

Summary
I denna studie var en nano-mikroflödessystem flödeskammare används för att visualisera och funktionellt karakterisera ryckningar motilitet Xylella fastidiosa, en bakterie som orsakar Pierce sjukdom i vinrankor.
Abstract
Xylella fastidiosa är en Gram-negativ icke-flagellated bakterie som orsakar ett antal ekonomiskt viktiga sjukdomar i växter. Ryckningar motilitet erbjuder X. fastidiosa ett medel för långväga inom växt rörelse och kolonisering, bidrar mot patogenicitet i X. fastidiosa. Den ryckningar motilitet X. fastidiosa drivs av typ IV pili. Typ IV pili av Xylella fastidiosa regleras av pilG, en kemotaxi regulator i Pil-CHP operon som kodar för proteiner som är involverade med signaltransduktionsvägar. Att belysa de roller pilG i ryckningar motilitet X. fastidiosa, en pilG med brist mutant Xf ΔpilG och dess komplementära stammen XfΔpilG- C innehållande nativt pilG utvecklades. En mikroflödeskammare integreras med en time-lapse bildregistreringssystemet användes för att observera ryckningar motilitet i XfΔpILG, XfΔpilG- C och dess vilda stammen. Med hjälp av detta registreringssystem, tillåter det långsiktiga rumsliga och tidsmässiga observationer av aggregering, migration av enskilda celler och populationer av bakterier via ryckningar motilitet. X. fastidiosa vild typ och kompletterande XfΔpilG- C-stammen visade typiska ryckningar motilitet egenskaper direkt observerats i mikroflödessystem flödeskammare, medan mutant XfΔpliG uppvisade ryckningar bristfällig fenotypen. Denna studie visar att pilG bidrar till ryckningar motiliteten hos X. fastidiosa. Den mikroflödessystem flödeskammare används som ett medel för att observera ryckningar motilitet.
Introduction
Xylella fastidiosa är en Gram-negativa icke-flagellated, patogen bakterie som orsakar en rad ekonomiskt viktiga grödor sjukdomar, inklusive Pierce sjukdom i vinranka (Vitis vinifera L.) 1,2, 3. Är begränsad till vattenledande xylem Denna bakterie kärl. Infektion av vinrankor orsakar blockering av xylem fartyg och resulterar i vattenstress och näringsbrist 3. Framgångsrik kolonisering beror på förmågan av bakterien för att flytta från det initiala infektionsstället till resten av anläggningen 3. Ryckningar motilitet är ett sätt att flagellar oberoende bakterie rörelse genom förlängning, kvarstad, och indragning av den polära typ IV pili 4 som har präglats i X. fastidiosa 5,6,7.
Ryckningar motilitet har observerats med laser pincett och atomkraftsmikroskopi (AFM) 8,9,10. Med användning av dessa tekniker, twitching motilities genereras av typ IV pilus av N. gonorrhoeae och P. aeruginosa kännetecknades av fl uorescently märkning pili och fånga deras rörelser mikroskopiskt. Även om båda metoderna har detaljerade vidhäftningskraften av enskilda bakterier, förfarandena är komplicerade och tidskrävande 9,10. Mikro fl uidic kamrar användes för att observera långväga migration av enskilda celler såväl som små aggregat av bakteriella celler 5,6. Dessa kammare var utformade som en mikro-nano-kanalen i en platta integrerad med en time-lapse bildregistreringssystem 11,12,13,14. Micro fl uidic kammar enheter erbjuder flera fördelar för att studera rörelse beteende och cell-cell interaktioner av bakterier (i) det ger en integrerad plattform med flera kanaler funktioner; (Ii) det kan undersöka rörelserna och sammansättningar av enstaka celler i nanoskala funktioner av bakterier; (Iii) den möjliggör direkt microscopic bildinspelning av bakterieceller och tidsförlopp analys, (iv) det ger långsiktiga rumsliga och tidsmässiga observationer av individuella och / eller populationer av bakterier i en mikro-miljö; (V) flödeshastigheten för odlingsmedium i en kanal kan styras exakt och (vi) endast en mycket liten volym (1 ml) av odlingsmedium krävs för varje experiment.
