Our labeling chamber is 1.2 m x 2.4 m x 3.6 m in size and holds 40,15 L pots (Figure 1). The computerized IRGA control system maintained CO2 concentrations between our set values of 360 and 400 ppm during the photosynthetically active period of the day (Figure 2a). The low CO2 alarm feature on the IRGA triggered solenoid valves to allow CO2 from the 13C enriched and natural abundance tanks into the chamber when the concentration dropped below the minimum threshold (e.g. 360 ppm). The dead band feature stopped the flow when the concentration reached the upper set point (e.g. 400 ppm). The iSeries temperature and humidity monitoring system connected to the air conditioner and dehumidifier held climate conditions within the set parameters throughout the growing season (Figure 2b). We used a one-ton (3.5 kW) air conditioning unit to keep the chamber cool.
The remote monitoring system allowed the logged data to be viewed at any time by a standard web browser. The CO2 concentrations, temperature and humidity values were down sampled by the web application to display graphs over the past 24-240 hr, in 24 hr increments. This created a quick visual to confirm that the daily fluctuations were within the expected limits. Viewing the web interface also showed the current chamber status, as well as provided alerts to potential problems such as not receiving recent data. At any time the complete dataset could also be downloaded from the web interface.
We measured photosynthetically active radiation (PAR) in the immediate interior and exterior of the chamber at four points with and without the lights on in the middle of the summer and the middle of the day using a quantum sensor. The PAR in the chamber was 31.5% lower than the exterior when the chamber lights were off and 22% lower than the exterior when the lights were on. Thus, the chamber lights help to significantly increase PAR penetration within the chamber by 9.5% (P<0.05).
Our continuous labeling system was able to produce 2759 g of A. gerardii biomass, 37% of which was aboveground biomass and 63% of which was belowground biomass. We achieved a 4.4 atom%13C whole plant label in our uniform plant material by setting the solenoid valves on the two CO2 tanks accordingly (Figure 1, Table 1). We achieved a 6.7 atom%15N whole plant label in our uniform plant material by mixing 98 atom%15N-KNO3 with 0.37 atom%15N-KNO3 in the KNO3 subsolution of a modified Hoagland's solution23 (Table 1). We watered the A. gerardii weekly with 750 ml total fluid (water plus Hoagland's solution) throughout the growing season. We fertilized with 200-500 ml of 15N labeled Hoagland's solution per week depending on plant productivity.
We utilized the hot water extraction method to determine if there were isotopic differences between the uniform and differentially labeled plant material. For the differentially labeled plants, upon harvest we removed any leaves that were completely dead and handled these separately as they were likely not differentially labeled. When looking at 13C content, all four incorporation days were significantly different from each other for the whole plant and the hot water extract, but for the hot water residue day 14 and 22 were not significantly different from each other (Table 1). When comparing the plant tissue fractions within each day, the hot water extract and residue were significantly different from each other for all four days and by day 22 the whole plant, extract, and residue were all significantly different from each other (Table 1). For the 15N incorporation into plant components, there were differences between days of incorporation and plant tissue fractions. For the hot water extract all four of the incorporation days were significantly different from each other for 15N, and for the whole plant and the hot water residue the shorter days of incorporation were significantly different than the longer days of incorporation (Table 1). The plant tissue fractions in the uniform plants were not significantly different from each other in 15N, but the hot water extract and residue were significantly different from each other in 15N for the differentially labeled litter.
All isotopic values are reported using the atom percent (atom %) notation (Equation 1), which is a more accurate notation than % to use at high levels of heavy isotope enrichment21. For example:
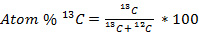 (1)
(1)
For this study, we ran statistical analyses using SAS version 9.2. We tested differences between the chamber interior and exterior light levels using a paired t-test. We tested differences between 13C and 15N labeling of hot water extracts and hot water residues using one-way analysis of variance (ANOVA) in PROC ANOVA. We used Duncan's multiple range test for multiple comparisons analysis. Significance was accepted at a P- level of 0.05. We used a Wilcoxon rank sum test to test that the data met the assumptions of the analysis.
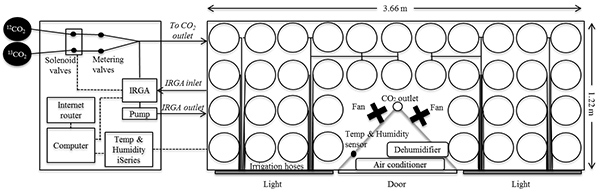
Figure 1. Schematic diagram of the 40 pot capacity continuous multi-isotope labeling chamber from a bird's eye view. Dotted lines represent electrical wiring, while solid lines represent gas or water tubing. Click here to view larger image.
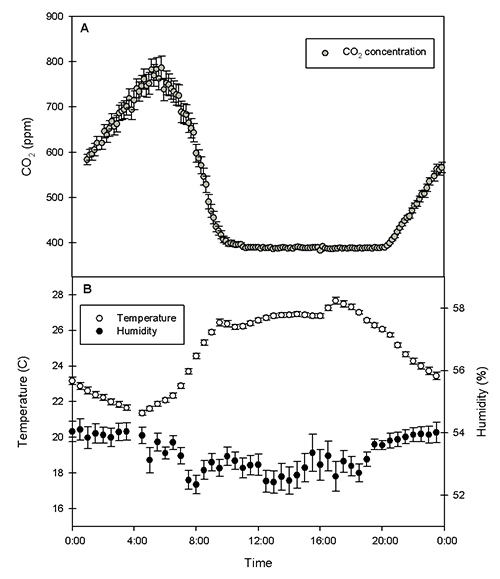
Figure 2. A) Average CO2 concentration (ppm) (+/- SE) over a twenty-four hour period for an entire growing season. B) Average temperature (ºC), open circles, and humidity (%), closed circles (+/- SE) over a twenty-four hour period for an entire growing season.
Click here to view larger image.
| Uniform (0) | Differential (7) | Differential (14) | Differential (22) | ||
| Whole Litter | 13C Atom % | 4.46±0.02 Aab | 3.93±0.05 Ba | 3.64±0.03 Ca | 3.35±0.06 Db |
| 15N Atom % | 6.69±0.07 Aa | 6.72±0.01 Aa | 6.33±0.06 Ba | 6.41±0.07 Ba | |
| Hot Water Extract | 13C Atom % | 4.59±0.04 Aa | 3.35±0.06 Bb | 2.79±0.06 Cb | 2.37±0.03 Dc |
| 15N Atom % | 6.69±0.03 Aa | 6.43±0.01 Bb | 5.89±0.07 Db | 6.16±0.05 Cb | |
| Hot Water Residue | 13C Atom % | 4.37±0.06 Ab | 4.1±0.03 Ba | 3.79±0.10 Ca | 3.66±0.05 Ca |
| 15N Atom % | 6.57±0.04 Ba | 6.71±0.02 Aa | 6.45±0.02 Ca | 6.44±0.03 Ca |
Table 1. Isotopic composition and litter chemistry for uniform and differentially labeled litter. Days of differential labeling outside the chamber are in parentheses. Comparisons between days of incorporation are in capital letters (across rows) and between litter fractions are in lower case letters (down columns) for each variable.
