

Summary
我们提出了一个先体外后体内的细胞迁移测定法,允许在各种生长因子的存在下肠道神经嵴细胞迁移的潜在精确定量。
Abstract
神经嵴细胞(NCC)是一个短暂的和多能细胞群源于背神经管和广泛地在整个发展脊椎动物胚胎迁移。除了提供周边神经胶质细胞和神经元,NCC生成的黑素细胞,以及大部分的颅面骨骼。 NCC迁移和分化是由它们的轴向原点沿神经管和他们接触到不同的区域外线索的组合控制。细胞外配体的这种贡献是特别明显的肠神经系统(ENS),神经节的一个复杂的相互连接的网络,本地控制(其中包括)肠道肌肉运动和肠蠕动的形成过程中。在吻端到尾鳍时尚 - - 准肠道的整个长度大多数ENS是从华北克拉通,为了殖民承接了漫长的旅程一个小的初始池派生。其中已知的多条信号通路影响肠道定植NCC,GDNF / Ret信号被认为是最重要的。事实上,RET配体GDNF的时空方式控制分泌肠道间质,主要是负责吸引和RET表达肠NCC指导,并在胚胎肠道。在这里,我们描述的体外细胞迁移测定,利用转基因小鼠系具有荧光标记的NCC,这使得在各种生长因子,包括GDNF的存在肠溶NCC迁移潜在精确定量。
Introduction
神经嵴细胞(NCC)是胚胎发育过程中形成了许多衍生工具的瞬态细胞类型特有的脊椎动物。该细胞群出现在神经板,相邻的非神经外胚层1的边界。在神经胚形成,为神经板的地方NCC沿形成神经管背侧边缘弯曲。 NCC再经过上皮 - 间质转化,分离和迁移远离神经管。 NCC殖各个胚胎结构,包括消化道那里形成整个肠神经系统(ENS),神经节的互连网络中嵌入肠壁。由于最近检讨2,3,许多基因都参与了这个复杂结构的发展。
大多数ENS的是从迷走神经管( 即约准后脑/脊髓边界)一个小水池NCC原始的派生4。这些神经祖细胞到达前肠围绕胚胎一天(五)9.0小鼠,然后尾端迁移肠道间质内,直到约e15.0拓殖整个胚胎肠。结肠的神经祖细胞的未成年子集也由骶NCC,其侵入后肠道的相反方向到盲肠4提供。既迷走神经和骶NCC需要多次迁移,增殖,存活和分化,促进线索,以确保完全形成ENS的。在这方面,动物模型 - 尤其是转基因小鼠 - 是有助的在几个必要的细胞外配体的鉴定:GDNF(神经胶质细胞源性神经营养因子),内皮素-3,神经营养素-3,骨形态发生蛋白(骨形态发生蛋白),netrin诱导,以及声波和印度刺猬(Shh和Ihh信号)5-10。其中,GDNF通过酪氨酸激酶跨膜受体RET(转染过程中整理的)信号被确认为日Ë最关键的途径的吸引力和NCC的指导,并在胚胎肠道。 GDNF是由肠道间质细胞分泌,并形成了一个时空方式控制rosrrocaudal梯度是直接chemoattractive到肠道NCC,其表达RET 11,12。
除其他功能外,ENS通过与肠壁平滑肌相互作用调节消化道内运动。在肠道导致先天性巨结肠的末端区域神经节的缺席:受影响区段的强直性收缩导致堵塞,消化材料的上游积累和肠道和腹部的大规模扩张。先天性巨结肠症发生大约在5,000一只活产婴儿。肠道NCC的rostro-尾迁移模式被认为是主要的促进因素先天性巨结肠症的病因。从迁移NCC和b的最后部分的源结肠,最远owel被殖民统治,是最容易受到ENS中形成的缺陷。按照肠道NCC迁移的关键作用,GDNF / Ret信号的干扰是先天性巨结肠13的主要已知的遗传原因。
为了更好地研究华北克拉通和ENS的发展,我们产生了转基因小鼠线-命名Gata4p [5KB]-GFP 14 -在这种迁徙NCC都贴有绿色荧光蛋白(GFP)。我们接下来完善的体外细胞迁移测定中,由其它组11,12,15适于从发表的作品,即现在允许在各种生长因子,如GDNF的存在肠溶NCC迁移潜在精确定量。
Subscription Required. Please recommend JoVE to your librarian.