Nyligen har mikro fl uidic fl ödet systemet använts för att undersöka beteenden bakterieceller under olika mikromiljöer 14,15,16. Vidhäftningsförmågan och ytan fastsättning av E. coli 15, X. fastidiosa 16, och Acidovorax citrulli 14 till glasytor bedömdes med hjälp av mikro fl uidic kammare. Aggregering och bio fi lm bildning förmedlas av typ IV pili av Acidovorax citrulli analyserades 14. Vidare rörelsen hos A. citrulli observeras under fl öde cILLKOR visat att typ IV-pili kan spela viktiga roller i kolonisering och spridning av A. citrulli i xylem fartyg enligt SAP fl ow villkor. Ryckningar motilities av Pseudomonas aeruginosa och X. fastidiosa celler framgångsrikt observer mot en vätskeström i en mikroflödeskammare 5,6,17. Typ IV pilus bristfällig pilB och pilQ mutanter av X. fastidiosa befanns grunden ändra hastigheten på ryckningar motilitet under fl öde förhållandena i mikro fl uidic enheter 5,6,18. De studier som utförts på bakteriell vidhäftning och rörlighet i mikro fl uidic enheter visade att mikro fl uidic kammare är särskilt lämpliga för att analysera ryckningar motilitet och migrering av pili-medierad bakterier in vitro. Dessa resultat förklarar ryckningar-medierad migration mekanism som underlättar cell cellvidfästning, aggregering och kolonisering inomvärden, så småningom leda till systemisk infektion.
Pil-CHP operonet av X. fastidiosa innehåller pilG, pili, pilJ, piller, chpB och chpC som kodar signaltransduktionsvägar 20. De trans chemoreceptors binder kemiska stimuli i det periplasmatiska domän och aktivera en signalkaskad i deras cytoplasmiska del att slutligen kontrollera bakteriell ryckningar motilitet. I Pil-CHP operonet X. fastidiosa, är en fosfor-transferprotein PilG en homolog till Chey. I E. coli och P. aeruginosa, är Chey svarsregulator i kemotaxi som samverkar med flagmotorproteiner 19, 21. Även om bidragen från Pil-CHP operonet mot virulens i X. fastidiosa undersöktes nyligen 20, roll pilG i kemotaxi operon som svar på miljösignaler och regleras / motortyp IV pili av X. fastidiosa är unclear. Att belysa insikten av kemotaxi regulator pilG i aktiviteten av ryckningar motilitet X. fastidiosa, är en mikro fl uidic kammaren används för att bedöma ryckningar motilitet av X. fastidiosa. Den pilG X. fastidiosa kännetecknas genom att jämföra fenotyper av en deletionsmutant Xf ΔpliG, kompletterande stam XfΔpliG -C och dess vildtyp in vitro. Resultaten belyser roll pilG i ryckningar motilitet X. fastidiosa.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. Perifer Fringe av bakteriekoloni
- Växa X. fastidiosa (Xf) Temecula vildtyp 22, pilG deletionsmutant Xf ΔpliG (med användning av tidigare beskrivna deletion strategi 23), och dess komplementära XfΔpliG -C (med användning av tidigare beskrivna kromosom-baserad genetisk komplemente strategi 24) på PD2-medium agarplattor 25 på 28 ° C i 5-7 dagar.
- Autoklav cellofan (1 x 1 cm 2) i vatten vid 121 ° C (249 ° F) under 15 min. Plocka upp en bit av cellofan, dränera vattnet genom att peka på ett hörn av cellofan på en tom petriskål, försiktigt lägga cellofan över 15% av agarytan och lufttorka.
