Survival rates.
In a representative study conducted by us we had a survival rate of 16 out of 20 female Lister hooded rats (200 to 250 g). The most common complication is breathing difficulties because of the trachea displacement or damage to the respiratory centers in the brainstem just dorsal to the pyramids (nucleus solitarii, ambiguous and parabrachilis). Similar results have been reported in the literature 16.
Behavioural outcomes.
A range of behavioural tests have been used to assess sensorimotor outcome after pyramidotomy in rodents: the horizontal ladder test, the single pellet reaching test, the cylinder rearing test and Montoya staircase test are the most commonly used behavioural assessments. Other tests described in the literature are the pellet reaching test, rope climbing test, gait analysis and the sticky tape test 3,17-19. Most of these show a functional motor deficit 1 week after injury. Currently, most studies described in the literature only perform behavioural testing up to 42 days 4,11,13,20,21, or for a shorter time 3,18,22. We have performed behavioural testing for a prolonged period of time and have seen essentially full recovery of motor function 8 weeks post pyramidotomy for some of the behavioural tests, except the Montoya staircase test and the horizontal ladder test. This is probably for several reasons. Firstly, the injured CST rostral to the lesion may form synapses on neurons in the brainstem that act as indirect relays to motor neurons in the spinal cord. Secondly, the uninjured (contralateral) CST may sprout and forms synapses on neurons in the brainstem or spinal cord that act as relays to motor neurons. Thirdly, other spared motor tracts such as the reticulospinal, vestibulospinal or rubrospinal tracts may take over some function. Thus, a lasting treatment effect is difficult to detect with many behavioural tests in the pyramidotomy model. Some behavioural testing in this model however could indicate accelerated recovery early after the injury, which is related to a treatment.
Cylinder Rearing test 3,4,23
Based on the cylinder rearing test the CST corresponding to the dominant forepaw was injured. Rats are placed in a plexiglass cylinder and rearing behaviour is observed for a period of 3 min. Preinjury there was a slight preference for the use of one forepaw, the dominant forepaw, during vertical exploration (Figure 3A). The injury affects the dominant side and the use of the contralesional forepaw decreased to 28% at 1 week postinjury (Repeated Measures ANOVA p <0.05; post-hoc analysis revealed animals have significant deficits at Week 1, 2 and 3, p <0.05, Fisher’s LSD). 4 weeks postinjury the animals recovered and used the affected forelimb for 45% of rears, and after 6 weeks this was 57% (n = 16, mean ± standard error is shown). Similarly, Starkey et al. (4) have assessed mice using the cylinder test for up to 42 days after pyramidotomy and reported a significant decrease in the use of the affected forepaw throughout the whole testing period.
Montoya Staircase test 24
Rats require pretraining for this test to acclimatize prior to injury. To increase motivation during the testing session animals are food restricted to 15 g food per rat the night before. The food deprived rats are placed in the Montoya staircase for 15 min. On each side, there is a staircase of 7 steps with wells, each containing 3 pellets. The total number of retrieved pellets on each side are recorded. The number of retrieved pellets with the contralesional/affected forepaw decreased significantly from 61% preinjury to 3% 3 days postinjury (Figure 3B). The ability to retrieve pellets was still significantly impaired at 4 weeks postinjury with 20% of pellets retrieved. Rats show a significant deficit up to week 8 with only 29% of the pellets eaten at that time point (Repeated Measures ANOVA p <0.05, post hoc analysis revealed animals have significant deficits on the contralesional side at all weeks, p <0.05, Fisher’s LSD). The ipsilesional forepaw is affected up to 1 week postinjury; however this recovers by the second week (Repeated Measures ANOVA p <0.05, post hoc analysis revealed animals have significant deficits at Day 3 and Week 1 on the ipsilesional forepaw, p <0.05, Fisher’s LSD). The initial deficits of the ipsilesional forepaw might be explained by the lesioning of the ipsilateral portion of corticospinal tract running through the cut pyramid, with subsequent sprouting and recovery.
Horizontal Ladder test 25
Rats require pretraining for this test. Rats walk three times over a 1 m long ladder with irregular rung spacing. Videos are analysed later in slow motion and the total number of foot slips and misses for each paw is quantified. The graph in Figure 3C represents the total number of misses and slips of the affected and less affected side (forepaw + hindpaw) as a percentage to the total number of steps taken. Rats have a significantly increased number of errors 1 week postinjury with 10% or 11% on the affected side and less affected side respectively. This deficit is persistent on the contralesional/affected side up to week 8 with 10% errors made, however, resolves on the ipsilesional/less affected side by 2 weeks (Repeated Measures ANOVA, post hoc analysis revealed animals have significant deficits at all weeks on the contralesional side and significant deficits at Week 1 and 3 on the ipsilesional side, p <0.05, Fisher’s LSD).
