

Summary
우리는 다양한 성장 인자의 존재 하에서 장용성 신경 능선 세포 이동 전위의 정확한 정량을 허용 생체 세포 이동 분석을 제시한다.
Abstract
신경 능선 세포 (NCC)는 등의 신경관에서 유래 및 개발 척추 동물의 배아에 걸쳐 광범위하게 마이그레이션 과도 다 능성 세포 집단이다. 말초 뉴런 및 아교를 제공 할뿐만 아니라, NCC는 멜라닌 세포뿐만 아니라 부교감 얼굴 골격의 대부분을 생성한다. NCC 마이그레이션 및 차별화 신경 튜브 및 지역에 별개의 세포 외 신호에 대한 노출을 따라 자신의 축 원점의 조합에 의해 제어된다. 세포 외 리간드의 이러한 기여는 장용 신경계 (ENS), 복잡한 상호 연결 (다른 것들 사이에) 로컬로 제어하는 신경 중추의 네트워크 창자 근육 운동과 장 운동의 형성시에 특히 분명하다. 꼬리 패션에 주동이의 - - 미래의 창자의 전체 길이를 ENS의 대부분은 정착하기 위해 긴 여행을 수행 NCC의 작은 초기 수영장에서 파생됩니다. 로 알려진 몇 가지 신호 전달 경로 중장내 NCC의 식민지에 영향을 미치는, GDNF / RET 신호가 가장 중요한 것으로 인식되고 있습니다. 사실, 창자의 중간 엽으로 RET 리간드 GDNF의 시공간으로 제어 분비 배아 창자 및 내 매력과 RET - 표현 장내 NCC의 안내를 주로 담당하고 있습니다. 여기, 우리는 GDNF 등 다양한 성장 인자의 존재에 장내 NCC 마이그레이션 가능성 정확한 정량화 할 수 찬란 NCC를 가진 유전자 변형 마우스 라인을 사용하고, 생체 세포 이동 분석에 대해 설명합니다.
Introduction
신경 능선 세포 (NCC)의 배아를 개발하는 동안 많은 유도체를 형성하는 척추 동물에 고유 한 과도 세포 유형입니다. 이 세포 집단이 아닌 신경 외배엽 1에 인접한 신경 판의 경계에서 발생한다. neurulation 동안, 성형 신경관의 등쪽 가장자리를 따라 신경 판의 장소 NCC의 굽힘. NCC는 분리하는 멀리 신경 튜브에서 마이그레이션, 상피 - 간엽 전환을 받고있다. NCC들은 전체 장용 신경계 (ENS)를 형성 소화관 장벽에 내장 신경 신경절의 상호 연결된 네트워크를 포함하는 다양한 배아 구조를 정착. 최근 2,3 리뷰, 많은 유전자는이 복잡한 구조의 개발에 참여하고있다.
ENS의 대부분은 (즉, 미래의 후뇌 / 척수 경계 정도) 미주 신경 튜브에서 NCC의 원래의 작은 수영장에서 파생 4.이 신경 전구 세포는 쥐의 배아 일 (E) 9.0 주위에 foregut에 도달 약 e15.0 전체 배아 창자를 식민지 때까지 용기의 중간 엽에서 꼬리 쪽 마이그레이션 할 수 있습니다. 대장의 신경 전구 세포의 작은 부분 집합은 최대 맹장 4 반대 방향으로 후방 창자 침입 성례 NCC에 의해 제공됩니다. 미주와 성례의 두 NCC가 필요 여러 마이그레이션, 핵 확산, ENS의 완전한 형성을 보장하기 위해 생존과 분화 촉진 단서.을 이와 관련, 동물 모델 - 특히 유전자 변형 생쥐는 - 여러 가지 중요한 세포 외 리간드의 식별 수단이되었습니다 GDNF (아교 세포 유래 신경 영양 인자), 엔도 텔린-3, 뉴로-3, BMP에 (뼈 형태 형성 단백질), 네트 린에게 뿐만 아니라, 음과 인도의 고슴도치 (쉬와 IHH) ~ 10. 이들 중, (형질 전환시 재 배열) 티로신 키나제 횡단 수용체 RET을 통해 신호 GDNF는 번째로 인식되고배아 창자 및 내 매력과 NCC의 지침에 대한 전자의 가장 중요한 통로. GDNF는 창자의 중간 엽에서 분비 및 RET (11, 12)을 표현 장내 NCC, 직접 chemoattractive 인 시공간으로 제어 rosrrocaudal 구배를 형성한다.