- Plocka upp enskilda X. fastidiosa kolonier med sterila rundade tandpetare och spot celler aseptiskt på en steriliserad ark av cellofan överlagras på 15% av agarytan i agarplattorna. Inkubera plattans vid 28 ° C under 2-3 dagar.
- Undersök kanten morfologi av kolonierna med hjälp av en dissekera mikroskop med en 2X objektiv och en 10X okularlinsen. Fotografera den perifera kant runt kolonier.
2. Mikroskopi och Mikroflödes Flow Chambers
- Tillverka mikroflödessystem enheter som använder foto litografiska förfaranden som liknar dem som tidigare beskrivits 5,18. Design fyra parallella kanaler med datorstödd design mjukvara på huvudkiselskiva med användning av standard litografiska metoder 26.
- Skapa mikroflödeskammare från kiselskiva mästare med polydimetylsiloxan (PDMS). Pour opolymeriserade PDMS över kiselskivan master och bota det vid 60 ° C under 1 timme. Dra av PDMS replik från skivan master och trimma PDMS replik med ett blad in i en 22 mm x 40 mm som samma storlek på ett täckglas.
- Exponera PDMS replik, ett täckglas (22 x 40 mm 2), och en MicroscOPE slide (51 x 76 mm 2) till luftplasma vid 30 W i 2 min 27. Sandwich PDMS kroppen mellan glaset täckglas och glas mikroskop för att bygga mikro fl uidic kammare.
- Borra ett hål (5,5 mm i diameter) genom PDMS vid varje ände av den mönstrade-kanalen. Skär silikongummislangen i 12-20 cm lång. Infoga en ände av silikongummislang (5,1 mm ytterdiameter, 2,1 mm innerdiameter, 0,8 mm vägg) i vart och öppningsänden av kanalerna i PDMS replik, och försegla den med opolymeriserade PDMS vid 60 ° C under 1 timme.
- Anslut en annan ände av slangen till hullingförsedda änden av plast lueranslutningar. Linda de hopsatta mikrofluidkamrarna med aluminiumfolie och autoklavera dem under 20 min.
- Samla in bakterieceller av X. fastidiosa vildtyp, mutant Xf ΔpliG och kompletterande XfΔpliG -C via skrapning med hjälp av engångs ympa loopar från PD2 mediumplattor. Justera celltäthettill en optisk densitet av 0,05 vid 600 nm i PD2 buljong som beskrivits tidigare 23. Samla in bakteriecellen lösningen i en gastät spruta 1 ml.
- Montera mikrofluidikanordning på ett inverterat mikroskop skede. Anslut ett inloppsrör till Gastät sprutan 5 ml innehåller PD2 buljong. Montera 5 ml Gastäta spruta med sprutpumpar.
- Ansluter utloppsröret till en behållare avfall. Upprätthålla en medelflödeshastighet av 0,2 | j, l min -1 under 30 min för att stabilisera systemet.
- Anslut sidoinloppsrören till en 1 ml Gastäta spruta innehåller bakteriecellen lösning. Spola den bakteriella cellen lösningen genom gummislangen tills kanalen har nåtts. Upprätthålla en medelflödeshastighet av 0,2 l min -1 under ytterligare 30 minuter för att spola obundna celler från kammaren före bildtagning.
- Montera mikroskop slutare under fältet justerade delen av mikroskop för att styra ljuset. Anslut luckan till slutar control systemet och anslut slutarstyrsystemet till datorn.
- Montera en digitalkamera till videoporten av mikroskopet och anslut den till datorn. Kör time-lapse inspelningsprogram, välj "slutare" -funktionen på menyn, och känner automatiskt den installerade slutare som standard i programvaran för att upprätta anslutningar till slutaren med programvaran.
- Välj "digitalkamera" funktion från menyn för time-lapse inspelning programvara för att automatiskt känna igen den digitala kameran som standardinspelningsenhet i programvaran och upprätta kommunikation med den digitala kameran med programvaran.