Protocol
伦理声明
在老鼠的实验进行了以下的在医学研究中使用动物的照顾和处理动物护理指引加拿大议会。涉及动物的操纵协议已获魁北克蒙特利尔大学的伦理委员会(科米特Institutionnel去保护DES ANIMAUX;编号0512-R3-650-0513)。
1。胶原蛋白凝胶的制备
在无菌的方式工作,在组织培养罩。
- 准备完整的5倍DMEM培养基(Eagle的修改必需培养基),包括标准的抗生素。溶解3.37克DMEM粉末和0.925克的NaHCO 3在20毫升水中。穿过一个0.22微米的过滤器进行消毒。添加2.5无菌100X青霉素/链霉素和25毫升无菌热灭活胎牛血清的溶液中。贮存于4℃。
- 冰,混合800微升的I型胶原溶液(3.77 mg / ml的0.02N的乙酸,过滤灭菌的),600微升的完全DMEM培养液5倍和17微升1N的NaOH。用无菌水稀释至3毫升的最终体积。包括在组合有关的生长因子。我们通常使用的GDNF在10毫微克/毫升,以刺激肠道NCC迁移。
- 存款约480μl,以单行的在24孔板中每个孔中。避免气泡。其余行可用于测试的其他生长因子对细胞迁移的影响。
- 让胶原蛋白在37℃下聚合至少1小时,在无菌恒温箱
2。 16动物的解剖
- 成立交配和检查阴道塞第二天早晨。一天E0.5是中午阴道塞被发现的当天,隔离女性和等待12天,直到E12.5。
- 麻醉孕鼠与异氟醚和二氧化碳吸入安乐死。
- 喷鼠标,用70%的乙醇。提起腹部s亲属,并与解剖剪刀打开腹部腔。
- 取出子宫到玻璃培养皿中充满了冰冷的PBS(磷酸盐缓冲盐水)。切子宫横向个人蜕膜肿胀之间的隔离每个胚胎着床部位。
- 每个植入位点在另一个玻璃培养皿充满冰冷的PBS分别工作。在解剖显微镜下,用细镊子取出子宫肌肉层。
- 打开卵黄囊和羊膜揭示胚胎。切断血管加盟胚胎的胎盘/卵黄囊的时候,因为他们是交织在一起的发展肠子照顾。
- 断绝胚胎的头颈部。
- 插入一个封闭镊子在腹腔胚胎的空腔,就在暗红色色肝上,并让钳子打开的本身(停止施加压力),以使一个横向开口在腹部空腔中。抠开镊子下来,对胚胎完全打开腹部的后端。
- 抢肝背后的结缔组织和拉的胆量了腹部,注意不要打破肠(大肠连接到肛门)。
- 切断结肠,从胚胎的其余部分释放的胆量。切割可以在沿着结肠的任何位置进行。保留尾部后。
- 梳理出从盲肠结缔组织,肠子然后休息。要小心,不要伤了肠子,而这样做。
- 切掉肝和胃(在小肠的喙末端),以及中肾和生殖嵴,如果一些都存在。
- 隔离小肠。再次,要小心不要夹到肠。从现在起,肠道组织的rosrrocaudal方向可以使用锋利的曲率目前在喙末端进行跟踪。留在小肠的PBS中于室温下尽可能短的时间尽可能烯前床上用品(见步骤3.3)。
