

Summary
ويتم تنظيم الخلايا البكتيرية مكانياً عاليا. لمتابعة هذه المنظمة على مر الزمن في بطء تزايد الخلايا إكسانثوس ميكسوكوككوس ، قد وضعت إعداد لتصوير خلية يعيش الأسفار مع ارتفاع القرار الزمانية المكانية على مدى عدة أجيال. باستخدام هذا الأسلوب، يمكن تحديد الديناميات الزمانية المكانية من البروتينات الهامة للعزل الصبغي وانقسام الخلية.
Abstract
الأسفار خلية يعيش تصوير الخلايا البكتيرية طريقة رئيسية في تحليل ديناميات المكانية والزمانية للبروتينات والصبغيات الكامنة وراء أحداث دورة الخلية المركزية. ومع ذلك، التصوير هذه الجزيئات في البكتيريا بطيئة النمو يمثل تحديا بسبب فوتوبليتشينج فلوروفوريس ولالضيائيه أثناء الحصول على الصور. هنا، يمكننا وصف بروتوكول بسيط للتحايل على هذه القيود في حالة إكسانثوس ميكسوكوككوس (التي لديها وقت جيل ح 4-6). وتحقيقا لهذه الغاية، م. إكسانثوس خلايا تزرع في التي تحتوي على مغذيات أجار وسادة سميكة في بيئة رطبة التحكم في درجة الحرارة. في ظل هذه الظروف، علينا أن نحدد الوقت مضاعفة الخلايا الفردية باتباع نمو الخلايا المفردة. وعلاوة على ذلك، العمليات الخلوية الأساسية مثل العزل الصبغي وانقسام الخلية يمكن تصويرها بالتصوير خلية يعيش الأسفار من الخلايا التي تحتوي على بروتينات علامة المسمى فلوريسسينتلي ذات الصلة مثل فرع يفب، فتسز-التجارة والنقل، وبومكس مشري على مدى عدة دورات الخلية. بعد ذلك، تتم معالجة الصور المكتسبة لتوليد المونتاج و/أو الأفلام.
Introduction
وتنظم الخلايا البكتيرية مكانياً شديدة مع العديد من البروتينات إضفاء الطابع المحلي دون تناسق داخل مقصورات الهاتف الخلوي1،2،،من34. هذا التعريب ديناميكية للغاية في كثير من الأحيان، ويتغير مع مرور الوقت استجابة لمنبهات دورة الخلية أو إشارات خارجية. على قدم المساواة، والكروموسوم البكتيري مكانياً عالية تنظم مع فرادى المكاني يجري وضعه لمواقع سوبسيلولار محددة قبل وأثناء عملية الفصل5. هذه المنظمة المكانية الدينامية مهمة للنمو وشعبة، تنظيم دورة الخلية، والتمايز، وحركية، توصيل الإشارة، فضلا عن منظمة كروموسوم والعزل؛ وهكذا، أنه يؤثر على وظيفة البكتيرية أساسا جميع جوانب.
ويتم يجري تحليل ديناميات الزمانية المكانية لهذه العمليات الخلوية في مجموعة متنوعة من مختلف الأنواع البكتيرية مع الإشريكيّة القولونية، و نجحت عصية، والكوليرا، و كريسسينتوس كاولوباكتير خدمة هامة نموذج الكائنات الحية. ومع ذلك، هذه الأنواع الأربعة تشمل فقط مجموعة صغيرة من التنوع الجرثومي الهائل، وربما لم يكن مفاجئاً نظراً للمسافة الكبيرة النشوء والتطور بين هذه الأنواع، آليات التنظيم والاستقطاب الخلوية المختلفة في هذه البكتيريا. وهذا يثير الحاجة إلى دراسة الأنواع البكتيرية إضافية ليتمكن من استخلاص المبادئ العامة التي تستند إليها ديناميات الزمانية المكانية للخلايا البكتيرية في نهاية المطاف.
سلبية الغرام الدلتا-بروتيوباكتيريوم م. إكسانثوس هو كائن نموذج في دراسة السلوكيات الاجتماعية والتعاون في البكتيريا6. م. إكسانثوس جرثومات صارمة وحضور العناصر المغذية، وأنها تشكل المستعمرات التي تنتشر الخلايا إلى الخارج بدرجة عالية من التنسيق ويحتشدون الأزياء ويفترس على سائر الكائنات الحية الدقيقة7. استجابة لمجاعة المغذيات، خلايا الشروع في برنامج إنمائي أن النتائج في تشكيل الهيئات الاثمار التي تتكون من آلاف من الخلايا، وداخل، التفريق بين الخلايا متحركة على شكل قضيب كروية الجراثيم مثنوية8. كلا النوعين من السلوكيات، أي، يحتشدون وتشكيل الهيئة الاثمار، يتم تنفيذها فقط من الخلايا التي يتم وضعها على سطح صلب. وعلاوة على ذلك، تحت كل الظروف المغذيات، الخلايا المشاركة في العمليات التي تتضمن اتصالات مباشرة خلية--بما في ذلك تبادل البروتينات الدهنية الغشاء الخارجي قد حفز حركية أو تعمل كالسموم في المتلقي9،10 ، والخلايا تبادل لبس11، تنشيط حركية قبل والانسيابيه في المجاورة12، وبين الخلايا مما يشير إلى جانب خلية سطح ارتساء إشارات بروتين13،14.
في الآونة الأخيرة، م. إكسانثوس كما أصبح كائن نموذج لدراسة الآليات الأساسية لحركية والبند15، وانقسام الخلايا16،،من1718، ومنظمة كروموسوم19 ،،من2021. خطوات حاسمة في م. إكسانثوس دورة الخلية قد تم تحليلها بالتفصيل بالأسفار مجهرية باستخدام لقطة مفاجئة صور أو تسجيلات الوقت الفاصل بين القصير على سلالات تحمل البروتينات ذات الصلة المسمى فلوريسسينتلي16، 1718،،،من1920. ومن الناحية المثالية، ينبغي اتباع العديد من الخلايا مع القرار خلية واحدة fluorescence خلية يعيش التصوير لدورة الخلية كاملة واحدة على الأقل للحصول على بيانات كمية قوية على معلمات دورة الخلية. ومع ذلك، وهذا يشكل تحديا بالنسبة ل م. إكسانثوس نظراً لوقت طويل نسبيا جيل ح 4-6 تحت ظروف المختبر القياسية وسبب فوتوبليتشينج فلوروفوريس ولالضيائيه أثناء الحصول على الصور.
