Learning curve:
Mastery of a motor skill often requires persistent practice over time. A typical averaged learning curve is composed of two phases: an initial acquisition phase during which the success rate improves progressively, and a later consolidation phase when the success rate reaches the plateau (Figure 2C). It should be noted the learning curves of individual mice vary; different mice take different numbers of days to reach the plateau level, and the individual learning curves are usually not as smooth as the average one. Another way to present the mouse's improvement of reaching skills is the speed of success, which reflects the overall motor performance by taking accounts of the reaching speed as well as the accuracy. In general, the speed of success keeps improving after the mouse reaches its plateau success rate (Figure 2D).
In addition to individual variations of performance, some mice fail to learn the task (Figure 2E). These "nonlearners" usually make initial attempts to reach for the seeds, but for unknown reasons perform poorly despite repetitive training. They generally lose interest in reaching for the seeds after continuously failed attempts and stop reaching after 6-8 days. By contrast, some mice are over-shaped (Figure 2E). These over-shaped mice usually start with a very high success rate (>40%), and do not make significant improvement (i.e. 15% increase in success rate compared to first day of training) in motor performance with continued training. Their learning curves stay relatively flat, or may even decrease with continued training.
Motor memory:
Our previous data have suggested that once a motor skilled movement is mastered through repetitive practice, it can be stored as a form of motor memory and further practices are not required for its maintenance19. This motor memory can be measured by stopping the training after mice have reached plateau level of success rates, and retest their performance after an extended resting period (e.g. a few months). Retrained mice usually start with comparable success rates as have observed at the end of earlier training, and maintain high success rate in subsequent days. By contrast, age-matched naïve mice usually start with significantly lower success rates and progressively improve their performance with practice19 (Figure 2F). While food deprivation (2 days) before commencement of retraining is required, reshaping the previously trained mice is not necessary.
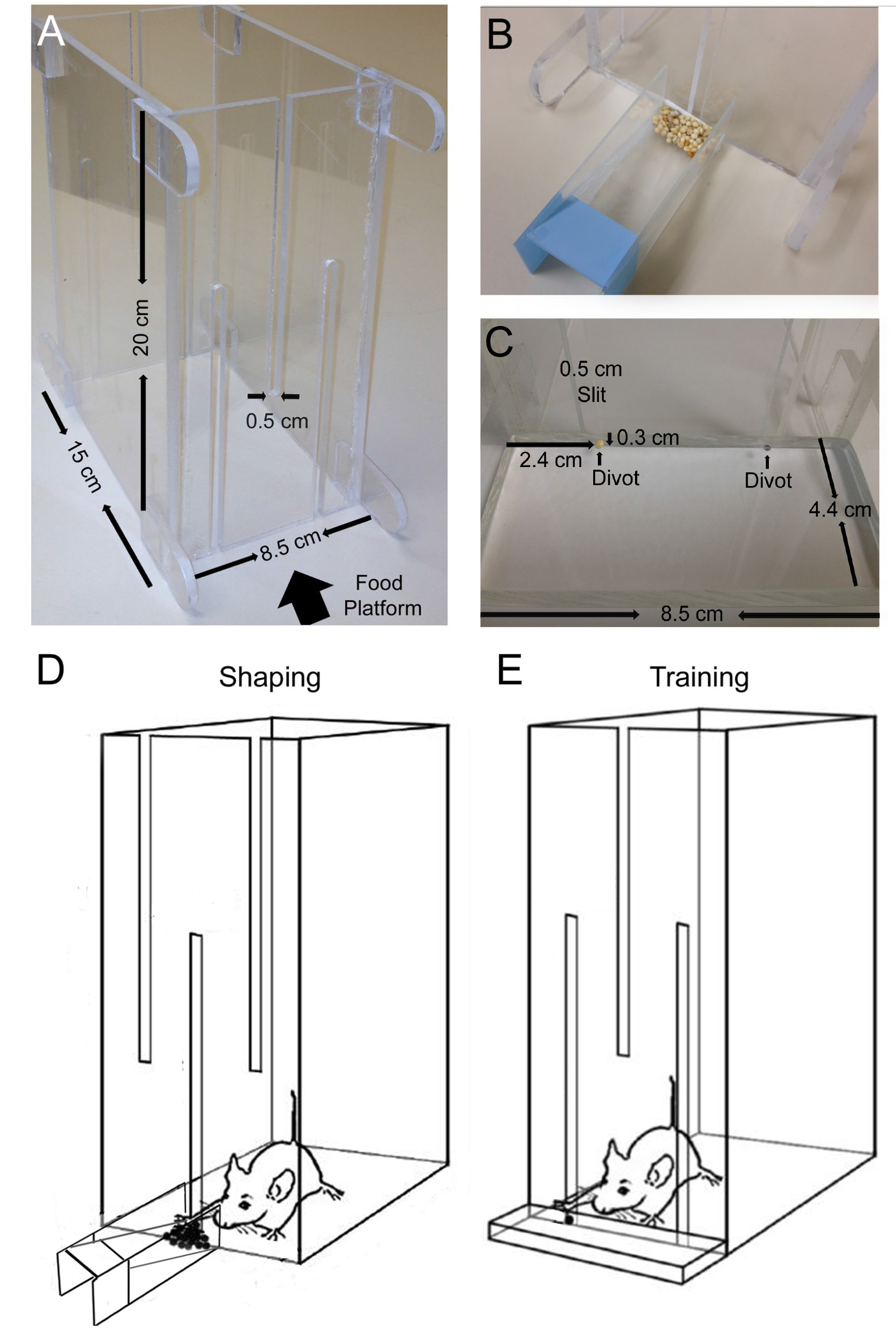
Figure 1. Design of the mouse training chamber. A. A photograph of the training chamber, with dimensions indicated. B. A photograph of the shaping seed tray, made from three slides glued together and placed in front of the shaping chamber. A pile of millet seeds are placed at the trough of the seed tray. C. A photograph of the food tray for training, placed in front of the training chamber. One millet seed is placed on the divot (pointed by arrows) that corresponds to right forelimb training. Dimensions and the location of the divot are indicated in the photograph. D. A cartoon drawing of the shaping chamber. The single-slit side of the training chamber is placed facing downwards. Seeds are placed in front of the middle slit and mice can use both paws to plow the seeds. E. A cartoon drawing of the training chamber. The double-slit side of the training chamber is placed facing downwards. One seed is placed on the food tray in front of the chamber slit corresponding to the preferred limb (in this case, the right forelimb). (D and E, modified from Xu et al.19). Click here to view larger image.

Figure 2. Representative results of mouse single-pellet reaching tasks. A. A general timeline of the experimental design. B. An example of bodyweight loss for a single mouse during food deprivation (F), shaping (S), and training (T). C. Average success rates improve over time during training (n=39). D. Average speed of success from the same group of mice presented in C. E. Success rates during training for an over-shaped mouse and a nonlearning mouse. F. Motor performance of pretrained (n=14) and naïve (n=10) adult mice during 4 day training (modified from Xu et al.19). All data are presented as mean ± s.e.m., ***, P<0.001. Click here to view larger image.
