

Summary
在这项研究中,纳米微流体流动室被用于可视化和功能上表征叶缘焦枯病菌,导致皮尔斯病在葡萄细菌的抽搐蠕动。
Abstract
叶缘焦枯病菌是革兰氏阴性非鞭 毛的细菌引起的若干植物的经济上重要的疾病。抽搐蠕动提供X. fastidiosa长距离内厂运行和殖民化,手段在十朝贡献致病fastidiosa,X的运动性抽动fastidiosa是由IV型菌毛操作。 叶缘焦枯病菌的IV型菌毛由pilG,在弼CHP操纵子编码的蛋白质趋化性调节器所涉及的信号转导通路调节。为了阐明pilG的角色X的抽动运动fastidiosa,一个pilG缺陷型突变体Xf为 ΔpilG及其互补株XfΔpilG-含有C本地pilG被开发。与时间推移图像记录系统集成的微流控室,观察抽搐蠕动在XfΔpILG,XfΔpilG-C及其野生型菌株。 利用这种记录系统中,其允许聚合的长期的空间和时间的观察,个体细胞,并通过抽动蠕动细菌种群的迁移。X. fastidiosa野生型和互补XfΔpilG-C株显示,微流体流室直接观察到典型的抽动运动特征,而突变XfΔpliG展出抽搐缺陷表型。这项研究表明,pilG有助于X的运动性抽动fastidiosa。微流体流动室被用作用于观察抽动运动的装置。
Introduction
叶缘焦枯病菌是革兰氏阴性非鞭 毛,病原菌引起许多经济上重要的作物疾病,包括皮尔斯在葡萄病( 葡萄属)1,2,3条。这种细菌是有限的导水木质部船只。葡萄感染引起木质部导管和水分胁迫的结果和营养不足3堵塞。成功定植取决于细菌的从感染的初始位点移动到植物3的其余部分的能力。抽搐蠕动是鞭毛无关细菌移动的装置通过扩展,附件,和极性IV型菌毛4已特征在于X的缩回fastidiosa 5,6,7。
抽搐运动已由激光镊子和原子力显微镜(AFM)8,9,10观察。使用这些技术,叔由N型菌毛IV产生的魔力motilities 淋球菌和P.由FL uorescently标签菌毛和显微镜捕捉他们的动作绿脓杆菌进行了表征。虽然这两种方法有详细的个体细菌的粘合力,该程序是复杂和耗时9,10。微FL uidic室,观察单个细胞的长途迁徙,以及细菌细胞5,6小聚集。这些室被设计成与隔时图像记录系统11,12,13,14一体的板一个微制造纳米通道。微FL uidic室装置用于研究细菌的运动行为和细胞 - 细胞相互作用提供几个优点:(i)其提供了具有多个信道功能的集成平台; (ii)其可以检查运动和单细胞的细菌的纳米级特征的聚合; (三)它允许直接米细菌细胞和时间推移分析icroscopic图像记录,(ⅳ)它提供了一种在微环境的细菌的个体和/或群体的长期的空间和时间的观测; (五)培养基中的信道的流量可以精确控制和培养基(ⅵ)仅有一个非常小的体积(1ml)中,需要对每个实验。
最近,微FL uidic溢流系统已被用于研究在各种微环境14,15,16细菌细胞的行为。的密合性和E的表面附着大肠杆菌 15,X。 fastidiosa 16和敏捷食酸citrulli 14到玻璃表面用微FL uidic室评估。由敏捷食citrulli的IV型菌毛介导的聚集和生物膜的形成进行了分析14。另外,A的运动citrulli下溢流Ç观察onditions表明,IV型菌毛可能在殖民化中发挥重要作用,传播的A. citrulli在木质部导管在边溢流条件。 铜绿假单胞菌和X的抽搐motilities fastidiosa细胞在微加工流动室5,6,17针对流体当前成功观察。 IV型菌毛短少X的pilB和pilQ突变fastidiosa被发现深刻地改变微FL uidic装置5,6,18的溢流条件下抽搐运动的速度。在微FL uidic器件细菌粘附和运动进行的研究表明,微FL uidic室是特别适合于在体外分析菌毛介导的细菌的抽搐运动和迁移。这些结果可以解释这有利于细胞间的连接,聚集和殖民内的抽动介导的迁移机制主机,最终导致全身感染。
