Production Protocol
The protocol can be divided into 3 phases corresponding to major changes in culture media composition: (a) purification of endothelial cells from the microvessels, (b) proliferation, and (c) differentiation on filters (12-well polyethylene plates with a porosity of 1 μm) in co-culture with astrocytes. The different steps of the production protocol were assigned to specific days of the week in order to increase the reproducibility of the primary cell cultures (Figure 1). On Wednesday of the first week, hence 9 days before the establishment of the co-culture system, the microvessels were isolated (Figure 2A). To purify endothelial cells, the cultures were kept in the presence of decreasing concentrations of puromycin for 5 days. Most contaminating cells (mainly pericytes) were eliminated and the growth of endothelial cells was slower without modification of their phenotype. Spindle-shaped cells grow out of the capillary fragments isolated from rat brain grey matter (Figure 2B). On Monday of the second week, rat astrocytes were plated on the bottom of 12-well plates. Wednesday, RBEC were added to the luminal compartment of the filters. High density seeding at 160 x 103 cells/cm2 (Figure 2C) helps to avoid gaps at the periphery of the filter (Figure 2D) and generates a homogeneous differentiation of the endothelial cell monolayer. The cells show the typical elongated spindle-shaped morphology, align longitudinally, and form a uniform monolayer. Thursday, astrocyte culture medium was changed for the differentiation medium conditioned with the previous co-culture media (Figure 2E). Friday, RBEC on filters were grown in culture medium containing 500 nM hydrocortisone and the filters were transferred to 12-well plates containing the astrocytes. On the third week, experiments were performed between Monday and Wednesday.
RBEC Characterization and Determining Elements of Monolayer Permeability
The RBEC were characterized by immunostaining of endothelial and BBB markers as well as by measurements of TEER, permeability of LY and functionality of efflux transporters such as P-gp.
RBEC obtained by the puromycin purification method (Figure 1 and 2) grew in non-overlapping continuous monolayers that exhibited growth inhibition at confluence and displayed tightly apposed, fusiform and spindle shaped morphology. RBEC cultures expressed typical markers of endothelial cells: they showed positive immunostaining for platelet endothelial cell adhesion molecule (Figure 3A; CD31/PECAM) and expression of von Willebrand factor (Figure 3B). Without puromycin treatment, RBECs are quickly invaded by pericytes detected by desmin immunostaining (Figure 3C). Glial fibrillary acidic protein (GFAP) expressed by astrocytes (Figure 3D) is an important differentiation marker of astrocytes. With high passage number, astrocytes lose their ability to induce differentiation of the endothelial cells. For this reason, astrocyte cultures were standardized for their time of culture and only underwent one passage.
Culturing RBEC with media supplemented with hydrocortisone, followed by co-culture with astrocytes and with conditioned medium from the lower compartment of previous co-cultures led to strong induction of interendothelial TJs in comparison to RBEC cultured alone. The cells expressed claudin-5, ZO-1 and occludin proteins, which were localized at the cell-cell junctions, as shown by immunofluorescence (Figure 4B-D) and revealed by western blot analysis for ZO-1 and occludin proteins (Figure 4E). The morphological distribution at cell-to-cell contacts in a zipper-like configuration reflects (a) the tightness of the monolayer, (b) the cerebral origin of the capillary endothelial cells, and (c) is characteristic of highly differentiated cerebral endothelial cells. Paracellular permeability of the endothelial layer was monitored by measuring the TEER and the rate of influx of LY (457 Da) from the upper to the lower compartment of the filters. TEER was measured using an ENDOHM-12 for 12 mm culture cups connected to an EVOM voltohmmeter. The TEER of brain endothelial monolayers, indicating the tightness of TJs reached 300 ohm · cm2 on average (filters of 12 well plates, data not shown). The Pe for LY (Pe(LY), used as an integrity control of the barrier and co-incubated with tested compounds) reached an average of 0.26 ± 0.11 x 10-3 cm/min (Figure 4F) over a 1 year period.