- Leta bakteriecellerna i en av kanalerna med hjälp av 20X faskontrast optik, sedan byta till 40X objektiv före bildfångst.
- Kör time-lapse inspelningsprogram, välj "bildtagning" funktion med standardparametrar från menyn för att förvärva bilder från mikroskopet.Därefter öppnar "Hämta time-lapse" funktion och ställ in tidsintervallet till 30 sek 5,18,28 löptid för 6-24 h beroende på experiment som behövs för att observera ryckningar motilitet X. fastidiosa 5,18,28. Klicka på "OK" för att starta time-lapse inspelning. Klicka på "Stack funktionen" från menyn, välj "spara som" för att stapla bilderna i destinationsmappen på datorn efter avslutad inspelning.
- För flera kanaler, fånga tidsförlopp bilder från den första kanalen var 30 sekund under 6 timmar. Flytta objektivlinsen hos mikroskopet till nästa kanal för att lokalisera målcellerna. Upprepa time-lapse funktion som beskrivits ovan för att ta bilder i var och en av fyra kanaler sekventiellt om experiment är inställd på att utnyttja fyra kanaler. Fortsätt time-lapse Bildinsamling så länge som tre dagar i följd. Alla experiment utfördes vid rumstemperatur (23 ± 2 ° C).
- Sammanställa time-lapse bilder till envideofil med hjälp av time-lapse inspelning bildprogram. Kör time-lapse inspelning mjukvara, klicka på "Stack-funktionen" i menyn och välj "öppna stack funktion" för att öppna de staplade filer från datorn.
- Starta "Gör filmen" -funktion från stapeln modulen, markera alla bilder och välja "AVI" output format. Klicka på "Spara som" för att spara videofilen i målmappen på datorn.
- Välj de sammanställda filmer från destinationsmappen på datorn och spela upp dem. Då, observera motilitet av de enskilda celler genom den resulterande visualisering av ryckningar motilitet aktiviteten av bakterieceller i den genererade videofiler.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Närvaron av en perifer koloni frans indikativ för typ IV pilus-medierad ryckningar motilitet, observerades i kolonierna av X. fastidiosa vild typ och kompletterande Xf ΔpliG -C-stam (Figur 1). Mutant XfΔpliG emellertid uppvisade inte en frans runt periferin av kolonierna (Figur 1). Time-lapse avbildning av bakterieceller i nanomikroflödesflödeskammare visade att ryckningar motilitet observerades i både vildtyp X. fastidiosa och den komplementära XfΔpliG -C (Supple V1, V3), medan XfΔpilG mutanta celler uppvisade inte ryckningar motilitet genom hela experimentet (Supplemental V2). Celler av mutant XfΔpilG bildade relativt små lösa aggregat i PD2 buljong (Supple V2). I motsats, celler av X. fastidiosa vildtyp och komplementär XfΔpilG- C d eveloped större aggregat i PD2 buljong (Figur 2, (Supplemental V1, V3).

Figur 1:. Den perifera utkanten av bakteriekoloni Colony marginal egenskaper X. fastidiosa från vildtyp, mutant Xf ΔpilG och kompletterande XfΔpilG- C odlas på PD2 agar täckt med en steriliserad ark av cellofan. Med undantag av mutant XfΔpilG alla kolonier uppvisade en perifer frans indikerar typ IV pilus-medierad ryckningar motilitet. Bilderna togs efter 5 dagars odling på odlingsmediet. Förstoring bar, 0,5 mm. Klicka här för att se en större version av denna siffra.
_upload / 53.816 / 53816fig2.jpg "/>
Figur 2: ryckningar motilitet X. fastidiosa celler i nanomikroflödesflödeskammare. Den ryckningar motilitet alla testade stammen celler spelades in under 6 dagar för observation. Bedömningarna genomfördes från tre oberoende videosegment. Förstoring bar, 20 | j, m.