- 最后,记录尾部体节的每个胚胎的数目准确地确定胚胎发育的阶段。
3。胚胎肠切片
- 在继续进行胚胎解剖,熔体1.5克琼脂糖于100ml PBS(磷酸盐缓冲盐水),并保持在50℃。
- 倒入琼脂糖融化成一个嵌入模具( 例如闭2 ml离心管,已被切割纵向切除约1/4的管壁)。让琼脂糖冷却至约42-45℃,它应该是只稍微温暖的触感。
- 就在琼脂糖固化之前嵌入胚胎肠(这可以用钳子提示进行评估,并会发生大约36-38℃)。保持由所述折叠喙端与钳肠道,通过沿着模具的长度琼脂糖拉得非常缓慢。这有助于保持肠道直而琼脂糖集。放开T只要它开始反抗移动的问题。跟踪肠道的喙,尾方向。
- 放于冰箱中2-3分钟的模具,以确保琼脂糖完全凝固。
- 取出琼脂糖从模具中(通过向外滑动打开的Eppendorf管中)。用刀片,取出多余的琼脂糖两端,使切口垂直于肠。
- 胶水肠/延髓结束琼脂糖挡了下来与振动叶片切片机的金属舞台。修剪对肠/琼脂糖块两侧的过量的琼脂糖。
- 安装在上述振动切片机室中的金属的阶段。如果有必要,调整阶段的角度,使肠道是尽可能垂直(因而垂直于叶片)。盖在冰冷的PBS试样。使切片机刀片向下到几毫米下的缓冲表面。
- 使200微米vibratome横向切口尾最小肠,确保该电子ACH琼脂糖片包含一个完整的肠道部分。
4。肠道外植体的培养
- 轻轻的存放新鲜切肠/琼脂糖片平放在胶原凝胶用钳子,把一片鸡犬中间好。
- 孵育3天37℃,在潮湿的5%CO 2的气氛中,以允许NCC的迁移出来的外植体。
- 取小肠/琼脂糖片落胶原凝胶轻轻地用钳子。小心不要损坏凝胶下方。
- 避免在为了随后的温育和洗涤步骤直接触摸胶原凝胶,以确保细胞迁移模式不受影响。修正了每孔1小时,在室温下500微升4%PFA(多聚甲醛)(在PBS)中。
- 更换固定液,用500μL的DAPI(4',6 - 二脒基-2 - 苯基吲哚)溶液(5μg/ ml的PBS中),每孔,孵育10分钟,在室温下进行。
- 每次洗好3倍用500μlPBS中5分钟。
- 拍摄嵌入在胶原凝胶内各孔的荧光细胞(GFP和DAPI通道)。
5。图像分析
我们广泛利用ImageJ的17来处理和量化外植体培养后生成的图像。
- 首先,成像微米幻灯片在相同的放大倍数为荧光细胞的照片。测量以像素为单位一微米(用直线工具)的长度。
- 设置比例(分析/设置刻度;像素/微米输入号码)。
- 对于每个GFP荧光照片,图像格式改变为8位灰度(Image/Type/8-bit)。
- 调整信号(图像/调整/亮度/对比度)的强度。
- 减去背景噪音,如果有必要(工艺/减去背景;调整滚球半径)。
- 设定阈值以突出细胞和细胞团块(图像/调整/阈值),则Ñ应用分水岭分割团块(工艺/二进制/流域)。