In order to induce inflammation of RBEC we used the pro-inflammatory cytokine TNF-α at 5 ng/ml for 24 hr, and followed the inflammatory response by measuring CCL2 (MCP-1) secretion by RBEC in the upper compartment of the culture system which doubled from 120 ng/ml (non-treated control) to 250 ng/ml (Figure 5A). TNF-α treatment at 5 ng/ml or 50 ng/ml for 24 hr also opened the BBB as revealed by Pe(LY) measurement (Figure 5B) .
P-gp was visualized in differentiated RBEC by immunocytochemistry (Figure 6A). P-gp localization is known to be highly polarized and expressed essentially in the apical membrane of endothelial cells44 while the Glutamate transporter-1 (GLT-1) is mainly found in the basolateral fraction45. To assess the extent of polarization of our cultured RBEC, apical and basolateral proteins were separated from plasma membrane preparations using sucrose density gradients46. Western blot analysis was performed to assess expression levels of P-gp and GLT-1 in plasma, apical and basal membrane preparations. Microvessels freshly extracted from rat brain were used as the closest positive controls to the in vivo distribution of these proteins (Figure 6B). In both microvessels and differentiated RBEC membrane preparations, P-gp was expressed as a band of 170 kDa, mainly localized in the F1 apical membrane fraction (Figure 6B, C). GLT-1 expression was indeed expressed in the F3 basal membrane fraction of microvessels as a band of 65-70 kDa, but was not detected in the cultured RBEC.
In parallel experiments, the functional activity of P-gp in the endothelial cell monolayer was tested using rhodamine 123 (R123) as a ligand. The abluminal to luminal (B to A) efflux of R123 was 1.7 times higher than in the opposite direction (Figure 6D). To prove P-gp involvement, we used specific P-gp inhibitors, verapamil (25 µM, 30 min preincubation) and cyclosporine A (1 µM, 30 min preincubation). With verapamil and cyclosporine treatment, R123 accumulation in RBEC was increased 1.6 fold and 2 fold respectively, compared to untreated control (Figure 6E).
Receptors Involved in Receptor Mediated Transcytosis (RMT) Mechanisms
The model was further characterized for expression of different receptors potentially involved in transcytosis mechanisms at the BBB, such as the LRP1, LDLR or TfR (Figure 7A).
Functional transport studies were performed with the ligands for TfR and LDLR. Live RBEC were incubated with rat transferrin labelled with CY3 (Tf-Cy3) for 180 min at 37 °C (Figure 7B, C). Quantification of Tf-Cy3 uptake and transport in the rat in vitro BBB model showed that the slope of the curves slightly decreased beyond 2,400 picomoles (Figure 7B), suggesting that binding/uptake was saturable. Moreover, the saturation of the binding/uptake was correlated with the accumulation of Tf in the lower compartment. To confirm this observation, Tf-Cy3 was incubated at 1,200 picomoles (ascending part of the curve) with an excess of non-fluorescent Tf at 4,800 picomoles (plateau/saturation) (Figure 7C). These experiments showed a decrease of Tf-Cy3 accumulation in the lower compartment and confirmed a saturable mechanism typical of ligand-receptor interaction in our in vitro BBB model.
Similarly, LDLR functionality was demonstrated by the uptake and transport of its ligand DiILDL across the endothelial cell monolayer (Figure 7D, E). Live RBEC were incubated with DiILDL for 30 min at 37 °C. The binding/uptake was not correlated with the accumulation of DiILDL in the lower compartment (Figure 7D). Quantification of DiILDL transport showed that the slope of the curves decreased beyond 2 µg, suggesting that transport was saturable and receptor-related in our model. Sodium azide was added at 0.05%, 15 min prior to DiILDL incubation (Figure 7E). The absence of toxicity of sodium azide incubation at 0.05% was assessed by Pe(LY) measurement (data not shown). These experiments showed decreased DiILDL accumulation in the lower compartment. Overall, our results suggest RMT for DiILDL in our in vitro BBB model.