Electrophysiological testing
In a terminal electrophysiological set-up the animal was anaesthetized intraperitoneally with urethane. Throughout the procedure, it was kept at 37 °C ± 1 °C. Its ventral neck region, its forelimbs and chest region were shaved and disinfected with iodine scrub. The pyramids were exposed as previously described and the radial nerve was exposed in the upper arm by a ventral approach incising the skin and the pectoralis major. The distal end of the radial nerve was cut and the nerve was laid over two silver-wire hook-electrodes in a mineral oil bath. Either pyramid was stimulated with concentric bipolar electrodes at different depths with 5 pulses at 300 Hz and increasing stimulation amplitude. Recordings were always made from the radial nerve of the disabled arm (Figure 4).
Uninjured animals had a strong response at latencies between 12 to 20 msec in the radial nerve when the contralateral pyramid was stimulated (Figure 4B). Ipsilateral stimulation only results in occasional activation (Figure 4C). Animals, which have previously received pyramidotomies, on the contralateral side to the recording arm show no response in the radial nerve when the lesioned tract is stimulated above the lesion (Figure 4B’). However, stimulation of the CST fibers in the pyramids ipsilateral to the recording electrodes resulted in a stronger activation in the radial nerve compared to uninjured animals at 12 weeks postinjury (Figure 4C’).
Anatomical outcomes.
Animals were euthanized 10 weeks postsurgery according to Schedule 1 of the Animals (Scientific Procedures) Act 1986. Two weeks previously, the uninjured CST was traced with biotinylated dextran amine (BDA) injections into the motor cortex 26 ipsilateral to the disabled arm (Figure 1). This allows quantification of uninjured fibers, which cross the midline and supply the contralesional spinal cord (Figure 5C, D). Uninjured animals only have a few fibers crossing the midline, whereas this increases in response to injury. Transverse cervical spinal cord sections were stained with protein kinase C (PKC) gamma to assess the proportion of denervation in the lesioned CST (Figure 5E)16. Alternatively, the completeness of the lesion can be assessed by injecting BDA tracer into the motor cortex on the ipsilesional side of the pyramidotomy injury. The absence of labeling in the contralateral dorsomedial CST and the ipsilateral ventromedial CST provides evidence of complete unilateral transection.

Figure 1: Schematic showing the corticospinal tracts in the rat and unilateral axotomy of the CST in the pyramids. Top image: Corticospinal neurons originate in the cortical layer 5 pyramidal cells. Biotinylated dextran amine (depicted in green) is injected into the contralesional cortex two weeks prior to the end of the experiment. Middle image: The tracts form the pyramids on the ventral surface of the medulla and at the spinomedullary junction the majority of axons decussate. Unilateral pyramidotomy is performed in the caudal medulla (shown in red). Lower image: The majority of the corticospinal tracts run down in the dorsomedial and dorsolateral parts of the spinal cord. A minority stays ipsilaterally and runs ventromedially.

Figure 2: Ventral view of the rat brain. The pyramids are distinctive because they are raised, parallel convexities (yellow box). The borders of the pyramids are defined medially by the basilar artery (bas) over the midline and laterally at the base of the convexity halfway between the basilar artery and the paraolivary artery (pol) as seen in (C). Craniotomy is performed over the basilar artery and lateral extending to the paraolivary artery (red box in C). One pyramid is cut perpendicularly to the basilar artery (red line in C, cut is shown on specimen in A and B). Lesions placed too caudally near the vertebral arteries (vert) will be incomplete because the CST starts to decussate and because the vertebral artery is set off the midline. (C) Image adapted from “The Rat Nervous System” by George Paxinos27. NOTE: This Figure shows a lesion on the left hand side of the images: this is the animal’s right pyramidal tract because it is lying supine. Please note however that the video shows a lesion to the animal’s left pyramidal tract.

Figure 3: Representative results for the cylinder rearing test, Montoya staircase pellet reaching test and Horizontal ladder test. There is a motor deficit detected by all tests after injury. However, no deficit can be detected with the cylinder rearing test at 4 weeks postinjury, whereas sustained deficits can be detected with the Montoya staircase test and the horizontal ladder test up to 8 weeks postinjury (n = 16 per group, means and standard error are shown). Asterisk indicates p <0.05 relative to preinjury baseline (Fisher’s LSD test).Please click here to view a larger version of this figure.