هنا، يمكننا وصف بروتوكول متابعة م. إكسانثوس الخلايا مع القرار خلية مفردة بالأسفار يعيش خلية التصوير لمالا يقل عن 24 ساعة وتغطي عدة دورات الخلية. الأهم من ذلك، خلال بروتوكول كامل، خلايا تمسك جهاز pad أجار وفي إغلاق الاتصال السماح للأنشطة التي تعتمد على الاتصال الأساسية لنمط الحياة الاجتماعية م. إكسانثوس. البروتوكول أيضا يسمح للمستخدمين لرصد الشكل والحجم، والانقسامات، والمسابير الفلورسنت عالية دقة زمنية ومع القرار خلية مفردة، وهكذا، يمكن التحديد الكمي لتقلب خلية إلى خلية والارتباطات لإحداث دورة الخلية.
Protocol
1-إعداد ونمو م. إكسانثوس سلالات
ملاحظة: انظر الجدول 1 و الجدول 2.
- تعد 1% كاستون مرق (ضريبة تحويل العملة) نمو متوسطة 1% (w/v) البنكرياس الخلاصة من الكازين (مثلاً، كاستون بكتو)، 10 مم تريس-HCl pH 8.0 ملم 1 خ2بو4 درجة الحموضة 7.6، 8 مم MgSO422، تكملها كاناميسين (50 ميكروغرام/مل) أو أوكسي تتراسيكيلين (10 ميكروغرام/مل). إضافة مع الجنتامايسين (10 ميكروغرام/مل) لجميع وسائل الإعلام الحد من خطر التلوث بالبكتريا الأخرى، منذ ذلك الحين م. إكسانثوس الخلايا تقاوم ذلك بطبيعة الحال.
- تلقيح 5 مل من 1% ضريبة تحويل العملة التي تحتوي على ذات الصلة antibiotic(s) مع مستعمرة البرية الطازجة نمت واحد اكتب DK1622 (WT) 23، SA4420 (Δمجلة)24، SA4797 (Δمجلة، Δبومكس/PpomZ مشري-بومكس )16، SA8241 (Δمجلا، فتسز+/PناتftsZ-التجارة والنقل)، أو SA4749 (Δمجلا، فرع+/Pناتفرع-يفب) في صباح اليوم الأول.
- ريسوسبيند واحد م. إكسانثوس مستعمرة في 500 ميليلتر من 1% ضريبة تحويل العملة وتستكمل مع المضادات الحيوية في أنبوب عقيم ونقل تعليق كامل إلى 50 مل قارورة Erlenmeyer الذي يحتوي على 5 مل من 1% ضريبة تحويل العملة.
ملاحظة: استخدم قارورة Erlenmeyer مع 10 مرات حجم الثقافة لضمان أيرياتيون كافية والنمو الأمثل.
- ريسوسبيند واحد م. إكسانثوس مستعمرة في 500 ميليلتر من 1% ضريبة تحويل العملة وتستكمل مع المضادات الحيوية في أنبوب عقيم ونقل تعليق كامل إلى 50 مل قارورة Erlenmeyer الذي يحتوي على 5 مل من 1% ضريبة تحويل العملة.
- تنمو الخلايا لثمانية أجيال (حوالي 40-48 ح مع وقت جيل من ح 4-6) في 32 درجة مئوية، والهز 220 لفة في الدقيقة، في الظلام. الحفاظ على الخلايا في مرحلة النمو الأسى (OD550 < 1.2) ومنعهم من الوصول إلى مرحلة ثابتة. إذا لزم الأمر، تمييع الخلايا إلى الطازجة 1% ضريبة تحويل العملة المتوسطة التي تحتوي على antibiotic(s) ذات الصلة التطوير التنظيمي550 من 0.1-0.2.
ملاحظة: التطوير التنظيمي أمثل550 للفحص المجهري خلية مفردة هو 0.5-0.7. في هذا التطوير التنظيمي550، يوجد عدد كاف من الخلايا كل صورة للسماح للقياس الكمي، فضلا عن التحليل الإحصائي لمعلمات الخلوية.
2-إعداد عينات الفحص المجهري
ملاحظة: توضع على ساترة مجهر الخلايا ليتم عرضه بواسطة الفحص المجهري وثم تغطيها جهاز pad [اغروس] الذي يحتوي على العناصر الغذائية. التصاق ساترة بلاستيك أو دعم الإطار المعدني لتوفير الميكانيكية. استعدادا للفحص المجهري، لوحة كبيرة من agarose/TPM/0.2% 1% ضريبة تحويل العملة وينبغي إعداد مقدما كما هو موضح في الخطوات 2.1-2.3. يرجى أيضا الرجوع إلى الجدول للمواد اللازمة للمنتجات المحددة المستخدمة هنا.
- إعداد 500 مل TPM المخزن المؤقت (10 ملم تريس-HCl درجة الحموضة 7.6 مم 1 خ2بو4 درجة الحموضة 7.6، 8 مم MgSO4) وتعقيم اﻷوتوكﻻف أو عامل تصفية باستخدام عامل تصفية أعلى زجاجة.
ملاحظة: يمكن تخزين المخزن المؤقت العقيمة لعدة أشهر في درجة حرارة الغرفة. - إعداد 1% [اغروس] مجهرية محلول يحتوي على 0.2% ضريبة تحويل العملة (ميكس 1 غرام من [اغروس] مع 80 مل من المخزن المؤقت TPM و 20 مل من 1% ضريبة تحويل العملة المتوسطة). الحرارة في فرن ميكروويف حتى [اغروس] المنصهر.
ملاحظة: 0.2% ضريبة تحويل العملة كاف للسماح للخلايا النمو ومنع المجاعة. سيؤدي إلى تركيزات أعلى من ضريبة تحويل العملة في الأجلين المتوسط والفحص المجهري في الأسفار خلفية عالية. - ملء طبق بيتري مع [اغروس] المنصهر بسمك 0.5 سم (لساحة 11.5 سم × 11.5 سم طبق بيتري، ما يقرب من 60 مل من المنصهر [اغروس] مطلوب) وندعه يبرد وصولاً إلى درجة حرارة الغرفة.
ملاحظة: يمكن تخزين لوحة المفاتيح [اغروس] في 4 درجات مئوية في بيئة رطبة لمدة تصل إلى 2 يوما.- قبل الحارة في لوح agarose/TPM/0.2% 1% ضريبة تحويل العملة عند 32 درجة مئوية على الأقل 15 دقيقة قبل استخدامها.
ملاحظة: إعداد الخلايا للفحص المجهري، اتبع الخطوات 2.4-2.8.
- قبل الحارة في لوح agarose/TPM/0.2% 1% ضريبة تحويل العملة عند 32 درجة مئوية على الأقل 15 دقيقة قبل استخدامها.