X的弼热电联产操纵子fastidiosa包含pilG,霹雳,pilJ,丸剂,chpB和CHPC其中编码信号转导通路20。跨膜化学感受器结合化学刺激在周质结构域和在其细胞质部分以最终控制细菌抽搐蠕动激活信号级联。在X的弼CHP操纵子fastidiosa,一个磷酸化蛋白班车是PilG同源到崔泰源。在E.大肠杆菌和P.假单胞菌,崔泰源是在与该鞭毛马达蛋白19,21交互趋化系统的响应调节。虽然弼CHP操纵子在X的贡献对毒力fastidiosa最近检查20,pilG在趋操纵子的作用,响应于环境信号及X的调节/马达IV型菌毛fastidiosa是UNC李尔。为了阐明X的抽动运动的活动趋调节pilG的洞察力fastidiosa,微FL uidic室被用来评估X的抽搐蠕动fastidiosa。 十fastidiosa的pilG的特征在于,通过比较一个缺失突变体 Xf为 ΔpliG,互补应变XfΔpliG-C及其野生型在体外 的表型 。结果突出pilG在X的抽动运动中的作用fastidiosa。
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1.细菌菌落外周边缘
- 十,成长fastidiosa(Xf为 )蒂梅丘拉野生型22,pilG缺失突变Xf为 ΔpliG(使用先前描述删除策略23),以及其互补XfΔpliG-C(使用先前描述基于染色体遗传互补策略24)上的 PD2培养基琼脂板25在28℃下5-7天。
- 高压釜玻璃纸(1×1 cm 2)的水在121℃(249°F)的15分钟。拿起一块玻璃纸,通过在一个空的培养皿接触玻璃纸的一个角落里沥干水,仔细地打好玻璃纸在琼脂表面和空气干燥的15%。
- 拿起个别X. fastidiosa菌落用无菌牙签圆润和现货细胞无菌到在琼脂平板覆盖在琼脂表面的15%的玻璃纸杀菌片。孵育所述板■在28°C 2-3天。
- 检查使用具有2X物镜解剖显微镜和10X目镜殖民地的边缘形态。照片周围的殖民地外围边缘。
2.显微镜和微流控流动室
- 先前描述5,18使用类似于光刻过程制造微流体装置。使用标准光刻方法26设计与电脑辅助设计软件的主硅片上四个并行通道。
- 与聚二甲基硅氧烷(PDMS)创建一个从硅片主微流控室。倒未聚合的PDMS在硅片主,并在60℃下固化它为1小时。剥离从晶片主与PDMS复制品并修剪PDMS复制品用刀片成22毫米×40mm的作为玻璃盖玻片的大小相同。
- 露出PDMS复制品,在玻璃盖玻片(22×40mm的2),以及一个microscOPE幻灯片(为50 x 75 平方毫米)空气等离子体在30W 2分钟27。夹层玻璃盖玻片,玻璃显微镜打造微FL uidic室之间的PDMS身体。
- 在图案化的通道的每一端钻透PDMS中的孔(5.5毫米直径)。切硅橡胶管插入长12-20厘米。插入硅橡皮管的一端(外径5.1毫米,内径2.1毫米,0.8毫米壁)插入PDMS复制品的信道的各开口端部,并在60℃下用未聚合的PDMS密封它为1小时。
- 管子的另一端连接到塑料鲁尔连接器的倒刺末端。包组装微流体腔室与铝箔和高压灭菌他们20分钟。
- 十,收集细菌细胞fastidiosa野生型,突变体Xf为 ΔpliG,互补XfΔpliG-C通过刮,用一次性接种从PD2中板循环。调整细胞密度到0.05在PD2肉汤600纳米的光密度如前面23所描述。收集细菌细胞溶液到1毫升气密注射器。
- 安装在倒置显微镜阶段微流体装置。入口管连接到包含PD2肉汤中的5毫升注射器气密性。适应中的5毫升注射器气密用注射器泵。
- 出口管连接到废物储存器。保持0.2微升min的介质流速-1 30分钟以稳定该系统。
- 侧入口管连接到含有细菌细胞溶液1毫升气密注射器。冲洗通过橡胶管的细菌细胞溶液,直到达到信道。以冲洗从室图像捕获之前未结合的细胞保持0.2微升min的介质流速-1另外的30分钟。
- 安装在显微镜来控制光的场调整部分在显微镜下快门。活门连接到快门共ntrol系统和闸门控制系统连接到计算机。
- 安装一台数码相机到显微镜的视频端口,并将其连接到电脑。运行时间推移录音软件,从菜单中选择了“快门”功能,并自动识别软件安装快门为默认建立与软件的快门连接。
