gel innenfor hver brønn.
5. Bildeanalyse
Vi gjorde utstrakt bruk av ImageJ 17 å behandle og kvantifisere bilder generert etter eksplantering kultur.
- Start med å avbilde en mikrometer sklie på samme forstørrelse som fluorescerende celle fotografier. Mål lengden på én mikrometer i piksler (lineært verktøyet).
- Still skalaen (Analyze / Set Scale; innspill antall piksler / mikron).
- For hver GFP fluorescens fotografi, endre bildeformatet til 8-bits gråtoner (Image/Type/8-bit).
- Juster intensiteten av signalet (Bilde / Tilpass / lysstyrke / kontrast).
- Trekk fra bakgrunnsstøy om nødvendig (Prosess / Trekk Bakgrunn, justere rullende ball radius).
- Sett en terskel for å markere celler og celleklumper (Bilde / Tilpass / Terskel), denn gjelder et vannskille for å dele klumper (Prosess / Binary / Watershed).
- Analyser partikler til å angi områder av interesse (ROI) og generere celle nummer statistikk (Analyser / Analyser Partikler; sett størrelse minimum for å utelukke gjenværende piksler).
- Sett måle alternativer for å inkludere Feret diameter (Analyser / Set Målinger).
- Gruppe ROIs og måle som helhet til å bestemme Feret diameter, en indikasjon på celle spredning (Analyse / Verktøy / ROI ansvarlig / Mer / ELLER, så ROI ansvarlig / Legg, og ROI ansvarlig / Measure).
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
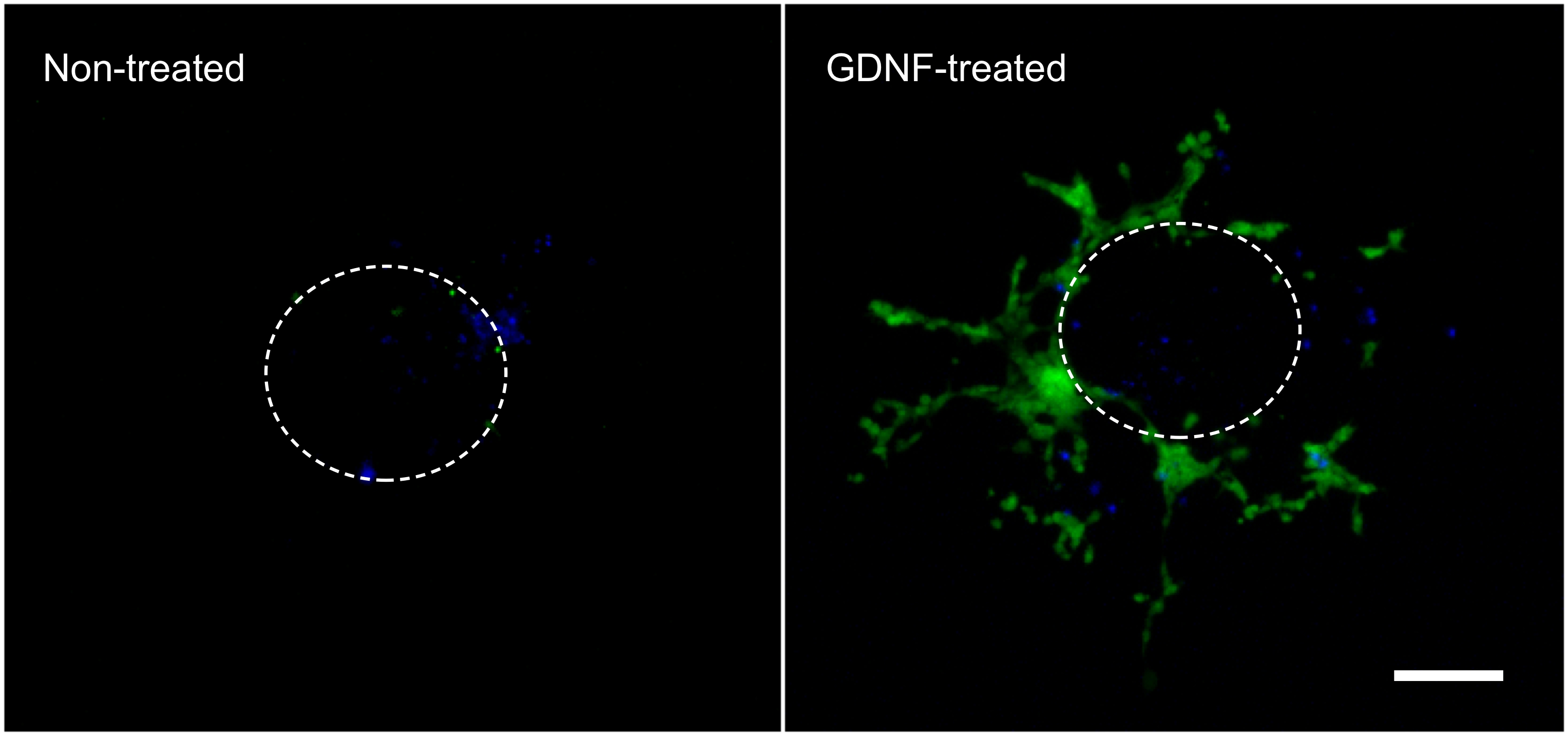
De følgende resultater er representative for det som kan oppnås med den teknikk som er beskrevet her (fig. 1). Bruken av vekstfaktorer (f.eks GDNF) stimulerer migrasjonen av GFP-uttrykkende enteriske NCC ut av intestinal eksplantering og inn i kollagen-gel (figur 2). Skjønt noen celler kommer ut av eksplantering i fravær av vekstfaktorer, er disse stort sett ikke GFP-merket og representerer passive oppføring. Det er nødvendig å fjerne intestinal stykke fra kollagen-gel for å registrere resultatene, da dette vevet er fremdeles tett befolket av fluorescerende celler, og som ellers ville skjule de celler som ligger i kollagen under. Kvantifisering av disse resultatene viser at mange flere celler