다른 기능 사이에, ENS는 장 벽에 부드러운 근육과의 상호 작용을 통해 소화 기관 내에서의 움직임을 조절한다. Hirschsprung의 질병에있는 장 결과의 말단 부위에있는 신경 중추의 부재 : 영향을받는 부분의 토닉 수축 막힘, 소화 물질의 상류 축적과 창자와 복부의 거대한 팽창에 이르게한다. Hirschsprung 병은 약 5,000 정상 출산을 발생합니다. 장내 NCC의 rostro - 꼬리 이주 패턴은 Hirschsprung의 질병의 원인에 대한 주요 기여 요인이 될 것으로 생각됩니다. NCC와 B의 마지막 부분을 마이그레이션하는 소스의 결장, 먼정착 할 owel는 ENS 형성의 결함에 가장 민감하다. 장내 NCC 마이그레이션에서의 중요한 역할에 따라, GDNF / RET 신호의 중단 Hirschsprung의 질병 (13)의 주요 알려진 유전 적 원인이다.
[5킬로바이트-GFP 14 Gata4p 이름 - - 철새 NCC는 녹색 형광 단백질 (GFP)으로 표시되는 더 나은 NCC와 ENS 개발을 연구하기 위해, 우리는 유전자 변형 마우스 라인을 생성합니다. 우리는 다음에 지금과 같은 GDNF 등의 다양한 성장 인자의 존재에 장내 NCC 마이그레이션 가능성의 정확한 정량화 할 수 있습니다 다른 그룹 11,12,15에서 출판 작업에서 적응 생체 세포 이동 분석을, 완성.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
윤리 문
마우스를 포함하는 실험은 의학 연구에 사용 된 동물의 관리 및 조작을위한 동물 보호 지침의 캐나다 협의회 다음을 수행 하였다. 동물의 조작을 포함하는 프로토콜은 몬트리올 퀘벡 대학의 기관 윤리위원회에 의해 승인되었습니다 (위원회 (CCT) Institutionnel 드 보호 데 Animaux, 참조 번호 0512-R3-650-0513).
1. 콜라겐 젤의 제조
조직 문화 후드에서 멸균 방식으로 작업 할 수 있습니다.
- 표준 항생제 등을 포함한 5 배 DMEM (이글의 수정 필수 매체)를 준비합니다. 20 ㎖의 물에 DMEM 분말과 탄산 수소 나트륨의 0.925 g의 3.37 g을 녹인다. 0.22 μm의 필터를 통해 전달하여 소독. 무균 100X 페니실린 / 스트렙토 마이신 및 멸균 열 불 활성화 된 소 태아 혈청 25 ㎖의 2.5 ML을 추가합니다. 4 ℃에서 보관
- 얼음에서 800 μl를 혼합완전한 배 DMEM 600 ㎕의 1 N NaOH를 17 ㎕의 콜라겐 I 용액 (0.02 N 아세트산, 필터 소독에 3.77 ㎎ / ㎖). 3 ㎖의 최종 부피로 멸균 수로 희석. 믹스 관련 성장 인자를 포함합니다. 우리는 일반적으로 장내 NCC 마이그레이션을 자극하는 10 NG / ㎖에서 GDNF를 사용합니다.
- 24 - 웰 플레이트의 단일 행의 각 웰 예금 480 μL. 거품을 피하십시오. 나머지 행은 세포 이동의 다른 성장 인자의 효과를 테스트하기 위해 사용될 수있다.
- 콜라겐은 37 ℃에서 멸균 배양기에서 1 시간 이상을 중합하자
2. 동물 (16)의 해부
- 교배를 설정하고 다음 날 아침에 질 플러그를 확인합니다. 일 E0.5는 질 플러그가 발견되는 일 정오되는 여성을 격리하며 E12.5 12 시까 지 일을 기다려야.
- 이소 플루 란으로 임신 한 쥐를 마취 및 CO 2 흡입에 의해 안락사.