Other In Vitro BBB Models based on Endothelial Cells from Rat spinal Cord or Mouse Brain
Enzymatic digestion with a mix of collagenases / dispase is one of the major parameters to control in terms of cell viability and production yield of cerebral microvessels. The concentration of enzyme and more important the ratio between the quantity of enzyme relative to tissue, needs to be calibrated precisely between rat brain, rat spinal cord and mice brain. Microvessel seeding density and number of endothelial cells to reach 90% confluence (9 days post microvessel seeding) were linked and are important parameters to improve cell growth, limit the number of population doubling and the quality of the models. Rat spinal cord microvessels were produced with the same protocol as the one described herein for rat brain microvessels (in terms of tissue amount, 2 spinal cords corresponding approximately to 1 brain) to obtain the same quantity of cells per T75 flask within the same time scale. Meninges from 5-week-old C57Bl6 mice brain were difficult to eliminate because they stick to the cortex. A brief treatment of the brain with dispase appears to facilitate removal of the meninges. The important parameters that contribute to the production of reproducible endothelial cell monolayers from rat brain, rat spinal cord and mouse brain are highlighted and summarized in Figure 8.

Figure 1. Flow chart summarizing the main steps of the in vitro BBB model preparation. The different steps of the production protocol were assigned to specific days of the week in order to increase the reproducibility of the primary endothelial cell cultures. The protocol starts on Wednesday of the first week, with microvessel production from 5-week-old Wistar rats.