- 从时间推移刻录软件的菜单中选择“数码相机”功能可以自动识别数码相机作为软件默认的捕获设备,并建立与数码相机相关的软件通信。
- 定位在使用20X相衬光学通道之一的细菌细胞,然后切换到之前的图像捕获的40X物镜。
- 运行时间推移录音软件,使用从菜单中默认参数,以获得从显微镜图像选择“图像采集”功能。接下来,打开“采集延时”功能,时间间隔设置为30秒5,18,28为根据,以观察X的运动性抽动实验需要6-24小时的时间fastidiosa 5,18,28。点击“确定”,启动延时录制。点击“堆栈功能”,从菜单中选择“另存为”,以完成录音后,堆在目标文件夹中的图像在计算机上。
- 对于多个通道,从第一通道捕获时间推移的图像,每隔30秒6小时。移动显微镜的物镜到下一个通道来定位目标细胞。如上所述,如果实验设置为利用四个通道捕捉图像中的每一个顺序地四个通道重复时间推移功能。继续只要连续三天的时间推移图像采集。所有实验均在室温(23±2℃)下进行。
- 编译时间推移图像成使用时间推移录制的图像处理软件的视频文件。运行时间推移录音软件,单击“堆栈功能”,从菜单,并选择“打开堆叠功能”,从计算机打开堆放的文件。
- 从堆栈模块启动“制作电影”功能,选择所有图像并选择“AVI”格式输出。点击“另存为”的视频文件存储在计算机上的目标文件夹。
- 选择在计算机上从目标文件夹编译电影并播放它们。然后,通过细菌细胞的抽搐运动活动中所生成的视频文件生成的可视化观察单个细胞的运动。
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
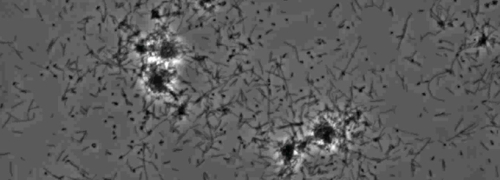
表示IV型菌毛介导的抽搐运动的周菌落边缘的情况下,在X的菌落观察fastidiosa野生型和互补Xf为 ΔpliG-C菌株( 图1)。突变XfΔpliG,然而,并没有表现出在菌落周围的周边的边缘( 图1)。在纳米微流体流动腔室的细菌细胞的延时成像显示,在野生型X的观察抽搐蠕动fastidiosa和互补XfΔpliG-C(补充V1,V3),而XfΔpilG突变的细胞没有表现出在整个实验(补充V2)抽搐蠕动。突变XfΔpilG的细胞在肉汤PD2(补充V2)形成相对较小疏松的聚集。 X的相反,细胞fastidiosa野生型和互补XfΔpilG-C D在PD2肉汤eveloped较大的聚集体( 图2,(补充V1,V3)。

图1:X的细菌菌落的外围边缘殖民地缘特征从野生型,突变体Xf为 ΔpilG,互补XfΔpilG-çfastidiosa生长在覆盖着玻璃纸的杀菌片PD2琼脂。与突变体XfΔpilG的以外,所有的菌落表现出周边边缘,表示IV型菌毛介导的抽搐蠕动。照片是经过培养基上生长5天服用。放大倍率吧,0.5毫米。 请点击此处查看该图的放大版本。
_upload / 53816 / 53816fig2.jpg“/>
图2:X的运动性抽动 fastidiosa 细胞的纳米微流体流室。所有测试菌株细胞的运动抽动在观察6天记录。评估从三个独立的视频段进行。放大倍率酒吧,20微米。
注:X的运动性抽动fastidiosa细胞是通过扩展,附件,和极性IV型菌毛缩回其特征在于通过在玻璃表面的单细胞的运动。在迁移中观察到优先对在微加工的流动室的流体当前的单个单元格。 请点击此处查看该图的放大版本。
6 / 53816supfig1.jpg“/>
参考图1:四通道微流室 。在每个月底在媒体和媒体输出接头每个通道。 请点击此处查看该图的放大版本。
补充电影1:运动性抽动。 (点击下载)。野生型X. fastidiosa抽搐在微流体流室。
补充电影2:受损颤搐摹蠕动。 (点击下载)。Xf为突变体蠕动。 XfΔpilG在amicrofluidic流室观察。
补充电影3:恢复抽搐蠕动。 (点击下载)。抽动Xf为补充的应变蠕动。 XfΔpilG-C中的微流体流动室观察。
Subscription Required. Please recommend JoVE to your librarian.