- 70 % 에탄올로 마우스 스프레이. 복부의 리프트친척 및 해부 가위로 복부에 구멍을 엽니 다.
- 얼음처럼 차가운 PBS (인산염 완충 식염수)로 채워진 유리 페트리 접시에 자궁을 제거합니다. 각각의 배아 주입 사이트를 분리하기 위해 가로 방향으로 개별 deciduum의 부종과 자궁을 잘라.
- 얼음처럼 차가운 PBS로 가득 다른 유리 페트리 접시에 따로 따로 각 주입 사이트에서 작동합니다. 해부 현미경 하에서 자궁 근육층을 제거하는 미세 집게를 사용한다.
- 배아를 나타 내기 위해 내장 난황과 양막을 엽니 다. 그들이 개발 창자 고리로 연결되었다는로, 태반 / 내장 난황에 배아를 결합하는 혈관을 절단하는 경우주의하십시오.
- 목에서 배아의 머리를 절단.
- 그냥 어두운 붉은 색, 간 이상, 태아의 복강에 닫힌 집게를 삽입하고 복부 구멍에 횡단 구멍을 만들기 위해 (압력을 적용 중지) 자체의 집게가 열려하자. 향해 열린 집게를 아래로 당겨완전히 복부를 열 수있는 배아의 뒤쪽 끝.
- 간 뒤에 결합 조직을 잡고 (콜론 항문에 부착) 내장을 파괴하지 않도록주의하고, 복부 밖으로 내장을 빼냅니다.
- 배아의 나머지 부분에서 용기를 무료로 결장을 잘라. 컷 콜론의 임의의 위치에서 할 수있다. 이후의 꼬리 부분을 예약.
- 맹장의 결합 조직, 내장의 다음 나머지를 애타게. 그렇게하는 동안 장내 상처하지 않도록주의하십시오.
- 약간의 위험이있는 경우에는 간, 위 (소장의 주동이의 끝)뿐만 아니라 mesonephros 성기 능선을 버려야.
- 소장을 분리합니다. 또, 대장을 잘라 내지 않도록주의하십시오. 지금부터, 창자 조직의 rosrrocaudal 방향은 주동이의 끝에 존재하는 날카로운 곡선을 사용하여 추적 할 수 있습니다. EM 전에 가능한 한 짧은 시간 동안 실온에서 PBS에 소장 남기기침구 (단계 3.3 참조).
- 마지막으로, 정확하게 배아 발달의 단계를 결정하기 위해 각각의 배아에 대한 꼬리 somites의 수를 기록한다.
3. 배아 창자의 단면
- 배아 해부 진행하기 전에 100 ㎖의 PBS (인산염 완충 식염수)에 1.5 g 아가로 오스를 녹여 50 ℃로 유지
- 부어 삽입 금형에 용융 아가로 오스 (예를 들어, 약 1 / 4 튜브 벽의 절제하는 길이 절단 된 2 ML의 microcentrifuge 관을 폐쇄). 아가로 오스는 약 42 ~ 45 ° C의 열을 식히, 그것은 터치에 약간 따뜻해야한다.
- 다만 아가로 오스 응고하기 전에 배아 소장을 포함 (이것은 집게 팁으로 평가 될 수 있으며, 주변에 36 ~ 38 ° C 발생)을. 접힌 주동이의 말에 집게로 장을 들고, 금형의 길이를 따라 아가로 오스를 통해 매우 천천히 잡아 당깁니다. 이것은 바로 아가 세트 동안 창자를 유지하는 데 도움이됩니다. T 릴리스즉시 이동하고 저항하기 시작 실행합니다. 창자의 주동이의 - 꼬리 방향을 확인합니다.
- 아가로 오스가 완전히 설정 한 보장하기 위해, 냉장고 2-3분에 금형을 넣어.
- (열린 에펜 도르프 튜브 밖으로 밀어) 금형에서 아가로 오스를 꺼내. 블레이드, 소장에 수직 커트를하고, 양쪽 끝에서 초과 아가로 오스를 꺼내.
- 진동 블레이드와 마이크로톰의 금속 무대에서 아래로 차단 아가 소장 /의 주동이의 끝을 붙입니다. 소장 / 아가로 오스 블록의 측면에 여분의 아가로 오스를 낸다.
