Susceptibility to infection with H. polygyrus is controlled to a large degree by the genetic background of the mouse strain (Table 1); C57BL/6 and CBA mice are highly susceptible 21,22. For maintenance of the parasite life cycle, the F1 hybrid between these two strains has been chosen for its ability to withstand much higher worm burdens without morbidity (excessive intestinal epithelial damage) compared to either parental strain. Oral gavage of 400 L3 larvae is used to maintain the life cycle in F1 mice (resulting in adult worm burdens shown in Figure 2), whilst a dose of 200 L3 larvae is generally used for experiments in homozygous inbred strains (e.g., C57BL/6 or BALBc). However, this dosage may need to be reduced depending on environmental co-factors that may differ between animal facilities, such as variations in gut flora.
Batches of HES have proven reproducible efficacy in functional assays and in protein composition; moreover, when supernatants from each successive week of culture were analysed, up to a total of 4 weeks, the protein profiles were found to be very similar (Figure 3). When HES concentration is measured by Bradford assay (see 5.5), the total protein is usually approximately 1 mg/ml (Figure 4).
An alternative method of concentrating HES is with a centrifugal concentrator (eg Vivaspin 3-kDa cut-off membrane), in which samples are spun at up to 4,000 g in a swing-out rotor centrifuge, removing buffer salts and low molecular weight components. Centrifugal concentration is best suited to small processing volumes (1-10 ml) and are limited to a maximum concentration factor of approximately 30x.
When collecting HES, avoidance of contamination is critical. To avoid contamination with host molecules, we discard HES-containing culture media from the first 24 hr after adult worm harvest from the mouse intestine. We also quantify the level of LPS contamination in each batch of HES with a Chromogenic LAL assay (see 5.7). 1 U of LPS equates to ~100 pg LPS and levels below this are considered negligible 23. In our hands, most batches of HES are significantly below this limit, the mean concentration of LPS in HES being 0.23 U/µg (Figure 5). The effects of LPS in in vivo models of pathology (for instance the suppression of asthmatic responses) requires at least 10 ng of LPS 23. Hence in vivo administration of 5 µg HES from a batch with 100 pg LPS/µg HES will include 500 pg LPS, well below the effective concentration where LPS becomes a problem.

Figure 1: Baermann Apparatus Baermann Apparatus setup for collection of adult H. polygyrus (as described in section 2).

Figure 2: Mean Worm Burden 14 Days After Infection with 400 L3 Larvae Mean worm burdens 14 days after infection with 400 L3 larvae. Data points shown are from 19 separate rounds of infection of C57BL/6xCBA mice. Mean ± SEM shown.

Figure 3: Protein Profiles of HES From Successive Weeks in Culture SDS-PAGE profile of HES proteins collected in successive weeks of culture.

Figure 4: Final Quantity of HES from 500 ml ES-Containing Media Yield of HES protein from 11 different batches derived from approximately 500 ml of culture supernatant. Mean ± SEM shown.

Figure 5: LPS Contamination of HES Levels of LPS contamination in 41 batches of HES measured by the Limulus ameobocyte assay. Median LPS concentration = 86 U per mg of HES.
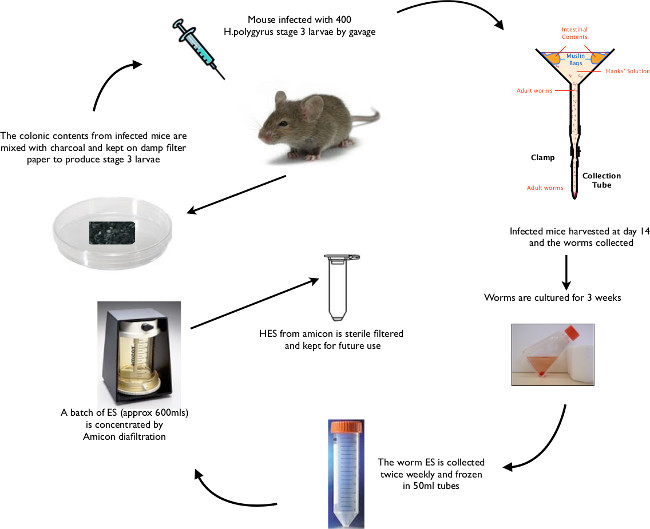
Figure 6: Animated Schematic of H. polygyrus Life Cycle Summary of key life cycle stages from oral gavage of L3 larvae, through recovery of larvae and adult worms to isolation of HES.

Figure 7: Animated Schematic of HES and its Functions Key immunomodulatory effects of soluble mediators and exosomes contained within HES.
| Genotype (and background strain) | Primary Infection Phenotype | Reference |
| Inbred strains | ||
| A/J, CBA, C3H | Highly susceptible | 22,29 |
| C57BL/6 and C57BL/10 | Susceptible | 22,30 |
| BALB/c, DBA/2, 129/J | Intermediate | 22,31 |
| NIH, SJL, SWR | Low susceptibility | 22,32 |
| Transgenic for cytokines or cytokine receptors | ||
| IL-1β-/- | More susceptible | 33 |
| IL-1R-/- | Less susceptible (a); no change in susceptibility but increased granulomas (b) | (a) 33 |
| (b) 34 | ||
| IL-2Rβ Transgenic (C57BL/6) | Resistant | 35 |
| IL-4-/- | Higher fecundity | 36 |
| IL-4R-/- (C57BL/6 or BALB/c) | Highly susceptible | 22 |
| IL-6–/– (BALB/c or C57BL/6) | Highly Resistant | 37 |
| IL-9 Transgenic (FVB) | Resistant | 38 |
| IL-21R –/– (C57BL/6) | Deficient Th2, decreased granuloma gormation | 39,40 |
| IL-25-/- (BALB/c) | More susceptible | 33 |
| IL-33R (T1/ST2) -/- (BALB/c) | No change in susceptibility | 33 |
| TGFβRIIdn (C57BL/6) | High Th1, Increased susceptibility | 41,42 |
| Transgenic for T cell markers | ||
| CD28–/– (BALB/c) | Marginally higher fecundity | 43 |
| CD86 (B7-2)–/– (BALB/c) | Higher fecundity | 44 |
| OX40L–/– (BALB/c) | Higher fecundity | 45 |
| Transgenic for innate Immune loci | ||
| Type 1 interferon receptor (IFNAR)-/- (C57BL/6) | Higher fecundity, more granulomas | 34 |
| MyD88–/– (C57BL/6) | More resistant, more granulomas | 34 |
| C-KitW/Wv (WBB6) | More susceptible | 46,47 |
Table 1: Primary infections with H. polygyrus in genetically different and gene-targetted mouse strains.