- ضع ساترة زجاجية معقمة (60 مم × 22 مم، سمك: 0.7 مم) في إطار بلاستيكية أو معدنية قد حفرة في الوسط (الشكل 1أ)؛ هذا الإطار بمثابة دعم ميكانيكية ساترة رقيقة، ويساعد على الحد من الانجراف خلال الفحص المجهري. إصلاح ساترة للإطار مع الشريط.
- لإعداد الإطار، قص خارج إطار 75 مم × 25 مم من لوحة معدنية سميكة 1 ملم، ثم قطع حفرة حجم مناسب (20 مم × 30 مم في هذه التجربة) في الوسط.
- إضافة 10-20 ميليلتر من نمت أضعافاً مضاعفة م. إكسانثوس الخلايا على ساترة.
- أضف الفلورسنت 0.5 ميكرومتر الجزئي المجالات الخرز الصغير كعلامات الاعتماد في الخلايا لتبسيط تتبع الخلايا أو البروتينات في الوقت الفاصل بين التسجيلات.
- يضعف الجزئي المجالات الخرز الصغير 1: 100 في المخزن المؤقت TPM وتخزينها في 4 درجات مئوية لتصل إلى عدة أشهر. يهز جيدا قبل الاستخدام وإضافة 5-10 ميليلتر من الجزئي المجالات الخرز الصغير المخفف للخلايا.
ملاحظة: هنا الجزئي المجالات الخرز الصغير التي الفلورية في جميع الأزرق المشتركة والأخضر والأصفر والأحمر واستخدمت قنوات الفلورسنت.
- يضعف الجزئي المجالات الخرز الصغير 1: 100 في المخزن المؤقت TPM وتخزينها في 4 درجات مئوية لتصل إلى عدة أشهر. يهز جيدا قبل الاستخدام وإضافة 5-10 ميليلتر من الجزئي المجالات الخرز الصغير المخفف للخلايا.
- قطع وسادة صغيرة بحجم ساترة لوحة ضريبة تحويل العملة agarose/TPM/0.2% 1% قبل حرارة كبيرة تقريبا ثم ضعه أعلى الخلايا (الشكل 1ب). ضع ساترة من أعلى 1% agarose/TPM/0.2% ضريبة تحويل العملة [اغروس] وسادة لمنع التبخر والحفاظ على الخلايا في بيئة رطبة.
ملاحظة: ساترة وحدها سوف يمنع التبخر كبيرة على الأقل 2 ح. لفترة أطول من الوقت الفاصل بين التسجيلات، ينبغي مختومة agarose/TPM/0.2% 1% ضريبة تحويل العملة ساندويتش لوحة وساترة مع الفيلم البرافين لمنع التبخر. - احتضان عينة مجهرية في 32 درجة مئوية لمدة 15-20 دقيقة للسماح للخلايا نعلق على الجزء السفلي من لوحة المفاتيح [اغروس]. ثم بدء تشغيل تسجيلات الوقت الفاصل بين الفحص المجهري.
3-المجهر الإعداد واكتساب الوقت الفاصل بين
ملاحظة: وضعت البروتوكول الموصوفة هنا من أجل مجهر مقلوب ويديفيلد مع ضبط تلقائي للصورة، 100 X/1.30 NA نفط PH3 الهدف X، Y المزودة بمحركات المرحلة، كاميرا سكموس، ومصدر الضوء، مرشحات للأخضر-فلوري، الفلورسنت الأحمر أو الأصفر فلوري تسيطر البروتينات، ودرجة حرارة غرفة الحضانة. وتبقى هذه الدائرة الخلايا محمية من الضوء وفي درجة حرارة ثابتة.
- قبل الحرارة غرفة الحضانة والمجهر إلى 32 درجة مئوية ح ~ 1-2 قبل البدء بالفحص المجهري.
ملاحظة: اعتماداً على تركيب المجهر، قد يستغرق تدفئة أطول. التدفئة قبل أمر ضروري للحد من الانجراف واستقرار نظام مراقبة ضبط تلقائي للصورة. - التبديل في المجهر والبدء برنامج حاسوبي لمراقبة المجهر. حدد الهدف الصحيح والمرايا الصحيح والمرشحات للحصول على المرحلة الصور على النقيض، فضلا عن صور من البروتينات الفلورية الخضراء أو الفلورسنت الأحمر أو الأصفر فلوري.
ملاحظة: يتم تزويد مجهر عادة مع برامج مفضلة لاقتناء مجهر مراقبة والصورة. (انظر الجدول للمواد) استخدمت هنا برمجيات متاحة تجارياً للتحكم في اقتناء مجهر والصورة. - أضف قطره زيت الغمر عالية الجودة على العدسة للهدف وإلى الجزء السفلي من نموذج المحتضنة مسبقاً عند 32 درجة مئوية. وضع الهدف في ممكن أدنى Z-موقف لتجنب إتلاف العدسة الهدف عندما يتم وضع العينة على مرحلة المجهر. ضع الإطار المعادن مع العينة على خشبة المسرح المجهر، ومع "الجانب الثقب" نحو تحقيق الهدف. ربط النموذج بشكل أمن في حامل المرحلة.
- التركيز على الخلايا عن طريق تحريك المرحلة في الاتجاه Z في أقرب إلى الهدف. نقل المرحلة أبطأ عند قطرات النفط على الجانب السفلي عينة وجعل العدسة الهدف الاتصال. الانتقال المرحلة في X/الاتجاه Y حتى عدة خلايا مفردة مرئية في منطقة العرض، عندما تكون الخلايا في المستوى البؤري. تأكد من أن ميكروسفيري نيون واحد على الأقل في منطقة عرض لمحاذاة الصور المكتسبة لاحقاً.
ملاحظة: تحت الظروف المثلى، ينبغي التوصل بكثافة خلية من خلايا 15-30 كل منطقة للعرض (2,048 x 2,048 بكسل أو 133.1 س 133.1 ميكرومتر). - افتح معالج متعدد الأبعاد اقتناء برمجيات التحكم المجهر لإعداد تجربة مرور وقت يسمح المجهر للحصول على الصور في أطوال موجية متعددة ومواقف المرحلة إذا لزم الأمر.
- في علامة التبويب الرئيسية تنشيط Timelapse و أطوال موجية متعددة. سوف تظهر علامات تبويب إضافية على الجانب الأيسر من النافذة.
- انقر فوق علامة التبويب حفظ و حدد الدليل لتحديد مجلد فارغ على القرص الثابت في الكمبيوتر حفظ الصور المكتسبة. تنشيط اسم قاعدة الزيادة في حالة وجود الملف للتأكد من أن مجموعات متتالية عدم الكتابة فوق تلك السابقة. ثم تعطي التجربة اسماً مع التاريخ واسم السلالة أو العنوان للتجربة.