Obs! Ryckningar motilitet X. fastidiosa celler kännetecknas av en enda cell rörelse över glasytor genom förlängningen kvarstad, och indragning av den polära typ IV pili. Den enda cell observerades i migreringen företrädesvis mot en vätskeström i en mikroflödeskammare. Klicka här för att se en större version av denna siffra.
6 / 53816supfig1.jpg "/>
Kompletterande Figur 1: En fyra-kanal mikroflödessystem flödeskammare. Varje kanal med media i och media ut kontakter i varje ände. Klicka här för att se en större version av denna siffra.
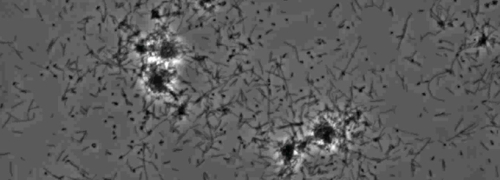
Kompletterande Movie 1: Ryckningar motilitet. (Högerklicka för att ladda ner). Ryckningar i vild-typ X. fastidiosa i en mikroflödessystem flödeskammare.
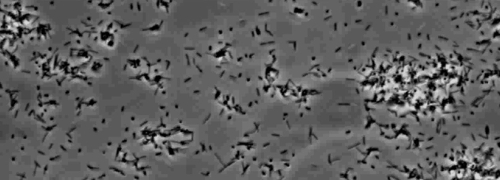
Kompletterande Movie 2: Försämrad twitching motilitet. (Högerklicka för att ladda ner). Motilitet Xf mutant. XfΔpilG observerats i amicrofluidic flödeskammaren.
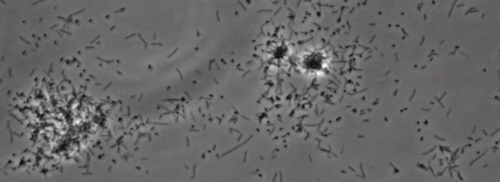
Kompletterande Movie 3: Återställd ryckningar motilitet. (Högerklicka för att ladda ner). Ryckningar motilitet Xf kompletterande stam. XfΔpilG-C observerades i ett mikrofluidflödeskammare.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
I denna studie har vi karaktäriserat rörelsebeteende X. fastidiosa PilG mutant Xf ΔpilG och dess komplementära XfΔpilG- C-stammar i nydesignade flera parallella nano-kanal mikro fl uidic kammare. De nydesignade mikro fl uidic kammare kan ha upp till fyra parallella kammare med 100 um nano-kanalen i bredd jämfört med tidigare konstruktioner med en enda 50 um bred kanal 18. Den förbättrade bredare nano-kanal underlättar införandet av bakterieceller med flödet av media. Dessutom är denna mikrofluidik kammaren 1) enkel att bygga och montera; 2) relativt billiga; och 3) lätt tillämpas på olika experimentella krav. Som ett resultat, denna kammare konstruktion medger långsiktig observation av rörelser bakteriecellerna enligt sorter experimentella mikromiljö.
Stabilisera fl ödet av strömmarna through mikro fl uidic kanal är det kritiska steget för att skapa en intakt flytande-micro fl uidic mikro för observation av rörligheten av bakteriecellerna under olika experimentella miljöer. Spolningen av mikro fl uidic kammare och anslutande slang med media före införandet av bakterieceller är också ett viktigt steg för att stabilisera flödet i kammaren. Emellertid kommer den höga hastigheten av flödet spola bakteriecellerna ut ur kammaren utan att behålla celler under loppet av kammaren. Rätt hastighet av media som strömmar genom mikro fl uidic kanaler behöver justeras med hjälp av en sprutpump. Under experimenten i denna studie, var den strömmande inställd och stabiliserad av pumpen vid 0,2 till 1 | j, l min -1 under minst 30 min före införandet av de bakteriella cellerna in i kanalen. När cellerna infördes i mikro fl uidic kanaler, mediet fl ödet hölls vid 0,2 | j, l mi -1 i 30 till 60 min för att stabilisera systemet och avlägsna nonattached celler. Det är mycket viktigt för att stabilisera flödet och för att hålla bakgrunden klar för att observera rörelsen hos bakteriecellerna. Bilder fångades var 30 sekunder för att bekräfta den ryckningar motilitet aktiviteten hos de införda cellerna upprätthålla en konstant strömningshastighet på 0,2 l min -1. Om försöken att behöva samla in bakteriecellerna i kammaren kan cellerna spolas ut genom att gradvis öka Flödet andelen media 0,2-110 l min -1 genom att justera sprutpumpen hastighet.