- 分析粒子指定感兴趣区域(ROI)的区域,并生成细胞数量的统计数据(分析/分析颗粒;集大小最小排除剩余的像素)。
- 设置测量选项包括费雷特直径(分析/设置测量)。
- 集团的投资回报,并测量作为一个整体来确定的Feret直径,指示细胞扩散(分析/工具/投资回报率经理/更多/ OR,则投资回报率经理/添加和ROI经理/测量)。
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
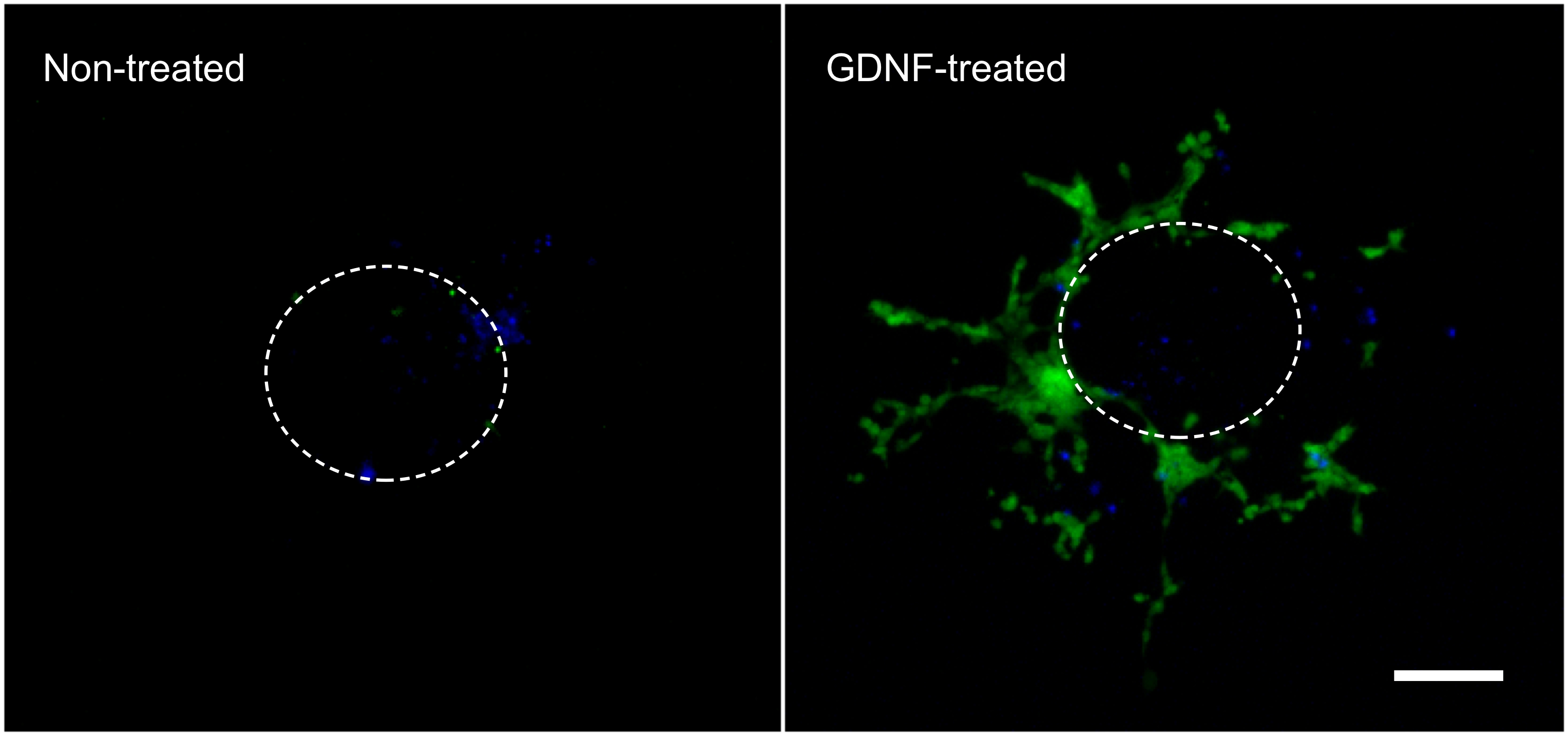
下面的结果是代表什么可以与这里描述的( 图1)的技术来获得的。使用生长因子( 即 GDNF)刺激的表达GFP的肠溶NCC出肠道外植体并进入胶原凝胶( 图2)的迁移。虽然一些小区出来的外植体在无生长因子,这些大多不是GFP标记和表示被动条目。有必要从胶原凝胶取出肠切片,以便记录的结果,因为这个组织仍然是人口稠密通过荧光细胞和本来隐藏的细胞趴在胶原底。这些结果量化显示,更多的细胞被发现在胶原凝胶中,当GDNF存在,而活动迁移发生( 图3)。的确,被动细胞被发现立即将外植下,肠切片的直径内,WHER EAS细胞,积极侵入胶原凝胶搬离起点和进一步蔓延。

图1。外植体培养技术。一)荧光肠NCC用于制造200μ厚的外植体尾部小肠区域内统一的人口概况 。比例尺:200μm的B)的含有胶原的培养基被沉积在24孔板中并使其硬化1小时。琼脂糖包埋胚胎肠Vibratome片沉积在凝胶上(每孔1片),和荧光肠溶NCC允许出来的外植体转移3天。该切片,然后取下成像的侵入胶原凝胶的细胞前。s/ftp_upload/50709/50709fig1highres.jpg“目标=”_blank“>点击这里查看大图。

图2。细胞迁移出肠道外植体,并在胶原蛋白凝胶中。细胞迁移出的肠道外植体3天的孵育在10纳克/毫升的GDNF的存在或不存在时进行固定,用DAPI染色(蓝色),拍摄到显示GFP标记的肠道NCC(绿色)。 70X的放大倍率。比例尺:100微米。虚线表示的近似大小和之前它被带下凝胶外植体的位置。 点击这里查看大图。 点击ħ埃雷查看大图。

图3。表达GFP的细胞后第3天迁移出肠道外植体的肠道NCC迁移潜力量化的数量和传播(费雷特直径)使用ImageJ软件17定量。在这两种情况下,存在未处理的和GDNF处理的情况之间的差异显著根据学生t检验(P <0.001,*)。肠片(黑色虚线:260微米)的平均直径被列入被动进入主动与移民之间的区别。 NT:非治疗中,n:处理外植体的数目点此查看大图 。
Subscription Required. Please recommend JoVE to your librarian.