er funnet i det kollagen-gel når GDNF er til stede, og at aktive migrering finner sted (figur 3). Faktisk er passive celler syntes umiddelbart under eksplantater, innenfor diameteren av en intestinal skive, der hvor EAS celler som aktivt invaderer kollagen gel bevege seg bort fra deres synspunkt opprinnelse og spre videre.

Figur 1. Oversikt over explant kultur teknikk. A) Uniform befolkning på fluorescerende ente NCC innen haletynntarmen regionen brukes til å lage 200 μ-tykke eksplantater. Skala bar:. 200fim B) Et kulturmedium inneholdende kollagen er deponert i 24-brønners plater, og igjen å herde i 1 time. Vibratome skiver av agarose-embedded embryonale tarmen er deponert på gels (en skive per brønn), og fluorescerende enteric NCC har lov til å migrere ut av explants for tre dager. Skivene blir så tatt ut før avbildning cellene som invaderte kollagen geler.s/ftp_upload/50709/50709fig1highres.jpg "target =" _blank "> Klikk her for å se større bilde.

Figur 2. Cell migrering ut av intestinal eksplantering og innen kollagen-gel. Celler som migrerte av en intestinal eksplantering i løpet av 3 dagers inkubering i fravær eller nærvær av 10 ng / ml GDNF ble fiksert, farget med DAPI (blå) og fotografert i viser GFP-merket mage NCC (grønn). 70X forstørrelse. Målestokk: 100 mikrometer. Den stiplede linjen representerer omtrentlig størrelse og plassering av explant før det ble tatt av gelen. Klikk her for å se større figur. Klikk here for å se større bilde.

Figur 3. Kvantifisering av ente NCC migrasjon potensial. Antall og spredning (Feret diameter) av GFP-uttrykke celler migrerer ut av tarm eksplantater etter tre dager ble kvantifisert ved hjelp ImageJ programvare 17. I begge tilfeller er det en betydelig forskjell mellom ubehandlede og GDNF-behandlede tilstander i henhold til en Student t-test (p <0,001, *). Den gjennomsnittlige diameter av intestinal skiver (stiplet svart linje: 260 mikrometer) ble inkludert for å skille mellom passive inngang og aktiv migrasjon. Nt: ikke-behandlet, n:. Antall eksplantater bearbeidet Klikk her for å se større bilde .