- انقر فوق علامة التبويب Timelapse لضبط الوقت الفاصل بين المعلمات. تعيين المدة إلى 24 ساعة وتعيين الفاصل الزمني إلى 20 دقيقة. سيتم تغيير عدد من النقاط الزمنية تلقائياً.
ملاحظة: الفترة الزمنية المثلى تعتمد على التجربة ووظيفة الخلوية يتم تحليلها. صورة متكررة عمليات الشراء قد يسبب فوتوبليتشينج. وبالتالي، يجب إيجاد مفاضلة بين الأزمنة وفوتوبليتشينج تجريبيا. في وقت تضاعف من 4-6 ح، الصور يمكن بسهولة الحصول عليها عند فاصل زمني لمدة 5 دقائق (أو حتى أصغر فترات إذا رغبت في ذلك) الطوري. إذا كان الفحص المجهري الأسفار على مدى دورة وقت 24 ساعة هو المطلوب ينبغي تسجيل الصور في فاصل زمني من حوالي 15-30 دقيقة. - انقر فوق علامة التبويب أطوال موجية حدد عدد الأطوال الموجية للحصول لكل صورة في كل نقطة من الوقت عن طريق تغيير العدد.
ملاحظة: لكل طول موجي، سوف تظهر علامة تبويب جديدة على الجانب الأيسر من اقتناء متعدد الأبعاد " سيتم الحصول على معالج والاطوال الموجية بالترتيب من الأعلى إلى الأسفل. لكل طول موجي، يمكن تعديل الإعدادات اقتناء كل على حدة. - انقر فوق علامة التبويب طول الموجه الأولى من الأعلى. حدد مرحلة التباين في القائمة المنسدلة الإضاءة . حدد 100 مللي ثانية التعرض وحدد نقطة كل الوقت في الحصول على القائمة المنسدلة. تعطيل الكشف التلقائي عن طريق اختيار ابدأ في القائمة المنسدلة.
- كرر الخطوة 3.5.5 لكل طول موجي الذي يحتاج إلى الحصول عليها في كل نقطة في الوقت. للإعداد التجريبية والبروتينات المسمى فلوريسسينتلي الموصوفة هنا، استخدم المعلمات التالية ل التعرض: ms 250 للبروتينات الانصهار مشري و 200 مرض التصلب العصبي المتعدد للبروتينات الانصهار يفب ms 1,000 للبروتينات فيوجن التجارة والنقل.
ملاحظة: إعدادات الإضاءة الأمثل لكل سلالة وبروتين فلوري تحدد مسبقاً عن طريق تغيير كثافة المصباح ووقت اكتساب صورة لكل طول موجي. مرات الحصول على صورة طويلة جداً سوف زيادة تأثير سمي ضيائي وتؤدي في النهاية إلى وفاة القبض والخلية النمو. ولذلك، ينبغي تحقيق مفاضلة بين سلامة الجودة وخلية الصورة. - الحصول على الصور من مواقف المرحلة متعددة لزيادة عدد الخلايا التي تم تسجيلها في التجربة نفسها.
- للحصول على الصور من مواقف المرحلة متعددة، تنشيط مواقف المرحلة متعددة في علامة التبويب الرئيسية . ثم انقر فوق علامة التبويب المرحلة وانقر فوق الزر حية لإلقاء نظرة على مجال الرؤية.
- نقل المرحلة في الاتجاه-X/Y حتى منطقة الاهتمام (ROI) في مجال الرؤية. حفظ إحداثيات س وص-بواسطة النقر فوق "+" في علامة التبويب مرحلة نقل المرحلة مرة أخرى في الاتجاه-س/ص حتى يتم العثور العائد على الاستثمار جديد وحفظ الإحداثيات مرة أخرى بواسطة النقر فوق "+". تستمر حتى يتم حفظ العدد المطلوب من المناطق.
ملاحظة: في حالة الحصول على الصور الفلورية، تأكد من أن المناطق ذات الاهتمام (رويس) ليست قريبة جداً من بعضها البعض لتقليل الضيائية.
- تحقق مرة أخرى أن الخلايا في التركيز عن طريق النقر على حفظ X-و Y-المواقف المختلفة وبدء جهاز ضبط تلقائي للصورة بواسطة النقر فوق عقد الاتحاد الآسيوي لإبقاء المحفوظة Z-الموقف ثابت على مدى التجربة.
- بدء التسجيلات الوقت الفاصل بين في برامج التحكم المجهر بالنقر فوق الحصول على في معالج اقتناء متعدد الأبعاد .
ملاحظة: سوف تظهر نافذة واحدة لكل طول موجي التي يتم الحصول عليها، وسوف تظهر نافذة إضافية يظهر عدد النقاط المكتسبة بالوقت والوقت حتى الحصول على الصورة التالية. - تحقق من أن الخلايا لا تزال في التركيز بعد قليل أول المرة-النقاط في الوقت الفاصل بين التسجيلات تحقيق أقصى قدر من جودة الصور وإعادة تركيز إذا لزم الأمر.
4-جيل من الوقت الفاصل بين الأفلام ومحاذاة الصورة
ملاحظة: تتوفر العديد من حزم البرامج التجارية والمجانية للحصول على الصور، وتحليل الصور. يمكننا استخدام برمجيات متاحة تجارياً (انظر الجدول للمواد) مع العديد من الإضافات المثبتة مسبقاً وأدوات إضافية.
- حفظ الصور الفردية من الوقت الفاصل بين التسجيلات على جهاز كمبيوتر يحتوي على تركيب البرمجيات التحليل/تجهيز الصورة.
- بدء تشغيل البرامج وفتح الصور مكدس بواسطة النقر فوق "مراجعة بيانات الأبعاد" | قم بتحديد ملف قاعدة | حدد الدليل. قم بفتح المجلد مع البيانات متعددة الأبعاد. التحقق من البيانات وانقر فوق طريقة العرض؛ سيظهر dataset كصور واحدة من الوقت نقطة واحدة حتى النهاية. تنشيط الطول الموجي (من أجل إنشاء مكدس)، حدد كافة الصور التي ينبغي أن تكون في بنية تخزين العناصر ثم انقر فوق تحميل صورة (صور). كرر هذه الخطوة لكل الأطوال الموجية وحفظ كدسات المكتملة.
- (اختياري) فتح جميع الصور المطلوبة للفيلم باستخدام الملف | فتح.