Discussion
我们展示了如何我们的体外外植体培养技术可以用来精确地量化肠溶NCC迁移潜力的GDNF的存在。这种精确的定量是通过使用200微米厚的vibratome肠组织切片而不是大块的近似大小,如先前所描述11,12,15大大便利。的确,这使得我们能与细胞中一种高度可重现设定合理数量的工作。值得注意的是,荧光肠溶NCC的小肠从该外植体条带被切断的尾最区域内的均匀分布还允许从一个单一的肠道( 图1A)的多个部分的分析。此外,鉴于双方肠NCC和轴突可以退出该组织这样的实验11,停药外植体在培养期结束时让我们专注于迁徙NCC。
最关键的步骤在协议文本进行了概述,但由于在W肠道外植体elfare是极为重要的获得健康的迁移NCC,尤其应注意。避免将胚胎组织,以突然的温度变化,尤其是当肠道被嵌入在琼脂糖(步骤3.3)。确保肠道是在室温和琼脂糖尽可能凉爽(但仍然熔化),以避免“烹调”的组织。肠道外植体应蓬勃发展的培养基填充胶原蛋白凝胶,往往增加的大小和蔓延的琼脂糖片。如果外植体出现不健康或更糟的是,往往孵化期间要灭亡,尝试在室温下用培养基代替PBS( 如 HEPES缓冲M2或添加有10%FBS),以帮助剥离时维持它。
一个主要限制我们的做法是,它依赖于鼠标线赋予荧光标记迁移到NCC的可用性。在没有这样的资源,对米的抗体igrating NCC( 例如抗Ret或抗SOX10)可以被用于标记细胞侵入胶原凝胶中。此外,由于肠道微环境是远远超过了一个简单的胶原凝胶更加复杂,这在体外测定法中获得的结果可能并不完全反映肠溶NCC 在体内的行为。涉及活细胞成像实验的其他建议,以评估这种行为。还值得注意的是,除了其作为趋化作用,GDNF是已知的促进迁移肠溶NCC 4的增殖。我们的GDNF的存在肠溶NCC迁移潜力的措施是这样大概的混合真实细胞迁移和细胞增殖的,类似于在体内导致肠道NCC定植机制。如果这两个过程之间有着明显的区别是理想,又多了一个细胞周期阻断剂( 如 AZD 5438 18)在培养基可以限制分析细胞migratioÑ。
该技术可以扩展到测试各种其他细胞外配体以及特定信号传导途径的抑制剂,并在其中的任何组合。其他组织也有可能被解剖和切片,让NCC移民在许多胚胎结构的研究。结合新颖的和/或未知突变小鼠品系与NCC发展可能存在的缺陷,本技术可被应用到响应于特定的信号事件快速筛选在迁移行为缺陷。
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
作者什么都没有透露。
Acknowledgments
我们感谢丹尼斯FLIPO用于图像处理和分析意见,戴维W在银鱼的实验室生成的Gata4p [5KB]-GFP的小鼠品系。研究在实验室皮隆是由CIHR,NSERC,FRQS和FRQNT资助。
Materials
| Name | Company | Catalog Number | Comments |
| DMEM powder | Wisent | 219-010-XK | |
| NaHCO3 | Bioshop | SOB999 | Biotechnology grade |
| Steriflip vacuum filtration system (0.22 micron) | EMD Millipore | SCGP00525 | |
| Penicilin-Streptomycin solution, 100x | Wisent | 450-201-EL | |
| Fetal bovine serum | Wisent | 095-150 | High quality grade |
| Collagen I | BD biosciences | 354236 | |
| NaOH | Bioshop | SHY700 | Diluted from 10 N stock then sterile-filtered |
| GDNF | Cedarlane | CLCYT305 | |
| Falcon 24-well Plate | BD biosciences | 353047 | |
| Dissecting scissors | Fisher Scientific | 089515 | |
| Glass Petri dish | VWR | 89000-306 | |
| PBS | Sigma | P5493 | Cell culture grade |
| Dissecting microscope | Leica | M125 | |
| Dumont #5 Forceps | Fine Science Tools | 11251-20 | |
| Agarose | Bioshop | AGA001 | Biotechnology grade |
| Surgical blade | Feather | 21 | |
| All Purpose Instant Krazy Glue Pen | Krazy Glue | KG824 | |
| HM 650V Vibrating-Blade Microtome | Thermo Scientific | 920110 | |
| Paraformaldehyde | Sigma-Aldrich | P6148 | |
| DAPI | Sigma-Aldrich | D9564 | |
Table of specific reagents and equipment. |
|||
References
- Bronner, M. E., Le Douarin, N. M. Development and evolution of the neural crest: An overview. Dev. Biol. 366 (1), 2-9 (2012).