Figure 2. Phase contrast photomicrographs of RBEC, astrocytes and the co-culture system. (A) 5-week old Wistar rat microvessel fragments at the time of plating out. (B) Three day old RBEC cultures treated for 2 days with the P-gp substrate puromycin at 4 µg/ml. (C) Pure confluent endothelial monolayers at day 1 after plating on the Millipore filters of a 12 well plate coated with collagen type IV and fibronectin. (D) Homogeneity of the cell monolayer at the periphery of the insert filter. (E) Confluent astrocyte culture on the day of establishment of the co-culture showing characterized honeycomb morphology. (F) Scheme representing the co-culture system with localization of the photomicrographs in C, D and E.
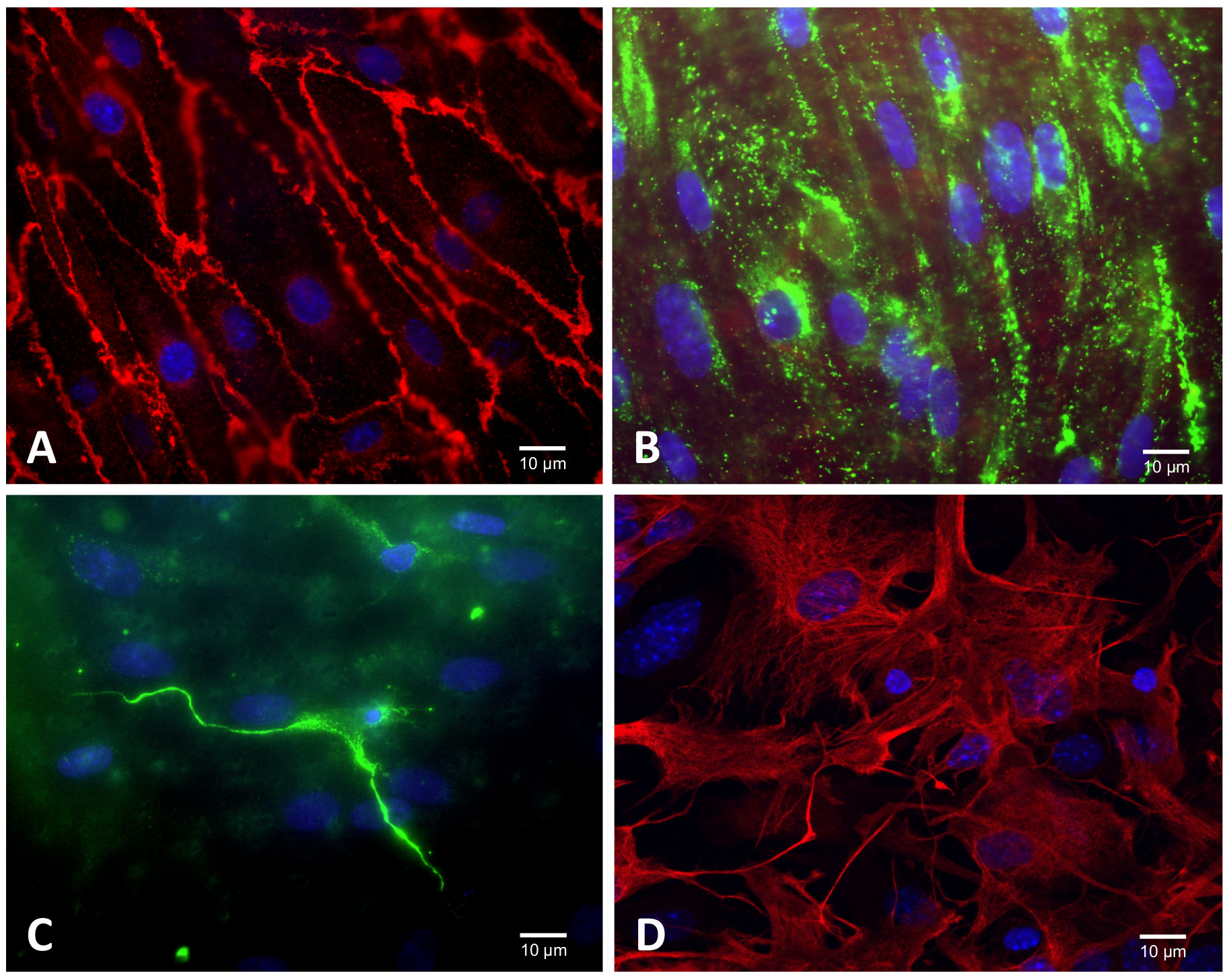
Figure 3. Characterization of RBEC cultures by immunofluorescence microscopy. (A) Brain endothelial cells express platelet endothelial cell adhesion molecule PECAM/CD31 at the cell border, (B) von Willebrand factor (VWF) shows a relatively punctate staining, brighter and more aggregated in some cells than others, and concentrated in the perinuclear zone. (C) Without puromycin treatment of RBEC cultures, pericytes are detected with a positive immunostaining for desmin (in green) and have characteristic small round nuclei. (D) Astrocytes express glial fibrillary acidic protein (GFAP). Please click here to view a larger version of this figure.