ملاحظة: من المستحسن لفتح الصور بطول موجي واحد المكتسبة في وقت لا تبطئ الكمبيوتر إذا كانت الطاقة الحسابية محدودة. إذا كان يجب أن يتم تخطي أجزاء معينة من تسجيلات الوقت الفاصل بين، مثلاً، البداية، النهاية، أو العديد من النقاط الزمنية، ثم هذا يمكن تعديلها في الفيلم المكتملة. - تنشيط كومة الصور التي تحتاج إلى تصويب للانجراف. افتح أداة المحاذاة ب تطبيقات | السيارات محاذاة.. الاختيار المكدس كمصدر للصور و الطائرة أول نقطة كمستوى مرجعي. حدد المكدس مع الزر مكدس المصدر ثم انقر فوق تطبيق.
ملاحظة: المحاذاة التلقائي سوف يستغرق بعض الوقت والطاقة الحسابية بل هو وسيلة جيدة لتصحيح مداخن كبيرة لانجراف تركيب المجهر. هذه المحاذاة التلقائية يعمل جيدا إذا كان يتم تضمين الجزئي المجالات الخرز الصغير لكن يمكن أيضا العمل بدونها. - حفظ المكدس تمت محاذاته.
- استخدام رويس.
ملاحظة: مجهرية الوقت الفاصل بين فلوري بسهولة إنشاء مجموعات كبيرة من ملفات البيانات التي يستغرق الكثير من الطاقة الحسابية وتبطئ التجهيز النهائي لهذه الأفلام. ولذلك نوصي بشدة بتحديد رويس وعزل الخلايا للعمل مع الملفات الأصغر حجماً.- حدد أداة منطقة مستطيلة . إنشاء دوروا حول الخلايا ذات الاهتمام بالرسم يدوياً عائد الاستثمار على الصورة التباين المرحلة. تأكد من أن الخلايا ذات الاهتمام مرئية وفي التركيز طوال الوقت الفاصل بين أسرة الفيلم.
- فتح الفيلم الوقت الفاصل بين من طول الموجه الثانية من نفس مجموعة البيانات. نقل العائد على الاستثمار من صور التباين المرحلة إلى الأسفار استخدام الصور من الطول الموجي الثاني أداة نقل المناطق مع المناطق | نقل المناطق. حدد مرحلة التباين dataset مصدر الصورة و dataset الطول الموجي الثاني صورة الوجهة. قم بتحديد جميع المناطق واضغط موافق.
- كرر الخطوة 4.6.2 لكل طول موجي المكتسبة لنفس مجموعة البيانات.
- تحديد العائد على الاستثمار، وتكرار ذلك مكدس مع تحرير | تكرار | مكدس الذاكرة المؤقتة... أو اضغط على مفاتيح Shift + Ctrl + D . ثم قم بحفظ بنية تخزين العناصر المكررة مع ملف | حفظ في نفس المجلد مثل البيانات الأصلية.
- كرر الخطوة 4.6.4 لكل عائد الاستثمار لكل طول موجي المكتسبة لنفس مجموعة البيانات
- لإنشاء أحد الأفلام في صيغ MOV أو AVI، فتح الدالة جعل الفيلم عبر المكدس | جعل الفيلم. حدد الوقت الفاصل بين التسجيلات مع الزر مكدس المصدر . حدد تنسيق الإخراج، معدل الإطار، والعدد من الإطارات، وانقر فوق حفظ.
Representative Results
م. إكسانثوس هو بكتيريا تزايد بطيء يتحرك على السطوح الصلبة. لاختبار الإعداد التجريبية لدينا، نحن إجراء تجربة الوقت الفاصل مع متحركة DK1622 WT الخلايا. مرحلة التباين الصور تم الحصول عليها على فترات لمدة 5 دقائق ح 24 (الشكل 2أ، ب). الانحياز معظم الخلايا في مجموعات. كما هو متوقع، خلايا عرض حركية ونقل معظمها في مجموعات. كذلك لاحظنا أن الخلايا أحياناً عكس اتجاه الحركة. وتوحي هذه النتائج أن خلايا وزن تحت ظروف اختبار تتصرف بشكل طبيعي من حيث حركية الخلية. ومع ذلك، حتى عندما تسجل الخلايا كل 5 دقائق، من الصعب تحديد خلايا فردية. وعلاوة على ذلك، لأن الخلايا غير متحركة، العديد من الخلايا الهروب أو أدخل مجال الرؤية مما يجعل من الصعب تتبع الخلايا لفترات طويلة.
من أجل تتبع نفس م. إكسانثوس الخلايا لعدة جولات من دورة الخلية بخلية يعيش التصوير، يمكن حذف السلالات الفردية للجينات مجلة ، الذي لا غنى عنه لحركية25. وهذا ما يمنع الخلايا من التحرك خارج مجال الرؤية من خلال بروتوكول التصوير. يتم إنشاؤها في إطار عمليات الحذف كما هو موضح بشي وآخرون. 26
كما هو متوقع، في مرحلة التصوير خلية يعيش التباين مع خلايامجلا Δ غير متحركة (الشكل 3)، لم يتم عرض خلايا حركة نشطة. كنا قادرين على متابعة النمو وتقسيم الخلايا الفردية أثناء تشكيل ميكروكولوني. واستنادا إلى الوقت الفاصل بين التسجيلات التي تم الحصول عليها من الصور على فترات لمدة 5 دقائق ح 24، كان من الممكن تحديد مقدار الوقت إينتيرديفيسيون (الفترة الزمنية بين حدثين انقسام الخلية) مع القرار خلية مفردة. خلايا متحولةمجلة Δ كان وقت بين شعبة من 235 ± 50 دقيقة (n = خلايا 97). مع ما يقرب من 4 ح، يشبه الوقت إينتيرديفيسيون إلى مضاعفة الوقت يقاس في الثقافات تعليق للخلايا WT. وهذا يوفر أدلة على أن م. إكسانثوس الخلايا تنمو على النحو الأمثل في ظل هذه الظروف التجريبية.