- 진동 마이크로톰 챔버의 금속 단계를 탑재합니다. 필요에 따라서는 소장 (따라서 블레이드에 수직) 가능한 한 수직 스테이지의 각도를 조정한다. 얼음처럼 차가운 PBS로 표본을 커버. 버퍼 표면에서 몇 mm까지 마이크로톰 블레이드를 가져옵니다.
- 해당 전자를 보장 꼬리 가장 소장의 200 ㎛ vibratome 가로 컷을 만드는ACH 아가로 오스 조각은 전체 장 섹션이 포함되어 있습니다.
4. 장내 외식 문화
- 부드럽게 잘 각각의 중앙을 향해 한 조각을 배치, 집게로 콜라겐 젤에 갓 잘라 소장 / 아가로 오스 조각 평면을 예금.
- 이식편 중 NCC의 마이그레이션을 할 수 있도록, 습도 5 % CO 2 분위기에서, 37 ° C 3 일간 배양한다.
- 집게 매우 부드럽게 콜라겐 겔 떨어져 소장 / 아가로 오스의 조각을 가져 가라. 아래의 젤이 손상되지 않도록주의하십시오.
- 세포의 이동 패턴이 교란되지 않도록하기 위해 후속 배양 및 세척 단계에서 직접 콜라겐 겔을 만지지 마십시오. 실온에서 잘 1 시간 당 (PBS에서) 4 % PFA (파라 포름 알데히드)의 500 μL로 수정합니다.
- 물론 당 DAPI 500 μL (4 ', 6 - diamidino -2 - 페닐 인돌) 솔루션 (PBS에서 5 ㎍ / ml)로 정착을 교체하고 실온에서 10 분 알을 품다.
- 잘 배 각각의 세척5 분 동안 PBS 500 μL와.
- 각 웰 내의 콜라겐 겔에 포함 된 형광 세포 (GFP 및 DAPI 채널)의 사진.
5. 이미지 분석
우리는 이식편 배양 한 후 생성 된 이미지를 처리하고 정량화 ImageJ에 17를 광범위하게 사용했다.
- 형광 세포 사진과 같은 배율에서 마이크로 미터 슬라이드 이미징에 의해 시작합니다. (직선 도구를 사용하여) 픽셀 1 미크론의 길이를 측정한다.
- (/ 설정 규모를 분석, 픽셀 / 마이크론의 입력 번호) 규모를 설정합니다.
- 각 GFP의 형광 사진을 위해, 8 비트 그레이 스케일 (Image/Type/8-bit)로 이미지 형식을 변경합니다.
- 신호 (이미지 / 조정 / 밝기 / 명암 대비)의 강도를 조정합니다.
- 필요 (프로세스 / 배경 빼기, 롤링 볼 반경을 조정)하면 배경 잡음을 뺍니다.
- 세포와 세포 덩어리 (이미지 / / 임계 값을 조정), 강조하기 위해 임계 값을 설정합니다N 덩어리 (공정 / 진 / 유역을) 분할하는 분수령을 적용합니다.
- 관심 영역 (ROI)을 지정하고 세포 수의 통계를 생성하는 입자를 분석 (입자 분석 / 분석, 설정 크기 최소 남아있는 픽셀을 제외).
- 의 Feret의 직경 (설정 / 측정 분석)를 포함하는 측정 옵션을 설정합니다.
- 그룹의 ROI와의 Feret의 직경을 결정하기 위해 전체로 측정, 세포의 확산의 표시 (/ 도구 / ROI 관리자 / 더 / 또는 다음 ROI 관리자 추가 / ROI 관리자 / 측정 분석).