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Vi viser hvordan vår ex vivo eksplantering kultur teknikken kan brukes til nøyaktig å kvantifisere enteric NCC migrering potensiale i nærvær av GDNF. En slik presis kvantifisering, gjøres meget lettere ved hjelp av 200 mikrometer tykke vibratome gut seksjoner i stedet for store stykker av omtrentlig størrelse, som tidligere beskrevet 11,12,15. Faktisk gjør dette at vi å arbeide med et rimelig antall celler i en reproduserbar innstilling. Til opplysning, tillater ensartet fordeling av fluorescerende enteriske NCC i kaudal-de region av tynntarmen fra hvilken eksplantering skiver skjæres også analyse av flere seksjoner fra ett enkelt gut (figur 1A). Videre, gitt at både mage NCC og axons kan avslutte vev i slike analyser 11, tilbaketrekking av explants på slutten av kultur periode lar oss fokusere utelukkende på trekkende NCC.
De fleste kritiske trinnene ble skissert i protokollen tekst, men som welfare av tarm explant er viktig å skaffe sunn migrerer NCC, særlig forsiktighet bør utvises. Unngå å utsette embryonale vev for plutselige temperaturendringer, spesielt når tarmen er innebygd i agarose (trinn 3,3). Pass på at tarmen er ved romtemperatur og agarose så kjølig som mulig (men likevel smeltet) for å unngå "matlaging" vevet. Tarm eksplantering skal trives på kulturmediet fylt collagen-gel, ofte øker i størrelse og søle ut av agarose skive. Hvis explant vises usunn eller verre, har en tendens til å gå til grunne under inkubasjon, prøve å erstatte den PBS med kultur media ved romtemperatur (f.eks HEPES-bufret M2 eller DMEM supplert med 10% FBS) for å hjelpe holde den oppe i disseksjon.
En viktig begrensning i vår tilnærming er at det er avhengig av tilgjengeligheten av en muselinje overdragelse et fluorescerende etiketten til migrering NCC. I fravær av en slik ressurs, et antistoff mot migrating NCC (for eksempel anti-Ret-eller anti-Sox10) kan brukes til å merke celler som invaderte collagen-gel. Videre, gitt at tarmen mikro-miljøet er langt mer komplisert enn en enkelt collagen-gel, resultatene som oppnås med dette in vitro-analysen kan ikke helt reflekterer oppførselen til enteriske NCC in vivo. Andre forsøk med live-cell imaging er anbefalt å vurdere denne atferden. Det er også verdt å merke seg at i tillegg til sin rolle som en kjemoattraktant, er GDNF kjent for å fremme spredning av trekkende enteric NCC fire. Vårt mål på enteriske NCC migrasjon potensiale i nærvær av GDNF er således sannsynligvis en blanding av sanne cellemigrering og celleproliferasjon, beslektet til de in vivo mekanismene som fører til NCC kolonisering av tarmen. Dersom et klart skille mellom disse to prosessene er ønskelig, kan tilsetningen av en cellecyklus-blokkerer (f.eks AZD 5438 18) i vekstmedium begrense analysen til celle migration.
Denne teknikken kan utvides til å teste forskjellige andre ekstracellulære ligander så vel som inhibitorer av spesifikke signalveier, og hvilken som helst kombinasjon deri. Andre vev kan også potensielt bli dissekert og seksjonert, slik at studiet av NCC migrasjon i mange embryonale strukturer. Kombinert med romanen og / eller uncharacterized mutant musestammer med mulige defekter i NCC utvikling, kan vår teknikk brukes til raskt å screene for mangler ved migrasjon atferd som svar på konkrete signale hendelser.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Forfatterne har ingenting å avsløre.
Acknowledgments
Vi takker Denis Flipo for bildebehandling og analyse råd, og David W. Silver i hvis laboratoriet Gata4p [5kb]-GFP mus linjen ble generert. Forskning i Pilon laboratoriet er finansiert av CIHR, NSERC, FRQS og FRQNT.
Materials
| Name | Company | Catalog Number | Comments |
| DMEM powder | Wisent | 219-010-XK | |
| NaHCO3 | Bioshop | SOB999 | Biotechnology grade |
| Steriflip vacuum filtration system (0.22 micron) | EMD Millipore | SCGP00525 | |
| Penicilin-Streptomycin solution, 100x | Wisent | 450-201-EL | |
| Fetal bovine serum | Wisent | 095-150 | High quality grade |
| Collagen I | BD biosciences | 354236 | |
| NaOH | Bioshop | SHY700 | Diluted from 10 N stock then sterile-filtered |
| GDNF | Cedarlane | CLCYT305 | |
| Falcon 24-well Plate | BD biosciences | 353047 | |
| Dissecting scissors | Fisher Scientific | 089515 | |
| Glass Petri dish | VWR | 89000-306 | |
| PBS | Sigma | P5493 | Cell culture grade |
| Dissecting microscope | Leica | M125 | |
| Dumont #5 Forceps | Fine Science Tools | 11251-20 | |
| Agarose | Bioshop | AGA001 | Biotechnology grade |
| Surgical blade | Feather | 21 | |
| All Purpose Instant Krazy Glue Pen | Krazy Glue | KG824 | |
| HM 650V Vibrating-Blade Microtome | Thermo Scientific | 920110 | |
| Paraformaldehyde | Sigma-Aldrich | P6148 | |
| DAPI | Sigma-Aldrich | D9564 | |
Table of specific reagents and equipment. |
|||
References
- Bronner, M. E., Le Douarin, N. M. Development and evolution of the neural crest: An overview. Dev. Biol. 366 (1), 2-9 (2012).