Discussion
在这项研究中,我们的特点X的运动行为fastidiosa PilG突变Xf为 ΔpilG和新设计的多个并行的纳米通道微FL uidic室互补XfΔpilG-Ç株。新设计的微FL uidic腔室可以具有100微米的纳米通道多达四个平行腔室宽度相比仅与单个50微米宽的通道18早期的设计。改进的更宽纳米通道便于引入细菌细胞与媒体的流动。此外,该微流体室1)简单的构建和组装; 2)相对便宜;和3)容易适用于不同的试验要求。其结果是,该室设计允许的细菌细胞的运动下的品种长期的观察实验的微环境。
稳定溢流的电流THROU的GH微FL uidic信道是根据各种实验环境中创建用于细菌细胞的运动的观察完整的流入微FL uidic微环境中关键的一步。的微FL uidic腔室的冲洗和连接管之前与引入细菌细胞的培养基也以稳定腔室中的流动的一个重要步骤。但是,该流的高速将刷新细菌细胞从室的无滞留细胞在腔室的过程中。需要流经微FL uidic通道媒体的适当的速度,以使用注射泵来调节。在本研究中的实验中,流过被设置在0.2到1微升分钟-1至少30分钟前引入细菌细胞进入通道,以通过泵稳定。一旦细胞被引入微FL uidic通道,所述介质溢流保持在0.2微升米在-1 30到60分钟,以稳定该系统,并删除nonattached细胞。以稳定的流动和保持背景明确,以观察细菌细胞的运动是很重要的。图像被捕获的,每30秒到CON组fi RM导入细胞以0.2微升分钟-1保持恒定流速的抽搐运动活性。如果实验需要在腔室中的细菌细胞的收集,细胞可通过调节注射器泵的转速为0.2至110微升分钟-1逐渐增大介质的溢流速率来冲出。
在微FL uidic室中的细菌细胞的活性是通过使用40X相差的光学元件和记录的数字照相机,其通过成像软件控制时间推移图象的倒置显微镜评估。流动速度和时间间隔为图像捕捉可以相应与experim调整ental要求。然而,在大多数情况下,对于一个介质流率设定为0.2微升分钟-1随时间推移的图像记录的菌毛介导的细菌细胞每30秒流动速度。在其它情况下,如果介质溢流速率增至0.1微升分钟-1的实验过程中,细菌细胞的行为将被捕捉图像,每10至15秒相应记录。时间推移的图像取每30秒为6-24小时的持续时间,并保存为每个测试菌株的来源的图像文件。影片然后从6-8小时从每个菌株,其中在突变体和野生型之间的抽动motilities所示相反的表型拍摄的图像遵从/补充株(参考V1,V2,V3)。
在宏观平行板溢流室的纳米微FL uidic室设备的意义包含了运动的直接检查,唱的聚合细菌LE细胞。除了 低成本5,低试剂和样品量的要求,微FL uidic室的优点是便于施工进行细菌培养,并在流体溢流速度精确控制溢流微环境的。多个平行通道允许差动细菌菌株的观察在单一实验设置其中用于分析提供兼容的数据。与极性菌毛的细菌的鞭毛无关抽搐运动是特别适用于本纳米微FL uidic室分析。然而,这种微FL uidic室是不太适合鞭毛依赖细菌运动,其中细菌的运动通常太快并呈现随机的方向。这种限制有时可以通过在每1 至2秒调节介质流率,以0.05微升分钟-1和改变隔时图像捕获率受到损害为1小时至ANALY泽在纳米尺度环境鞭毛介导的细菌的运动。
本文所使用的微流控室设备提供了PilG负责体外运动行为的功能评价直接的视觉证据。此外,该研究还表明,细胞通过抽动运动细胞聚集是生物膜的形成至关重要, 十信令和致病性fastidiosa。细菌通过微流体装置的抽动运动的可视化提供了新的方法来研究涉及这是不容易被其他分析方法测定细菌行为的基因功能。这种方法可以应用到其它细菌系统中。微流体腔室的设备用于表征与细胞 - 细胞附件,细胞聚集,运动模式和生物膜形成相关的细菌细胞的生理行为提供一种流系统。
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
作者宣称,他们没有竞争的经济利益。
Acknowledgments
这项研究是由美国农业部农业研究服务支持。本出版物中的商品名称或商业产品只是提到提供具体信息之目的,并不由农业部美国农业部暗示推荐或认可。美国农业部是一个平等机会提供者和雇主。
Materials
| Name | Company | Catalog Number | Comments |
| Biology materials | |||
| X. fastidiosa (Xf) Temecula wild type | Costa, H. S., et al., 2004 22 | ||
| pilG deletion mutant XfΔpliG | Shi, X. Y., et al., 2007 26 | ||
| pilG complementary strain XfΔpliG-C | Davis, M. J., et al. 1998 23 | ||
| Physical materials and equipment | |||
| Disposable inoculating loops | VWR international, Radnor, PA | #22-363-607 | quantitative procedures such as bacterial collection |