De bakteriella cellaktiviteter i mikro fl uidic kammare bedömdes genom inverterat mikroskop med hjälp av 40X faskontrast optik och tidsförlopp bilder som spelats in med en digital kamera, som kontrollerades av bildprogram. Flödeshastigheten och intervalltiden för bildfångst kan justeras i enlighet med experimental krav. Men i de flesta fall, flödeshastighet för ett medium flödeshastighet är inställd på 0,2 | j, l min -1 med tidsförlopp bilder registreras varje 30 sek för pilus-medierad bakterieceller. I andra fall, om mediet fl öde hastigheten ökas till 0,1 l min -1 under experiment beteende bakteriecellerna kommer att registreras genom att fånga bilder varje 10 till 15 sekunder därefter. De tidsförlopp bilder togs varje 30 sek löptid för 6-24 timmar och sparas som en källbildfiler för varje testad stam. Filmerna sedan följs från bilder tagna 6-8 timmar från varje stam, som visade kontrast fenotyper i ryckningar motilities mellan mutant och vild typ / komplement stammar (Supple V1, V2, V3).
Betydelsen av nano micro fl uidic kammar enheter över makroskala parallellplatt fl öde kammare omfattar direkt undersökning av rörelserna och ansamlingar av sjungale celler av bakterier. I tillägg till låg kostnad 5 och låga krav på reagens- och provvolym, fördelarna med mikro fl uidic kamrarna är lättheten att konstruktion av en fl öde mikromiljö för bakterieodling och noggrann kontroll över fl uid fl öde hastighet. De många parallella kanaler tillåta observation av differentialbakteriestammar i en enda experimentet som ger kompatibla data för analys. Flagellära oberoende ryckningar motilitet av bakterierna med den polära pili är särskilt lämplig för analys i denna nano-mikro fl uidic kammaren. Detta är dock micro fl uidic kammare mindre lämpliga för flagellar beroende bakterie rörelse, i vilken rörelsen av bakterier är oftast alltför snabb och uppvisar slumpmässiga riktningar. Denna begränsning ibland kan äventyras genom att justera medelflödeshastigheten till 0,05 l min -1 och ändra tidsförlopp bild avskiljningsgrad vid varje 1 till 2 sek under 1 timme till Analyze rörelser de flagellar-medierad bakterier i nanoskala miljö.
Mikroflödeskammar enheter som används häri ger direkt visuell bevis för funktionell utvärdering av PilG ansvarig för rörelsebeteende in vitro. Dessutom, denna studie visar också att cell till cell aggregering genom ryckningar motilitet är viktigt för biofilmbildning, signal- och patogenicitet i X. fastidiosa. Visualisering av bakterier ryckningar motilitet av mikroflödessystem enheter ger en ny metod för att studera geners funktion involverar beteende i bakterier som inte är lätt att mäta med andra analysmetoder. Detta tillvägagångssätt kan tillämpas på andra bakteriella system. Den mikroflödessystem kammare enheter ger ett flödessystem för att karakterisera den fysiologiska beteende av bakterieceller i samband med cell-cell bilagor, cellsamlingar, rörelsemönster och biofilmbildning.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Författarna förklarar att de inte har några konkurrerande ekonomiska intressen.