- Bergeron, K. F., Silversides, D. W., Pilon, N. The developmental genetics of Hirschsprung's disease. Clin. Genet. 83 (1), 15-22 (2013).
- Obermayr, F., Hotta, R., Enomoto, H., Young, H. M. Development and developmental disorders of the enteric nervous system. Nat. Rev. Gastroenterol. Hepatol. 10 (1), 43-57 (2012).
- Sasselli, V., Pachnis, V., Bursn, A. J. The enteric nervous system. Dev. Biol. 366 (1), 64-73 (2012).
- Sanchez, M. P., Silos-Santiago, I., et al. Renal agenesis and the absence of enteric neurons in mice lacking GDNF. Nature. 382 (6586), 70-73 (1996).
- Baynash, A. G., Hosoda, K., et al. Interaction of endothelin-3 with endothelin-B receptor is essential for development of epidermal melanocytes and enteric neurons. Cell. 79 (7), 1277-1285 (1994).
- Chalazonitis, A., Pham, T. D., et al. Neurotrophin-3 is required for the survival-differentiation of subsets of developing enteric neurons. J. Neurosci. 21 (15), 5620-5636 (2001).
- Goldstein, A. M., Brewer, K. C., Doyle, A. M., Nagy, N., Roberts, D. J. BMP signaling is necessary for neural crest cell migration and ganglion formation in the enteric nervous system. Mech. Dev. 122 (6), 821-833 (2005).
- Jiang, Y., Liu, M. T., Gershon, M. D. Netrins and DCC in the guidance of migrating neural crest-derived cells in the developing bowel and pancreas. Dev. Biol. 258 (2), 364-384 (2003).
- Ramalho-Santos, M., Melton, D. A., McMahon, A. P. Hedgehog signals regulate multiple aspects of gastrointestinal development. Development. 127 (12), 2763-2772 (2000).
- Natarajan, D., Marcos-Gutierrez, C., Pachnis, V., de Graaf, E. Requirement of signaling by receptor tyrosine kinase RET for the directed migration of enteric nervous system progenitor cells during mammalian embryogenesis. Development. 129 (22), 5151-5160 (2002).
- Young, H. M., Hearn, C. J., et al. GDNF Is a chemoattractant for enteric neural cells. Dev. Biol. 229 (2), 503-516 (2001).
- Amiel, J., Sproat-Emison, E., et al. Hirschsprung disease, associated syndromes and genetics: a review. J. Med. Genet. 45 (1), 1-14 (2008).
- Pilon, N., Raiwet, D., Viger, R. S., Silversides, D. W. Novel pre- and post-gastrulation expression of Gata4 within cells of the inner cell mass and migratory neural crest cells. Dev. Dyn. 237 (4), 1133-1143 (2008).
- Nagy, N., Goldstein, A. M. Endothelin-3 regulates neural crest cell proliferation and differentiation in the hindgut enteric nervous system. Dev. Biol. 293 (1), 203-217 (2006).
- Nagy, A., Gertsenstein, M., Vintersen, K., Behringer, R. Manipulating the mouse embryo: a laboratory manual. , 3rd ed, CSH Press. Cold Spring Harbor. 209-250 (2003).
- Schneider, C. A., Rasband, W. S., Eliceiri, K. W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods. 9, 671-675 (2012).
- Byth, K. F., Thomas, A., et al. AZD5438, a potent oral inhibitor of cyclin-dependent kinases 1, 2, and 9, leads to pharmacodynamic changes and potent antitumor effects in human tumor xenografts. Mol. Cancer Ther. 8 (7), 1856-1866 (2009).

{kind=link}