| Polydimethylsiloxane (PDMS) | Dow Corning Corporation | #0002709226 | Sylgard 184 silicone Elastomeric Kits |
| AmScope MD2000 digital camera | AmScope, Irvine, CA | SE305R-AZ-E | Image, video recording and measurement |
| Tubes line | Edgewood, NY | #T4300 | Connected to the syringe and microfluidic chamber |
| Plastic luer connectors | Edgewood, NY | Connected to the syringe and microfluidic chamber | |
| Syringe pumps | Pico Plus, Harvard Apparatus, MA | #702209 | The flow rate can be adjusted while the pump is running. |
| Syringes | Gastight, Hemilton Company, Reno, NV | #1005 | Provide the flowing broth |
| Inverted Olympus IMT-2 microscope | Olympus | IMT-2 FLuoro PHase | Image observation and recording |
| SPOT-RT digital camera | Diagnostic Instruments, Inc., MI | RT230 | Image, video recording and measurement |
| Microscope Shutter | The UNIBLITZ, US | #LS2T2 | Control camera’s exposure time |
| Microscope Shutter Control system | The UNIBLITZ, US | VCM-D1 | VCM-D1 Single Channel CE/UL/CSA Approved Shutter Driver |
| MetaMorph Image software | Universal Imaging Corp., PA | Real-time super-resolution image processing |
References
- Purcell, A. H., Hopkins, D. L. Fastidious xylem-limited bacterial plant pathogens. Annu. Rev. Phytopathol. 34, 131-151 (1996).
- Purcell, A. H. Xylella fastidiosa, a regional problem or global threat. J. Plant Pathology. 79, 99-105 (1997).
- Hopkins, D. L. Xylella fastidiosa: Xylem-limited bacterial pathogen of plants. Annu. Rev. Phytopathol. 27, 271-290 (1989).
- Mattick, J. S. Type IV pili and twitching motility. Annu. Rev. Microbiol. 56, 289-314 (2002).
- Meng, Y., et al. Upstream migration of Xylella fastidiosa via pilus-driven twitching motility. J. Bacteriol. 187, 5560-5567 (2005).
- Li, Y., et al. Type I and type IV pili of Xylella fastidiosa affect twitching motility, biofilm formation and cell-cell aggregation. Microbiology. 153, 719-726 (2007).
- Simpson, A. J. G., et al. The genome sequence of the plant pathogen Xylella fastidiosa. Nature. 406, 151-157 (2000).
- Maier, B., Potter, L., So, M., Long, C. D., Seifert, H. S., Sheetz, M. P. Single pilus motor forces exceed 100 pN. Proc. Natl. Acad. Sci. USA. 99, 16012-16017 (2002).