للتحقيق في ما إذا كان تنسيقنا الخلايا ينمو بشكل طبيعي أثناء تعقب البروتينات المسماة يفب على مدى فترات طويلة، أجرينا fluorescence الوقت الفاصل بين التصوير مع م. إكسانثوس الخلايا التي تعبر عن بروتين يفب معلم. وتحقيقا لهذه الغاية، تابعنا يفب فرع كعلامة لأصل النسخ المتماثل (أوري). فرع كمكون من مكونات نظام بارابS في م. إكسانثوس ، وربط المواقع بارس الدانية إلى أوري؛ ولذلك، يمكن فصل الازدواجية والكروموسوم الأصل يتبع19،،من2021. مع صورة اقتناء (مرحلة التباين والأسفار، ms 200 شراء الوقت في قناة يفب) كل 20 دقيقة، نمت الخلايا وتقسيم وعرض النمو حتى في ح 24 (الشكل 4أ). في بداية التسجيلات، شكلت فرع يفب مجموعتين في مناطق سوببولار في معظم الخلايا (الشكل 4أ). قبل فترة وجيزة أو بعد انقسام الخلية، فرع-يفب سوببولار الكتلة في القطب الخلية القديمة المكررة. وظلت واحدة من المجموعتين في القطب الخلية القديمة بينما النسخة الثانية ذوبانها إلى القطب الخلية الجديدة، وصلت إلى موقفها سوببولار النهائي بعد ما يقرب من 40-60 دقيقة (الشكل 4أ، ب). هذه الملاحظات بالاتفاق مع البيانات السابقة التي تم إنشاؤها من تسجيلات الوقت الفاصل بين قصيرة باستخدام منصات أجار رقيقة19. نخلص إلى أن هذا التشكيل التجريبي يسمح مجهرية الوقت الفاصل بين الأسفار لتتبع كروموسوم العزل على مدى عدة دورات الخلية في النمو البطيء م. إكسانثوس الخلايا، دون نمو الخلايا الانزعاج أو إليه العزل الصبغي.
في تجربة مماثلة، سعينا إلى تتبع علامات لانقسام الخلية بالأسفار الوقت الفاصل بين الفحص المجهري. مشابهة لما يقرب من جميع البكتيريا الأخرى، م. إكسانثوس يتطلب فتسز، جتباسي مثل توبولين بكتيرية، لانقسام الخلية16،،من1718. فتسز أشكال بنية تشبه خاتم في ميدسيل، ما يسمى Z-الحلبة، التي تساعد على توظيف جميع البروتينات الأخرى اللازمة لانقسام الخلايا27،28. في م. إكسانثوس، هو حفز تشكيل Z-الطوق وموقعها في ميدسيل بثلاثة بومكسيز البروتينات16،17. وتشكل هذه البروتينات الثلاثة معقدة مرتبطة بالكروموسوم الذي ينقل عبر نوكليويد من الموقع لانقسام الخلايا في خلية "الأم" في وسط نوكليويد في الخليتين ابنه. منتصف نوكليويد تتزامن مع ميدسيل، قبل العزل الصبغي، وهنا المجندين المعقدة بومكسيز فتسز ويحفز تشكيل Z-خاتم.
هنا، نحن أولاً ثم خلايا غير متحركة معربا عن فتسز-التجارة والنقل. لأنه يظهر فتسز-التجارة والنقل عموما إشارة الأسفار أضعف من فرع-يفب، قمنا بزيادة وقت التعرض إضعاف إلى 1 s في قناة التجارة والنقل. كما هو متوقع، تراكم قوية للتجارة والنقل فتسز لوحظ فقط في ميدسيل وتملي هذا التعريب موضع انقسام الخلية انقباض (الشكل 5ألف). FtsZ-التجارة والنقل أساسا تشكيل كتلة في ميدسيل في خلية أطول. كما كان من الواضح أن هذه المجموعة زيادة في كثافة على مر الزمن. وبعد انقسام الخلية، لاحظنا أن إعادة المتراكمة في ميدسيل في ابنه اثنين من فتسز-بروتينات فلورية خضراء الخلايا حوالي 2 ح في وقت لاحق (الشكل 5ب). وهذا يتفق مع الاستنتاج أن حوالي 50% خلايا في عدد سكان عرض الترجمة فتسز في ميدسيل استناداً إلى تحليل الأداة بالرصاص16،17.
في تجربة ثانية، تابعنا الخلايامجلة Δ غير متحركة ح 24 التي تعبر عن بومكس مشري. كجزء من نظام بومكسيز، بومكس يساعد على توجيه تشكيل Z-خاتم وتحديد المواقع، مما حفز انقسام الخلايا في ميدسيل16. إشارة بومكس مشري fluorescence قوية وتسمح بوقت تعرض في قناة الأسفار من السيدة 250 الأهم من ذلك، نما في حجم كافة الخلايا وعرض حدث انقسام الخلية على مدى التجربة، تشكيل ميكروكولونيس بعد 24 ساعة ( الشكل 6A). كما ذكر سابقا16، تقريبا كافة الخلايا الواردة كتلة بومكس مشري. أن غالبية هذه المترجمة في ميدسيل ومجموعات بعيداً عن ميدسيل ذوبانها إلى ميدسيل خلال فترة التجربة. خلال انقسامات الخلية، تم تقسيم المجموعات بومكس مشري، مع كل خلية ابنه تلقي كتلة. بدلاً من فتسز-التجارة والنقل، بومكس مشري المترجمة في ميدسيل 80-90% من دورة الخلية والتوصل إلى هذا الموقف قريبا بعد انقسام الخلية (الشكل 6ب).