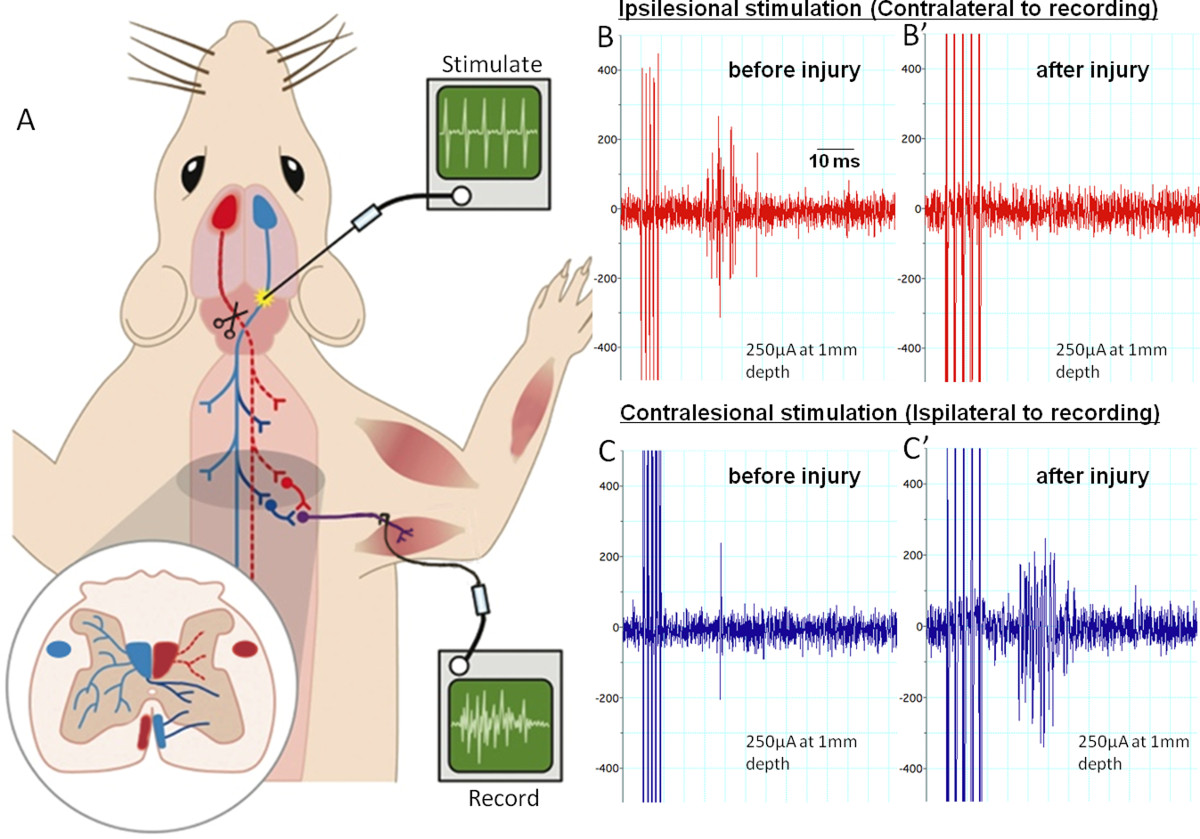
Figure 4: Electrophysiology assessing the corticospinal tract before and after pyramidotomy. (A) shows the set-up for the terminal experiment. We stimulate the pyramids with a train of 5 pulses at various depths rostral to the site of lesioning and decussation. The red tract is the lesioned tract. We record from the radial nerve on the contralesional side. (B) and (C) show example recordings with ipsilesional stimulation or contralesional stimulation before or 12 weeks after pyramidotomy. Ipsilesional stimulations evoke multiple compound action potentials (spikes in the trace) in the radial nerve before injury (B). These are abolished by the injury and activity does not return within 12 weeks (B’). Contralesional stimulation evokes rare single compound action potentials preinjury (C). 12 weeks after injury activity considerably is increased (C’), possibly due to intact axons sprouting across the midline and plastic changes within the spinal cord. Please click here to view a larger version of this figure.

Figure 5: Anatomical evaluation after pyramidotomy. (A, B, C, D) Biotinylated dextran amine was used to trace the corticospinal tract by injecting it into the motor cortex corresponding to the unlesioned CST (see green in Figure 1). The dorsal medial, the ventral medial components of the CST are clearly visible (A, dorsal medial part magnified in B). (C) and (D) are magnifications from (A) of fibers sprouting across the midline into the grey matter of the denervated side in response to a treatment. To assess the completeness of the lesion transverse sections of the cervical level 4 (12 weeks postinjury tissue) were cut and stained with antibodies against PKC gamma (E). The CST on the left hand side of the image is intact and can be seen bulging across the midline into the denervated side, presumably because of the structural disintegration of the injured dorsal CST.