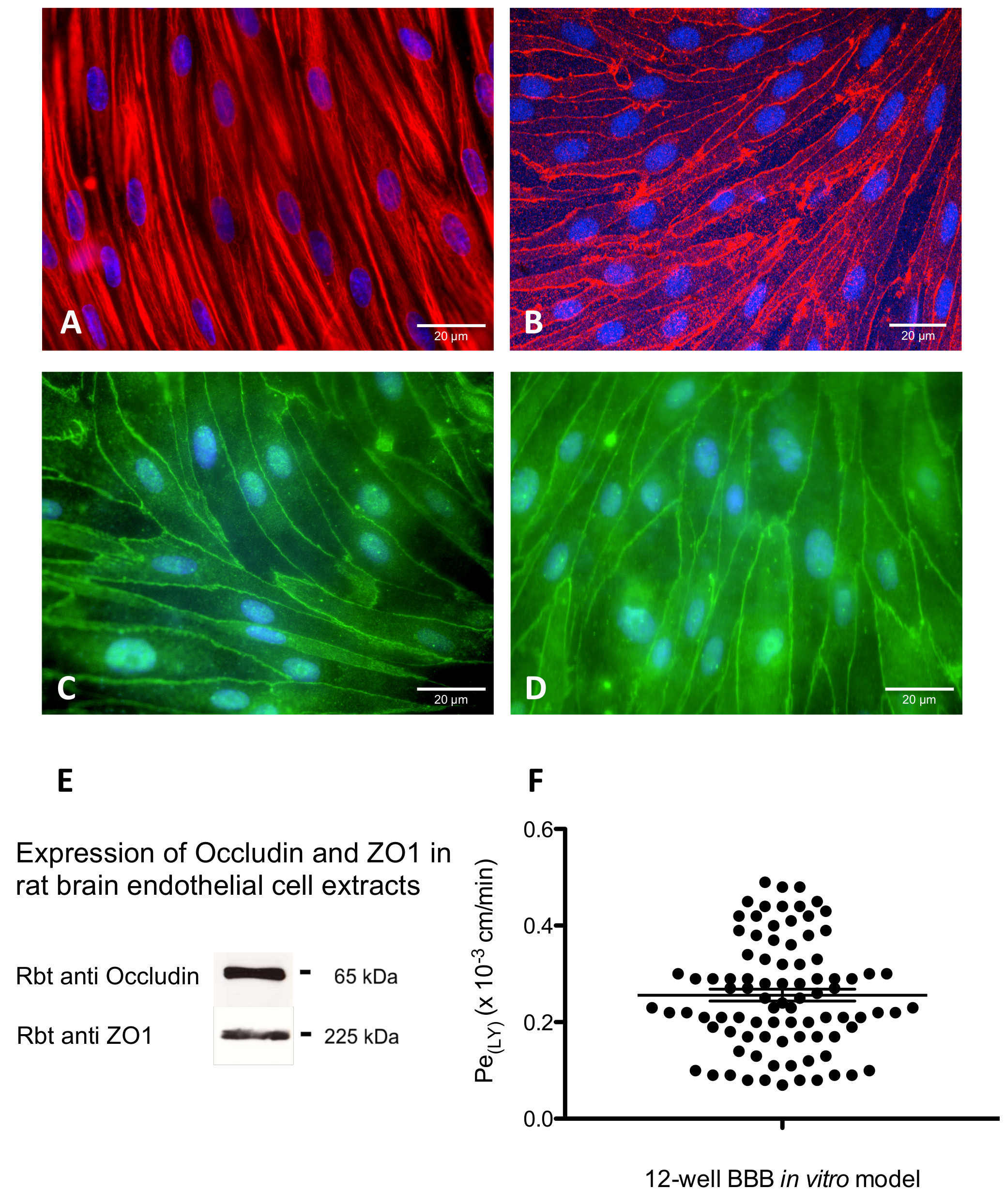
Figure 4. Immunocytochemical characterization of the rat in vitro BBB model and the paracellular permeability of LY across the endothelial monolayer. The cell monolayer was immunostained with (A) vimentin to reveal a confluent brain endothelial cell monolayer with non-overlapping morphology and typical spindle shaped cells with endothelial morphology and the expression of tight junction proteins (B) claudin-5, (C) ZO-1 and (D) occludin, also detected by western blot for ZO-1 and occludin proteins (E). (F) Monolayer tightness in the 12-well BBB model was assessed by measuring the transport of lucifer yellow (LY-CH, dilithium salt), a small hydrophilic molecule (MW 457 Da) known to be retained by the BBB. Briefly, LY was incubated in the upper compartment of the culture system in contact with cerebral endothelial cells for 60 min at 37 °C. After this time, the medium of the lower compartment was collected and fluorescence was quantified by fluorimetric analysis with a spectrofluorimeter with excitation at 430 nm, and emission at 535 nm. The results are expressed in permeability or Pe in 10-3 cm/min. The barrier is considered permeable or open when the Pe of LY is above 0.6×10-3 cm/min. During each 1 hr experiment, the average cleared volume was plotted versus time and the slope estimated by linear regression analysis. The slope of the clearance curve for control filters with collagen type IV-fibronectin coating was denoted PSf and the slope of the clearance curve for filters with brain endothelial cell monolayers was denoted PSt. The PS value for the endothelial monolayer (PSe) was calculated from: Please click here to view a larger version of this figure.