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
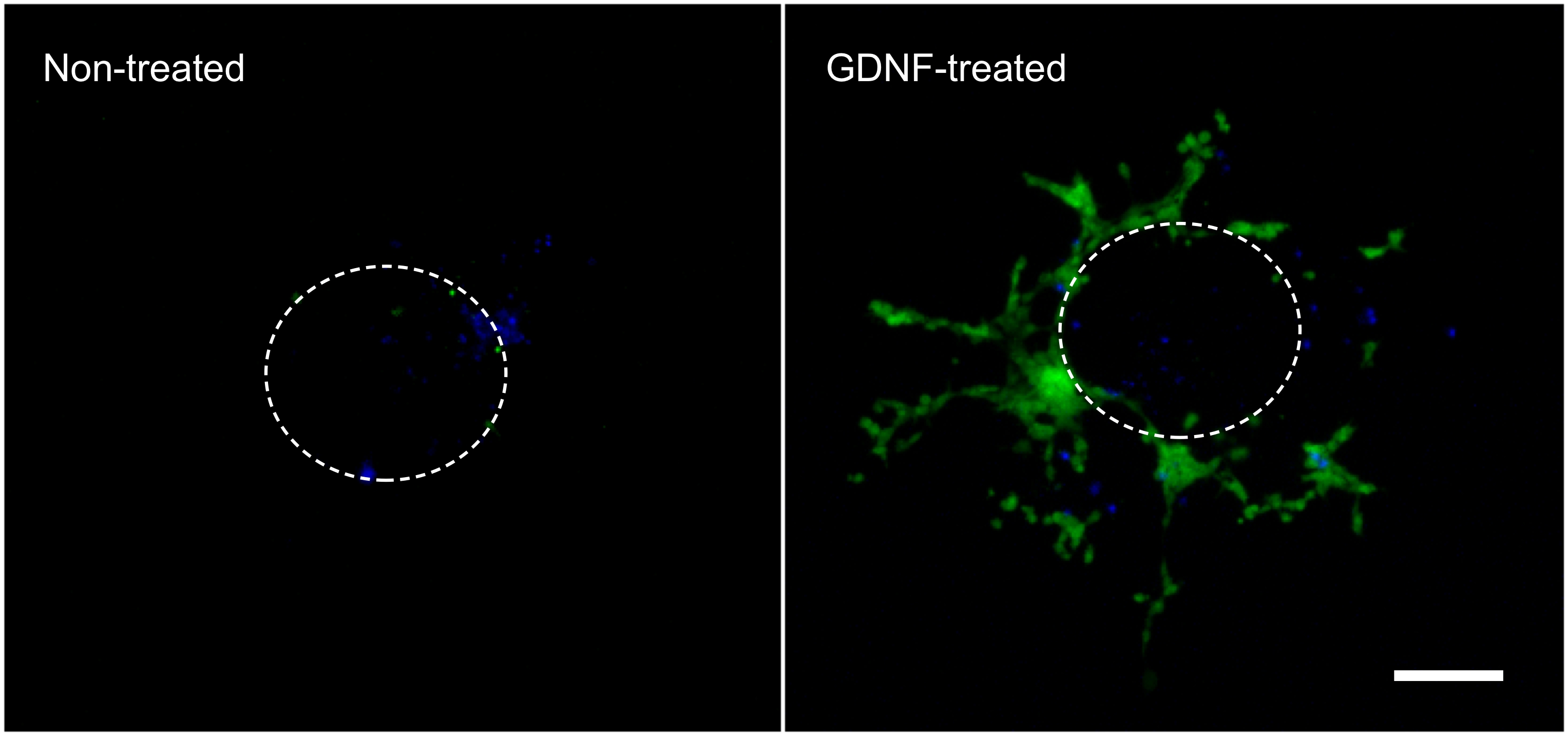
다음과 같은 결과 (그림 1) 여기에 설명 된 기술로 얻을 수있는 대표입니다. 성장 인자 (즉, GDNF)의 사용은 GFP를 발현하는 장 이식편에서 그리고 콜라겐 겔에 장내 NCC (그림 2)의 이동을 자극한다. 일부 세포가 성장 인자의 부재하에 이식편 밖으로 나와 있지만, 이들은 주로 GFP-표지 아니며 패시브 엔트리를 나타낸다. 그것은이 조직이 여전히 크게 형광 세포에 의해 채워지며, 그렇지 않으면 아래 콜라겐 거짓말 세포를 숨기는 것 같은 결과를 기록하기 위해 콜라겐 겔에서 장내 조각을 제거 할 필요가있다. 이러한 결과의 정량화는 GDNF가있을 때 더 많은 세포가 콜라겐 겔 내에서 발견되는 것을 보여줍니다, 그리고 활동적인 이동은 (그림 3) 일어난다. 실제로, 수동 세포는 장 슬라이스의 직경 내에서, 어디 있나, 바로 외식에서 찾을 수 있습니다 적극적으로 콜라겐 겔에 침입 EAS 세포가 떨어져 원래의 자신의 관점에서 이동 및 추가 확산.