Acknowledgments
Denna studie stöddes av United States Department of Agriculture, Agricultural Research Service. firmanamn eller kommersiella produkter i detta dokument är endast nämns för att tillhandahålla specifik information och innebär inte någon rekommendation av Förenta staternas Department of Agriculture. USDA är en lika möjligheter leverantör och arbetsgivare.
Materials
| Name | Company | Catalog Number | Comments |
| Biology materials | |||
| X. fastidiosa (Xf) Temecula wild type | Costa, H. S., et al., 2004 22 | ||
| pilG deletion mutant XfΔpliG | Shi, X. Y., et al., 2007 26 | ||
| pilG complementary strain XfΔpliG-C | Davis, M. J., et al. 1998 23 | ||
| Physical materials and equipment | |||
| Disposable inoculating loops | VWR international, Radnor, PA | #22-363-607 | quantitative procedures such as bacterial collection |
| Polydimethylsiloxane (PDMS) | Dow Corning Corporation | #0002709226 | Sylgard 184 silicone Elastomeric Kits |
| AmScope MD2000 digital camera | AmScope, Irvine, CA | SE305R-AZ-E | Image, video recording and measurement |
| Tubes line | Edgewood, NY | #T4300 | Connected to the syringe and microfluidic chamber |
| Plastic luer connectors | Edgewood, NY | Connected to the syringe and microfluidic chamber | |
| Syringe pumps | Pico Plus, Harvard Apparatus, MA | #702209 | The flow rate can be adjusted while the pump is running. |
| Syringes | Gastight, Hemilton Company, Reno, NV | #1005 | Provide the flowing broth |
| Inverted Olympus IMT-2 microscope | Olympus | IMT-2 FLuoro PHase | Image observation and recording |
| SPOT-RT digital camera | Diagnostic Instruments, Inc., MI | RT230 | Image, video recording and measurement |
| Microscope Shutter | The UNIBLITZ, US | #LS2T2 | Control camera’s exposure time |
| Microscope Shutter Control system | The UNIBLITZ, US | VCM-D1 | VCM-D1 Single Channel CE/UL/CSA Approved Shutter Driver |
| MetaMorph Image software | Universal Imaging Corp., PA | Real-time super-resolution image processing |
References
- Purcell, A. H., Hopkins, D. L. Fastidious xylem-limited bacterial plant pathogens. Annu. Rev. Phytopathol. 34, 131-151 (1996).
- Purcell, A. H. Xylella fastidiosa, a regional problem or global threat. J. Plant Pathology. 79, 99-105 (1997).
- Hopkins, D. L. Xylella fastidiosa: Xylem-limited bacterial pathogen of plants. Annu. Rev. Phytopathol. 27, 271-290 (1989).
- Mattick, J. S. Type IV pili and twitching motility. Annu. Rev. Microbiol. 56, 289-314 (2002).
- Meng, Y., et al. Upstream migration of Xylella fastidiosa via pilus-driven twitching motility. J. Bacteriol. 187, 5560-5567 (2005).
- Li, Y., et al. Type I and type IV pili of Xylella fastidiosa affect twitching motility, biofilm formation and cell-cell aggregation. Microbiology. 153, 719-726 (2007).
- Simpson, A. J. G., et al. The genome sequence of the plant pathogen Xylella fastidiosa. Nature. 406, 151-157 (2000).
- Maier, B., Potter, L., So, M., Long, C. D., Seifert, H. S., Sheetz, M. P. Single pilus motor forces exceed 100 pN. Proc. Natl. Acad. Sci. USA. 99, 16012-16017 (2002).
- Touhami, A., Jericho, M. H., Boyd, J. M., Beveridge, T. J. Nanoscale characterization and determination of adhesion forces of Pseudomonas aeruginosa pili by using atomic force microscopy. J. Bacteriol. 188, 370-377 (2006).
- Skerker, J. M., Berg, H. C. Direct observation of extension and retraction of type IV pili. Proc. Natl. Acad. Sci. USA. 98, 6901-6904 (2001).