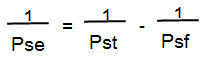
The PSe values were divided by the area of the porous membrane (1.1 cm2 for Millipore filters of 12-well plates) and the result was the permeability coefficient (Pe) with an average of 0.26 ± 0.11 x 10-3 cm/min. Results are presented with a vertical scatter plot representation for n = 90 assays (mean of n = 3 to n = 6 monolayers per assay).

Figure 5. In vitro BBB model response to treatment with the pro-inflammatory agent TNF-α. The culture media from the culture system was replaced just before TNF-α treatment at 5 ng/ml in the upper compartment for 6 hr, 12 hr and 24 hr. CCL2 release by RBEC was quantified in the upper compartment by ELISA assay in comparison with non-treated control. Note that MCP-1 levels increase slightly as a function of time, even in the non-treated wells. Integrity of the endothelial cell barrier measured by endothelial Pe for LY Pe(LY) revealed an increase in monolayer permeability by 24 hr treatment with TNF-α at 5 ng/ml or 50 ng/ml in the range of 0.62 ± 0.02 x 10-3 cm/min and 0.7 ± 0.01 x 10-3 cm/min respectively, compared to control at 0.33 ± 0.03 x 10-3 cm/min (Student’s t test, p < 0.05).

Figure 6. Functionality of the P-gp efflux transporter and RBEC polarization. Expression of the efflux transporter P-gp in brain endothelial cells co-cultured with astrocytes by immunocytochemistry (A) and western blot (B & C). Cell membrane proteins were extracted with a cellular protein fractionation kit. Plasma membranes were obtained by hypotonic lysis and differential centrifugations to eliminate organelles and nuclei. Separation of apical and basolateral proteins from plasma membranes were obtained by sucrose density gradient. P-gp was mainly localized in the apical fraction (F1) while Glutamate transporter-1 (GLT-1) was mainly localized in the basolateral fraction (F3). Purified microvessels prior to plating were used as control for the in vivo localization of these proteins. Activity of P-gp was determined by measuring the transport polarity of R123, a P-gp ligand. The flux of 1 µM R123 was measured for 1 hr at 37 °C in the luminal-to-abluminal (A to B) and in the opposite abluminal-to-luminal (B to A) directions. The same experiment was realized without RBEC to assess passive diffusion across the insert filter and data were normalized relative to this value for Pe(R123) calculation (see Figure 5 legend for Pe calculation method). R123 content in both compartments was measured with a spectrofluorimeter (excitation wavelength at 485 nm, emission wavelength at 535 nm). Data are expressed as 100% of the luminal-to-abluminal transport based on Pe(R123) (A to B). Verapamil (25 µM, 30 min preincubation) and cyclosporine A (1 µM, 30 min preincubation) were used as reference P-gp inhibitors. R123 accumulation in RBEC was measured after 2 hr with or without preincubation with the P-gp inhibitors (Student’s t test, p < 0.05). Please click here to view a larger version of this figure.