그림 1. 이식편 문화 기술.) 200 μ 두께의 절편을 만드는 데 사용되는 꼬리 소장 영역 내에서 형광 장내 NCC의 통일 인구의 개요. 스케일 바 :. 200 ㎛ B) 콜라겐을 포함하는 배양 배지를 24 - 웰 플레이트에 입금하고 1 시간 동안 강하게 남아있다. 아가로 오스 - 임베디드 배아 내장의 Vibratome 슬라이스 젤 (물론 당 하나의 조각)에 부착되고, 형광 장내 NCC는 3 일 동안 외식 밖으로 마이그레이션 할 수 있습니다. 슬라이스 한 후 콜라겐 젤을 침공 세포 이미징 전에 이륙합니다.s/ftp_upload/50709/50709fig1highres.jpg "대상 ="_blank "> 큰 이미지를 보려면 여기를 클릭하십시오.

그림 2. 10 NG / ML GDNF의 부재 또는 존재에서 3 일 배양시 장 이식편 밖으로 이주. 세포 장 이식편에서 그리고 콜라겐 겔 내에서 세포의 이동은, 고정 DAPI (파란색)로 염색하고 촬영 한 GFP - 라벨 장내 NCC (녹색)을 보여줍니다. 70X 배율. 스케일 바 : 100 μm의. 점선은 대략적인 크기와는 젤을 찍은 전에 이식편의 위치를 나타냅니다. 큰 그림을 보려면 여기를 클릭하십시오. 시간을 클릭더 큰 이미지를 볼 수 감수.

그림 3. 3 일 후에 장 외식 밖으로 이주 GFP - 표현 세포의 장내 NCC 마이그레이션 가능성의 정량화. 수와 스프레드 (의 Feret의 직경) ImageJ에 소프트웨어 (17)를 사용하여 정량 하였다. 두 경우 모두,이 치료와 GDNF 처리 조건 사이에 유의 한 차이는 학생의 T 테스트에 따라입니다 (P <0.001 *). 장 조각 (검은 점선 : 260 μm의)의 평균 직경은 수동 입력 및 활성 마이그레이션을 구별하기 위해 포함되었다. NT : 비 처리, N :. 처리 외식의 수 더 큰 이미지를 보려면 여기를 클릭하십시오 .
Subscription Required. Please recommend JoVE to your librarian.
Discussion
우리는 우리의 생체 이식편 배양 기술은 정확하게 GDNF의 존재에 장내 NCC 이주 가능성을 정량화하는 데 사용할 수있는 방법을 보여줍니다. 이러한 정확한 정량이 크게 11,12,15 이전에 설명 된대로, 200 ㎛ 두께의 vibratome 창자 부분을 사용하는 대신 대략적인 크기의 큰 조각에 의해 촉진된다. 사실, 이것은 우리가 재현성이 높은 환경에서 세포의 적절한 수와 함께 작업 할 수 있습니다. 참고로, 이식편 슬라이스가 절단되는 소장의 꼬리 - 대부분의 영역 내에서 형광 장내 NCC의 균일 한 분포는 하나의 창자 (그림 1A)에서 여러 부분의 분석을 할 수 있습니다. 또한, 장내 NCC와 축삭이 모두 문화 기간 말에 같은 분석 (11), 외식의 철수에 조직을 종료 할 수 있습니다 우리가 철새 NCC에 독점적으로 초점을 맞출 수 있다는 없습니다.