- Bergeron, K. F., Silversides, D. W., Pilon, N. The developmental genetics of Hirschsprung's disease. Clin. Genet. 83 (1), 15-22 (2013).
- Obermayr, F., Hotta, R., Enomoto, H., Young, H. M. Development and developmental disorders of the enteric nervous system. Nat. Rev. Gastroenterol. Hepatol. 10 (1), 43-57 (2012).
- Sasselli, V., Pachnis, V., Bursn, A. J. The enteric nervous system. Dev. Biol. 366 (1), 64-73 (2012).
- Sanchez, M. P., Silos-Santiago, I., et al. Renal agenesis and the absence of enteric neurons in mice lacking GDNF. Nature. 382 (6586), 70-73 (1996).
- Baynash, A. G., Hosoda, K., et al. Interaction of endothelin-3 with endothelin-B receptor is essential for development of epidermal melanocytes and enteric neurons. Cell. 79 (7), 1277-1285 (1994).
- Chalazonitis, A., Pham, T. D., et al. Neurotrophin-3 is required for the survival-differentiation of subsets of developing enteric neurons. J. Neurosci. 21 (15), 5620-5636 (2001).
- Goldstein, A. M., Brewer, K. C., Doyle, A. M., Nagy, N., Roberts, D. J. BMP signaling is necessary for neural crest cell migration and ganglion formation in the enteric nervous system. Mech. Dev. 122 (6), 821-833 (2005).
- Jiang, Y., Liu, M. T., Gershon, M. D. Netrins and DCC in the guidance of migrating neural crest-derived cells in the developing bowel and pancreas. Dev. Biol. 258 (2), 364-384 (2003).
- Ramalho-Santos, M., Melton, D. A., McMahon, A. P. Hedgehog signals regulate multiple aspects of gastrointestinal development. Development. 127 (12), 2763-2772 (2000).
- Natarajan, D., Marcos-Gutierrez, C., Pachnis, V., de Graaf, E. Requirement of signaling by receptor tyrosine kinase RET for the directed migration of enteric nervous system progenitor cells during mammalian embryogenesis. Development. 129 (22), 5151-5160 (2002).
- Young, H. M., Hearn, C. J., et al. GDNF Is a chemoattractant for enteric neural cells. Dev. Biol. 229 (2), 503-516 (2001).
- Amiel, J., Sproat-Emison, E., et al. Hirschsprung disease, associated syndromes and genetics: a review. J. Med. Genet. 45 (1), 1-14 (2008).
- Pilon, N., Raiwet, D., Viger, R. S., Silversides, D. W. Novel pre- and post-gastrulation expression of Gata4 within cells of the inner cell mass and migratory neural crest cells. Dev. Dyn. 237 (4), 1133-1143 (2008).
- Nagy, N., Goldstein, A. M. Endothelin-3 regulates neural crest cell proliferation and differentiation in the hindgut enteric nervous system. Dev. Biol. 293 (1), 203-217 (2006).
- Nagy, A., Gertsenstein, M., Vintersen, K., Behringer, R. Manipulating the mouse embryo: a laboratory manual. , 3rd ed, CSH Press. Cold Spring Harbor. 209-250 (2003).
- Schneider, C. A., Rasband, W. S., Eliceiri, K. W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods. 9, 671-675 (2012).
- Byth, K. F., Thomas, A., et al. AZD5438, a potent oral inhibitor of cyclin-dependent kinases 1, 2, and 9, leads to pharmacodynamic changes and potent antitumor effects in human tumor xenografts. Mol. Cancer Ther. 8 (7), 1856-1866 (2009).

{kind=link}