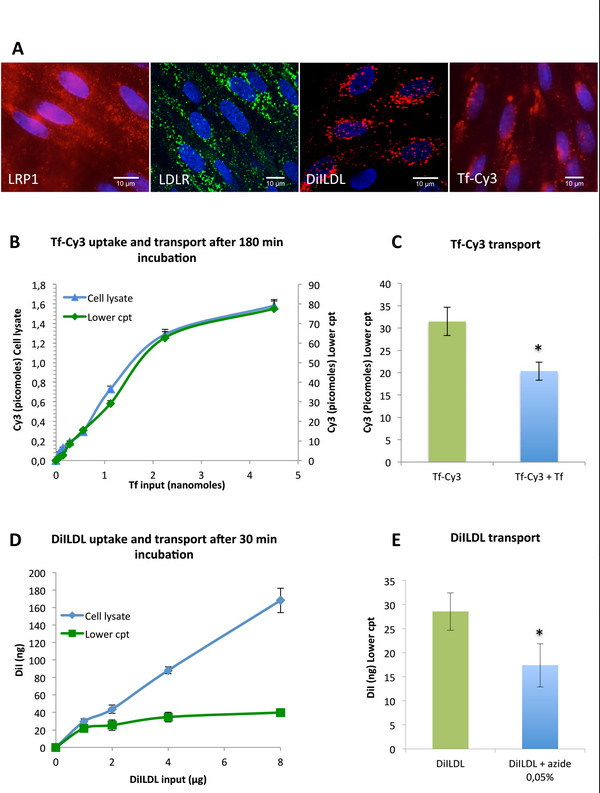
Figure 7. Characterization of receptors involved in Receptor Mediated Transcytosis (RMT). (A) The differentiated RBEC monolayer was immunostained with antibodies against the low density related lipoprotein receptor 1 (LRP1) and the low density lipoprotein receptor (LDLR). Confocal image of RBEC probed for uptake of DiILDL, a fluorescent ligand of the LDLR and for uptake of rat Transferrin-Cy3, a fluorescent ligand of the TfR show punctate staining over the cell surface, with some clustering in the perinuclear region. (B) Rat Tf-Cy3 was added to the upper compartment containing the differentiated RBEC monolayer at 75, 150, 300, 600, 1,200, 2,400 and 4,800 picomoles / insert for 180 min at 37 °C (200 µL in the upper compartment and 1,200 µL in the lower compartment). After this time, Cy3 fluorescence was quantified in the cell lysate (200 µL of PBS 0.1 % Triton X100) and in the lower compartment by fluorimetric analysis with a spectrofluorimeter with excitation at 550 nm and emission at 570 nm. Fluorescence units were transformed in picomoles using a linear working range. (C) For saturation experiments, rat Tf-Cy3 was added to the upper compartment containing the differentiated RBEC monolayer at 1,200 picomoles / insert for 180 min at 37 °C with or without 15 min pre-incubation with 4,800 picomoles / insert of non-fluorescent Tf. The transport in the lower compartment was quantified as in (B) (Student’s t test, p < 0.05). (D) DiILDL was added to the upper compartment containing the differentiated RBEC monolayer at 1, 2, 4 and 8 µg / insert filters for 30 min at 37 °C (200 µl in the upper compartment and 1,200 µl in the lower compartment). After this time, DiI fluorescence was quantified in the cell lysate (200 µl of PBS 0.1 % Triton X100) and in the lower compartment by fluorimetric analysis with a spectrofluorimeter with excitation at 554 nm and emission at 571 nm. Fluorescence units were transformed in µg using a linear working range. (E) For transport blocking experiments, sodium azide was added at 0.05% 15 min before the incubation of DiILDL at 2 µg / insert for 30 min at 37 °C. The transport in the lower compartment was quantified as in (D) (Student’s t test, p < 0.05). The absence of toxicity of sodium azide incubation at 0.05% was assessed by Pe(LY) measurement (data not shown). Please click here to view a larger version of this figure.

Figure 8. In vitro BBB models from rat spinal cord endothelial cells or from mouse brain. The table highlights important parameters for microvessel extraction from rat brain, rat spinal cord and mouse brain. Photomicrographs show immunostaining for ZO1 and occludin in endothelial cell monolayers prepared from rat brain, rat spinal cord and mouse brain.