가장 중요한 단계는하지만 w로서, 프로토콜 텍스트에 요약 하였다장 이식편의 elfare 건강 마이그레이션 NCC를 특히주의가되어야한다 얻기에 매우 중요합니다. 소장이 아가로 오스 (3.3 단계)에 포함되어 특히, 급격한 온도 변화에 배아 조직을 노출시키지 마십시오. 소장은 조직을 "요리"피할 수 (아무런 여전히 용융)와 같은 시원한 실내 온도와 아가로 오스에 있는지 확인합니다. 장 이식편은 종종 크기가 증가하고, 아가 로스 슬라이스 흘리, 배지 가득 콜라겐 겔에 번성한다. 이식편이 건강에 해로운 또는 악화가 나타나면, 배양 중에 멸망 해부하는 동안 그것을 유지하기 위해 (예를 들면 HEPES 버퍼 M2 또는 DMEM 10 % FBS와 보충) 실온에서 배양 배지와 PBS를 교체하려고하는 경향이있다.
우리의 접근 방식에 주요 한계는 NCC를 마이그레이션에 형광 라벨을 부여 마우스 라인의 가용성에 의존하고 있다는 점이다. 이러한 리소스의 부재에서 m에 대한 항체NCC를 igrating (예를 들어 안티 폐지 또는 방지 Sox10)는 콜라겐 겔에 침입 세포에 라벨을 사용할 수 있습니다. 또한,이 시험 관내 분석으로 얻어진 결과는 완전히 생체 내에서 장내 NCC의 행동을 반영하지 않을 수 있습니다, 장내 미세 환경이 훨씬 더 복잡 간단한 콜라겐 겔보다 주어진. 라이브 세포 이미징을 포함하는 추가 실험은이 동작을 평가하는 것이 좋습니다. 그것은 화학 유인 물질로서의 역할뿐만 아니라, GDNF는 장내 NCC 4 마이그레이션의 증식을 촉진하는 것으로 알려져있다 또한 주목할 만하다. GDNF의 존재 장내 NCC 마이그레이션 잠재적 인 우리의 조치는 그래서 가장 내장의 NCC의 식민지로 이어지는 생체 메커니즘에 가깝다 진정한 세포 이동 및 세포 증식의 혼합이다. 이러한 두 개의 프로세스 사이의 명확한 구별이 요구되는 경우, 배양 배지에서 세포주기 차단제 (예 AZD 5438 (18))의 첨가는 세포 migratio로 분석을 제한 할N.
이 기술은 다양한 세포 외 리간드뿐만 아니라 특정 신호 전달 경로의 억제제, 및 그 내부에 임의의 조합을 테스트하기 위해 확장 될 수있다. 다른 조직은 잠재적으로 해부 많은 배아 구조의 NCC 이주의 연구를 허용, 단면 할 수 있습니다. 소설 및 / 또는 NCC 개발 가능한 결함에 uncharacterized 돌연변이 마우스의 변종과 결합하여, 우리의 기술은 신속하게 특정 신호 이벤트에 대한 응답으로 마이그레이션 동작의 결함에 대한 화면으로 적용 할 수 있습니다.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
저자가 공개하는 게 없다.
Acknowledgments
우리는 이미지 처리 및 분석 조언 데니스 워드 컨트리의 .. 감사합니다, 데이비드 W. Silversides 그의 실험실에서 Gata4p [5킬로바이트-GFP 마우스 라인이 생성되었습니다. 필론 실험실에서 연구를 CIHR, NSERC, FRQS 및 FRQNT에 의해 투자된다.
Materials
| Name | Company | Catalog Number | Comments |
| DMEM powder | Wisent | 219-010-XK | |
| NaHCO3 | Bioshop | SOB999 | Biotechnology grade |
| Steriflip vacuum filtration system (0.22 micron) | EMD Millipore | SCGP00525 | |
| Penicilin-Streptomycin solution, 100x | Wisent | 450-201-EL | |
| Fetal bovine serum | Wisent | 095-150 | High quality grade |
| Collagen I | BD biosciences | 354236 | |
| NaOH | Bioshop | SHY700 | Diluted from 10 N stock then sterile-filtered |
| GDNF | Cedarlane | CLCYT305 | |
| Falcon 24-well Plate | BD biosciences | 353047 | |
| Dissecting scissors | Fisher Scientific | 089515 | |
| Glass Petri dish | VWR | 89000-306 | |
| PBS | Sigma | P5493 | Cell culture grade |
| Dissecting microscope | Leica | M125 | |
| Dumont #5 Forceps | Fine Science Tools | 11251-20 | |
| Agarose | Bioshop | AGA001 | Biotechnology grade |
| Surgical blade | Feather | 21 | |
| All Purpose Instant Krazy Glue Pen | Krazy Glue | KG824 | |
| HM 650V Vibrating-Blade Microtome | Thermo Scientific | 920110 | |
| Paraformaldehyde | Sigma-Aldrich | P6148 | |
| DAPI | Sigma-Aldrich | D9564 | |
Table of specific reagents and equipment. |
|||
References
- Bronner, M. E., Le Douarin, N. M. Development and evolution of the neural crest: An overview. Dev. Biol. 366 (1), 2-9 (2012).