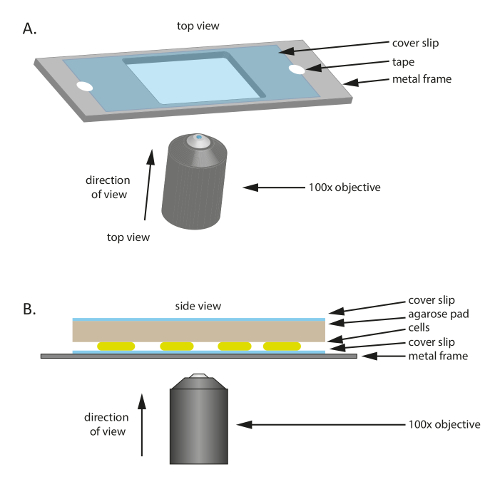
الشكل 1 : التخطيطي للإعداد التجريبية المستخدمة في هذه الدراسة. (أ) بالمعدن أو البلاستيك الإطار بمثابة دعم للعينة. هو ثابت ساترة للإطار المعدني مع الشريط للحد من الحركة للعينة. (ب) الجانب عرض إعداد نموذج تجريبي. هي الخلايا التي شنت على ساترة المبينة في (أ). يتم وضع لوح [اغروس] أن إمدادات المواد الغذائية والرطوبة إلى الخلايا أعلى الخلايا. اللوحة [اغروس] مشمولة ساترة إضافية لتقليل التبخر. لصور عالية الجودة، يتم استخدام X 100 زيت غمر مرحلة تباين هدف. الرجاء انقر هنا لمشاهدة نسخة أكبر من هذا الرقم-

الشكل 2 : مرحلة التباين الوقت الفاصل بين الفحص المجهري لوزن م. إكسانثوس الخلايا. خلايا تم اتباعها ح 24 والصور تم الحصول عليها كل دقيقة 5 (أ) تظهر الصور التمثيلية لنفس الحقل من عرض كل 5 دقائق. الأسهم الملونة تشير إلى اتجاه حركة الخلايا الفردية. يصادف نفس لون الخلية نفسها على مر الزمن. الأرقام تشير إلى الوقت بالدقائق. شريط الحجم: تظهر 5 ميكرومتر. (ب) الصور لنفس الحقل للعرض بعد كل ساعة. ملاحظة يتم إظهار نفس الحقل للعرض ولكن نظراً لنقل الخلايا، الخلايا باستمرار دخول وترك مجال الرؤية. الأرقام تشير إلى الوقت بالساعات. شريط الحجم: 5 ميكرومتر. درجة الحموضة: المرحلة التباين. الرجاء انقر هنا لمشاهدة نسخة أكبر من هذا الرقم-

الشكل 3 : مرحلة التباين الوقت الفاصل بين الفحص المجهري لغير متحركة م. إكسانثوس الخلايا. وتلت الخلايامجلا Δ 24 h. الصور تم الحصول عليها كل 5 دقائق وتظهر الصور التمثيلية بعد كل ساعة. يتم وضع علامة تضيقات انقسام الخلية المحددة مع الأسهم البرتقالية. الأرقام تشير إلى الوقت بالساعات. الرقم الهيدروجيني: مرحلة التباين. الرجاء انقر هنا لمشاهدة نسخة أكبر من هذا الرقم-
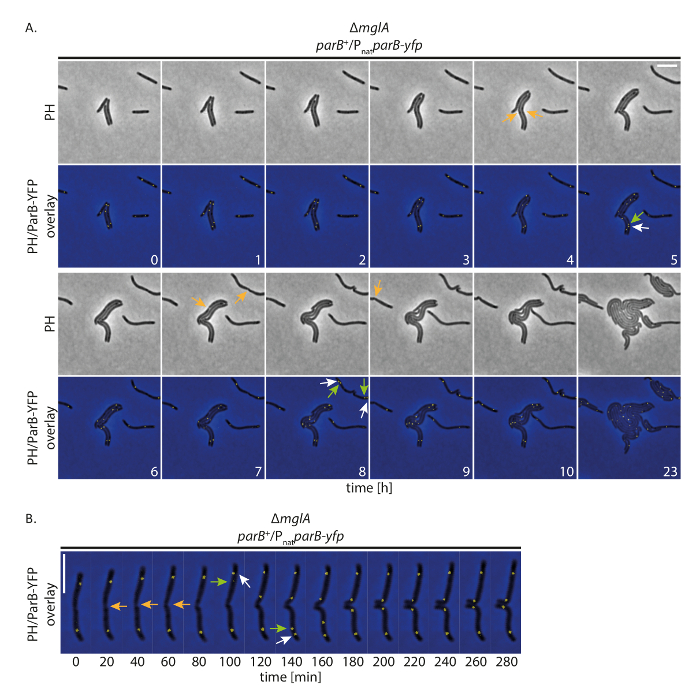
الشكل 4 : الوقت الفاصل بين الفحص المجهري fluorescence من فرع يفب في غير متحركة م. إكسانثوس الخلايا. خلايا من Δمجلة متحولةيفب فرع عن حضور الأم فرع (SA4749؛ Δمجلة؛ فرع +/Pناتفرع-يفب) تليها مرحلة الفحص المجهري التباين والأسفار عن ح 24- (أ) الصور تم الحصول عليها كل 20 دقيقة والممثل الصور كل ساعة حتى تظهر ح 10، جنبا إلى جنب مع نفس الخلايا بعد 24 حاء الصور تظهر في مرحلة التباين (PH) وكما تراكب من مرحلة التباين ويفب الإشارات. يتم وضع علامة انقسامات الخلية المحددة مع الأسهم البرتقالية. تشير الأسهم الأبيض والأخضر إلى أحداث ازدواجية يفب فرع الكتلة، مع الأسهم الخضراء وسم الكتلة ترانسلوكاتينج. الأرقام تشير إلى الوقت بالساعات. شريط الحجم: 5 ميكرومتر. (ب) الصور تم الحصول عليها كما هو الحال في (A) ولكن تظهر في أعلى الأزمنة. الأرقام تشير إلى الوقت بالدقائق. الأسهم كما هو الحال في (A). شريط الحجم: 5 ميكرومتر. الرجاء انقر هنا لمشاهدة نسخة أكبر من هذا الرقم-

الشكل 5 : الوقت الفاصل بين الفحص المجهري fluorescence من فتسز-التجارة والنقل في غير متحركة م. إكسانثوس الخلايا. خلايا متحولةمجلة Δ الإعراب عن فتسز-التجارة والنقل بحضور الأم فتسز (SA8241؛ Δمجلة؛ فتسز +/PناتftsZ-التجارة والنقل) تليها مرحلة الفحص المجهري التباين والأسفار عن ح 24. (أ) الصور تم الحصول عليها كل 20 دقيقة وتظهر الصور التمثيلية كل ساعة حتى ح 10، جنبا إلى جنب مع نفس الخلايا بعد 24 حاء الصور تظهر في مرحلة التباين (PH) وكما تراكب الإشارات بروتينات فلورية خضراء والمرحلة على النقيض من. يتم وضع علامة انقسامات الخلية المحددة مع الأسهم البرتقالية. تشير الأسهم البيضاء إلى مجموعات فتسز-التجارة والنقل في ميدسيل. الأرقام تشير إلى الوقت بالساعات. شريط الحجم: 5 ميكرومتر. (ب) الصور تم الحصول عليها كما هو الحال في (A) ولكن تظهر في أعلى الأزمنة. الأرقام تشير إلى الوقت بالدقائق. الأسهم الخضراء والبيضاء علامة الكتل FtsZ-التجارة والنقل في الخلايا اليسار واليمين، على التوالي. تشير الأسهم البرتقالية إلى انقسامات الخلية. شريط الحجم: 5 ميكرومتر. الرجاء انقر هنا لمشاهدة نسخة أكبر من هذا الرقم-