- Brown, D. C., Larson, R. S. Improvements to parallel plate flow chambers to reduce reagent and cellular requirements. BMC Immunol. 2, 9 (2001).
- Thomas, W. E., Nilsson, L. M., Forero, M., Sokurenko, E. V., Vogel, V. Shear-dependent 'stick-and-roll' adhesion of type 1 fimbriated Escherichia coli. Mol. Microbiol. 53, 1545-1557 (2004).
- Thomas, W. E., Trintchina, E., Forero, M., Vogel, V., Sokurenko, E. V. Bacterial adhesion to target cells enhanced by shear force. Cell. 109, 913-923 (2002).
- Bahar, O., Fuente, D. L., Burdman, S. Assessing adhesion, biofilm formation and motility of Acidovorax citrulli using microfluidic flow chambers. FEMS Microbiol. Lett. 312, 33-39 (2010).
- Thomas, W. E. Using a laminar flow system to explain shear-enhanced bacterial adhesion. Proceedings of ICMM2005, Third International Conference on Microchannels and Mini-channels. Toronto, Ontario, Canada, , 751-759 (2005).
- Fuente, D. L., et al. Assessing adhesion forces of type I and type IV pili of Xylella fastidiosa bacteria by use of a microfluidic flow chamber. Appl. Environ. Microbiol. 73, 2690-2696 (2007).
- DeLange, P. A., Collins, T. L., Pierce, G. E., Robinson, J. B. PilJ localizes to cell poles and is required for type IV pilus extension in Pseudomonas aeruginosa. Curr Microbiol. 55, 389-395 (2007).
- Fuente, D. L., Burr, T. J., Hoch, H. C. Mutations in type I and type IV pilus biosynthetic genes affect twitching motility rates in Xylella fastidiosa. J. Bacteriol. 189, 7507-7510 (2007).
- Ferandez, A., Hawkins, A. C., Summerfield, D. T., Harwood, C. S. Cluster II che genes from Pseudomonas aeruginosa are required for an optimal chemotactic response. J. Bacteriol. 184, 4374-4383 (2002).
- Cursino, L., et al. Identification of an Operon, Pil-Chp, That Controls Twitching Motility and Virulence in Xylella fastidiosa. Mol. Plant Microbe Interact. 10, 1198-1206 (2011).
- Hazelbauer, G. L., Falke, J. J., Parkinson, J. S. Bacterial chemoreceptors: High-performance signaling in networked arrays. Trends Biochem. Sci. 33, 9-19 (2008).
- Costa, H. S., et al. Plant hosts of Xylella fastidiosa in and near southern California vineyards. Plant Dis. 88, 1255-1261 (2004).
- Shi, X. Y., Dumenyo, C. K., Hernandez-Martinez, R., Azad, H., Cooksey, D. A. Characterization of regulatory pathways in Xylella fastidiosa: genes and phenotypes controlled by algU. Appl. Environ. Microbiol. 73, 6748-6756 (2007).
- Matsumoto, A., Young, G. M., Igo, M. M. Chromosome-Based Genetic Complementation System for Xylella fastidiosa. Appl. Environ. Microbiol. 75, 1679-1687 (2009).
- Davis, M. J., Purcell, A. H., Thomson, S. V. Isolation Media for the Pierce's Disease Bacterium. Phytopathology. 70, 425-429 (1980).
- Xia, Y. N., Whitesides, G. M.
- Chaudhury, M. K., Whitesides, G. M. Direct measurement of interfacial interactions between semispherical lenses and flat sheets of poly-(dimethylsiloxane) and their chemical derivatives. Langmuir. 7, 1013-1025 (1991).
- Cruz, L. F., Parker, J. K., Cobine, P. A., De La Fuente, L. Calcium-enhanced twitching motility in Xylella fastidiosa is linked to a single PilY1 homolog. Appl. Environ. Microbiol. 80, 7176-7196 (2014).

{kind=link}