- Bergeron, K. F., Silversides, D. W., Pilon, N. The developmental genetics of Hirschsprung's disease. Clin. Genet. 83 (1), 15-22 (2013).
- Obermayr, F., Hotta, R., Enomoto, H., Young, H. M. Development and developmental disorders of the enteric nervous system. Nat. Rev. Gastroenterol. Hepatol. 10 (1), 43-57 (2012).
- Sasselli, V., Pachnis, V., Bursn, A. J. The enteric nervous system. Dev. Biol. 366 (1), 64-73 (2012).
- Sanchez, M. P., Silos-Santiago, I., et al. Renal agenesis and the absence of enteric neurons in mice lacking GDNF. Nature. 382 (6586), 70-73 (1996).
- Baynash, A. G., Hosoda, K., et al. Interaction of endothelin-3 with endothelin-B receptor is essential for development of epidermal melanocytes and enteric neurons. Cell. 79 (7), 1277-1285 (1994).
- Chalazonitis, A., Pham, T. D., et al. Neurotrophin-3 is required for the survival-differentiation of subsets of developing enteric neurons. J. Neurosci. 21 (15), 5620-5636 (2001).
- Goldstein, A. M., Brewer, K. C., Doyle, A. M., Nagy, N., Roberts, D. J. BMP signaling is necessary for neural crest cell migration and ganglion formation in the enteric nervous system. Mech. Dev. 122 (6), 821-833 (2005).
- Jiang, Y., Liu, M. T., Gershon, M. D. Netrins and DCC in the guidance of migrating neural crest-derived cells in the developing bowel and pancreas. Dev. Biol. 258 (2), 364-384 (2003).
- Ramalho-Santos, M., Melton, D. A., McMahon, A. P. Hedgehog signals regulate multiple aspects of gastrointestinal development. Development. 127 (12), 2763-2772 (2000).
- Natarajan, D., Marcos-Gutierrez, C., Pachnis, V., de Graaf, E. Requirement of signaling by receptor tyrosine kinase RET for the directed migration of enteric nervous system progenitor cells during mammalian embryogenesis. Development. 129 (22), 5151-5160 (2002).
- Young, H. M., Hearn, C. J., et al. GDNF Is a chemoattractant for enteric neural cells. Dev. Biol. 229 (2), 503-516 (2001).
- Amiel, J., Sproat-Emison, E., et al. Hirschsprung disease, associated syndromes and genetics: a review. J. Med. Genet. 45 (1), 1-14 (2008).
- Pilon, N., Raiwet, D., Viger, R. S., Silversides, D. W. Novel pre- and post-gastrulation expression of Gata4 within cells of the inner cell mass and migratory neural crest cells. Dev. Dyn. 237 (4), 1133-1143 (2008).
- Nagy, N., Goldstein, A. M. Endothelin-3 regulates neural crest cell proliferation and differentiation in the hindgut enteric nervous system. Dev. Biol. 293 (1), 203-217 (2006).
- Nagy, A., Gertsenstein, M., Vintersen, K., Behringer, R. Manipulating the mouse embryo: a laboratory manual. , 3rd ed, CSH Press. Cold Spring Harbor. 209-250 (2003).
- Schneider, C. A., Rasband, W. S., Eliceiri, K. W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods. 9, 671-675 (2012).
- Byth, K. F., Thomas, A., et al. AZD5438, a potent oral inhibitor of cyclin-dependent kinases 1, 2, and 9, leads to pharmacodynamic changes and potent antitumor effects in human tumor xenografts. Mol. Cancer Ther. 8 (7), 1856-1866 (2009).

{kind=link}