- Touhami, A., Jericho, M. H., Boyd, J. M., Beveridge, T. J. Nanoscale characterization and determination of adhesion forces of Pseudomonas aeruginosa pili by using atomic force microscopy. J. Bacteriol. 188, 370-377 (2006).
- Skerker, J. M., Berg, H. C. Direct observation of extension and retraction of type IV pili. Proc. Natl. Acad. Sci. USA. 98, 6901-6904 (2001).
- Brown, D. C., Larson, R. S. Improvements to parallel plate flow chambers to reduce reagent and cellular requirements. BMC Immunol. 2, 9 (2001).
- Thomas, W. E., Nilsson, L. M., Forero, M., Sokurenko, E. V., Vogel, V. Shear-dependent 'stick-and-roll' adhesion of type 1 fimbriated Escherichia coli. Mol. Microbiol. 53, 1545-1557 (2004).
- Thomas, W. E., Trintchina, E., Forero, M., Vogel, V., Sokurenko, E. V. Bacterial adhesion to target cells enhanced by shear force. Cell. 109, 913-923 (2002).
- Bahar, O., Fuente, D. L., Burdman, S. Assessing adhesion, biofilm formation and motility of Acidovorax citrulli using microfluidic flow chambers. FEMS Microbiol. Lett. 312, 33-39 (2010).
- Thomas, W. E. Using a laminar flow system to explain shear-enhanced bacterial adhesion. Proceedings of ICMM2005, Third International Conference on Microchannels and Mini-channels. Toronto, Ontario, Canada, , 751-759 (2005).
- Fuente, D. L., et al. Assessing adhesion forces of type I and type IV pili of Xylella fastidiosa bacteria by use of a microfluidic flow chamber. Appl. Environ. Microbiol. 73, 2690-2696 (2007).
- DeLange, P. A., Collins, T. L., Pierce, G. E., Robinson, J. B. PilJ localizes to cell poles and is required for type IV pilus extension in Pseudomonas aeruginosa. Curr Microbiol. 55, 389-395 (2007).
- Fuente, D. L., Burr, T. J., Hoch, H. C. Mutations in type I and type IV pilus biosynthetic genes affect twitching motility rates in Xylella fastidiosa. J. Bacteriol. 189, 7507-7510 (2007).
- Ferandez, A., Hawkins, A. C., Summerfield, D. T., Harwood, C. S. Cluster II che genes from Pseudomonas aeruginosa are required for an optimal chemotactic response. J. Bacteriol. 184, 4374-4383 (2002).
- Cursino, L., et al. Identification of an Operon, Pil-Chp, That Controls Twitching Motility and Virulence in Xylella fastidiosa. Mol. Plant Microbe Interact. 10, 1198-1206 (2011).
- Hazelbauer, G. L., Falke, J. J., Parkinson, J. S. Bacterial chemoreceptors: High-performance signaling in networked arrays. Trends Biochem. Sci. 33, 9-19 (2008).
- Costa, H. S., et al. Plant hosts of Xylella fastidiosa in and near southern California vineyards. Plant Dis. 88, 1255-1261 (2004).
- Shi, X. Y., Dumenyo, C. K., Hernandez-Martinez, R., Azad, H., Cooksey, D. A. Characterization of regulatory pathways in Xylella fastidiosa: genes and phenotypes controlled by algU. Appl. Environ. Microbiol. 73, 6748-6756 (2007).
- Matsumoto, A., Young, G. M., Igo, M. M. Chromosome-Based Genetic Complementation System for Xylella fastidiosa. Appl. Environ. Microbiol. 75, 1679-1687 (2009).
- Davis, M. J., Purcell, A. H., Thomson, S. V. Isolation Media for the Pierce's Disease Bacterium. Phytopathology. 70, 425-429 (1980).
- Xia, Y. N., Whitesides, G. M.
- Chaudhury, M. K., Whitesides, G. M. Direct measurement of interfacial interactions between semispherical lenses and flat sheets of poly-(dimethylsiloxane) and their chemical derivatives. Langmuir. 7, 1013-1025 (1991).
- Cruz, L. F., Parker, J. K., Cobine, P. A., De La Fuente, L. Calcium-enhanced twitching motility in Xylella fastidiosa is linked to a single PilY1 homolog. Appl. Environ. Microbiol. 80, 7176-7196 (2014).

{kind=link}