الرقم 6 : Fluorescence مجهرية الوقت الفاصل بين من مشري-بومكس في غير متحركة م. إكسانثوس الخلايا. Δ عدم متحركةبومكس الخلايا تتراكم بومكس مشري (SA4797؛ Δمجلة؛ Δبومكس/PpomZ مشري-بومكس) كانت تليها مرحلة التباين والأسفار مجهرية كل 20 دقيقة (A) ح 24 الممثل الصور تظهر كل ساعة حتى ح 10، جنبا إلى جنب مع نفس الخلايا بعد 24 حاء الصور تظهر في مرحلة التباين (PH) وكما تراكب مشري والمرحلة على النقيض من الإشارات. يتم وضع علامة انقسامات الخلية المحددة مع الأسهم البرتقالية. تشير الأسهم البيضاء والخضراء إلى مجموعات مشري-بومكس قبل وبعد تقسيم الأحداث، على التوالي. الأرقام تشير إلى الوقت بالساعات. شريط الحجم: 5 ميكرومتر. (ب) الصور تم الحصول عليها كما هو الحال في (A)، وتظهر في أعلى الأزمنة. الأسهم كما هو الحال في (A). شريط الحجم: 5 ميكرومتر. الرجاء انقر هنا لمشاهدة نسخة أكبر من هذا الرقم-
| سلالة بكتيرية | النمط الوراثي ذات الصلة1 | مرجع |
| DK1622 | Wildtype | 23 |
| SA4420 | Δمجلا | 24 |
| SA4749 | Δمجلة؛ وفرع+/أتتب:: فناتيفب فرع (pAH7) | هذه الدراسة |
| SA4797 | Δمجلة؛ Δبومكس/أتتب::فpomZ متشيري-بومكس (pAH53) | 16 |
| SA8241 | Δمجلة؛ فتسز+/mxan18-19::فناتftsZ-التجارة والنقل (pDS150) | هذه الدراسة |
| والبلازميدات بين أقواس معقوفة تحتوي على اندماج الجين المشار إليها وكانت intergated في المواقع المشار إليها على الجينوم. والبلازميدات متكاملة في موقع أتتب أو أعرب عن منطقة إينتيرجينيك mxan18-19 من المروج الأصلي (فnat) أو المروج الأصلي من pomZ (فpomZ). |
||
الجدول 1: قائمة بالسلالات البكتيرية المستخدمة في هذه الدراسة.
| البلازميدات | الخصائص ذات الصلة | مرجع |
| pAH7 | فناتفرع يفب؛ أتتب Mx8؛ TetR | 19 |
| pAH53 | PomZ ف مشري-بومكس؛ أتتب Mx8 ؛ كمصاد | 16 |
| pDS150 1 | فناتftsZ-التجارة والنقل ؛ mxan18-19 ؛ TetR | هذه الدراسة |
| pMR3691 | بلازميد للتعبير الجيني إيندوسيبلي فانيلاتي | 18 |
| pKA51 | فناتftsZ-التجارة والنقل ؛ أتتب Mx8؛ TetR | 17 |
| 1 pDS150: pDS150 هو مشتق pKA51 التي استعيض عن الموقع أتتب Mx8 بمنطقة إينتيرجينيك mxan18-19 . لهذا اتسع نطاق المنطقة إينتيرجينيك mxan18-19 من pMR3691 مع الإشعال Mxan18-19 الموجه إليه بسدري (جكجاتكاتجكجكجككاجاكجاتاكاجك) والقس Mxan18-19 بلبي (جكجكتجاجكككجكجككجاكاككجكاكك) والمستنسخة في pKA51. |
||
الجدول 2: قائمة والبلازميدات المستخدمة في هذه الدراسة.
Discussion
تصوير خلية يعيش الأسفار أصبح أداة قوية لدراسة ديناميات الزمانية المكانية للخلايا البكتيرية. مجهرية fluorescence الوقت الفاصل بين البكتيريا المتزايد متحركة وبطيئة مثل م. إكسانثوس، ومع ذلك، كانت صعبة وأنجز إلا لفترات زمنية قصيرة. هنا، نحن نقدم وسيلة سهلة الاستخدام وقوية لتصوير خلية يعيش م. إكسانثوس بالأسفار الوقت الفاصل بين الفحص المجهري. هذا الأسلوب يسمح للمستخدم بتتبع الخلايا والبروتينات المسمى فلوريسسينتلي لعدة جولات من دورة الخلية مع القرار خلية مفردة.
وهناك العديد من المتطلبات الأساسية التي تؤثر على نجاح التصوير خلية يعيش من النمو البطيء م. إكسانثوس الخلايا بما في ذلك: 1) سطح صلب لمرفق الخلية؛ 2) توافر المغذيات والأكسجين؛ 3) ثابتة من الرطوبة ودرجات الحرارة؛ و 4) الاستغلال الأمثل للظروف التجريبية مثل تواتر التعرض لاكتساب الوقت والصورة.
في لدينا الإعداد التجريبية، ونحن استخدام منصات سميكة [اغروس] استكمال مع المواد الغذائية. باستخدام منصات سميكة [اغروس] بدلاً من أجهزة موائع جزيئية لتتبع الخلايا المفردة لديه بعض الفوائد الأساسية ولكن أيضا بعض العيوب. أولاً، يوفر لوح [اغروس] ليس فقط سطح ل م. إكسانثوس خلية المرفقات وحركة ولكن أيضا المواد المغذية الكافية للنمو لمالا يقل عن 24 ساعة. وثانيا، المفاجئة التحليلات الرصاص تستخدم عادة لدراسة التعريب داخل الخلايا من البروتينات المسماة فلوريسسينتلي تم مسبقاً على نفس النوع من [اغروس] منصات16،،من1729. ولذلك، يمكن مقارنة البيانات من التحليلات الإضافية النار مباشرة إلى البيانات التي تم الحصول عليها بالطريقة الموضحة هنا. وثالثاً، يمكن تعديلها واستكمالها مع المضادات الحيوية أو المكملات الأخرى مثل CuSO4 منصات [اغروس] بسهولة وفانيلاتي التي تستخدم عادة للجينات التعبير التوجيهي18،30. أخيرا، لأن الخلايا مسموح بها بشكل ميكروكولونيس خلال فترة التجربة، هذا التشكيل يسمح أيضا دراسة تأثير التفاعلات الخلية الخلية مباشرة على معلمة معينة يجري تحليلها. هذا الجانب مهم بشكل خاص في حالة من م. إكسانثوس لأن هذه البكتيريا يعرض عدة الاتصال تعتمد على التفاعلات. ويعتبر العيب الرئيسي لهذا الأسلوب أن يتم مسبقاً الظروف التجريبية لمدة تجربة. وفي المقابل، تسمح أجهزة موائع جزيئية عموما تغيير الظروف التجريبية خلال فترة التجربة عن طريق إضافة المضادات الحيوية31على سبيل المثال.
تتوفر حزم البرمجيات الحرة (مثلاً، ميكروبيج، أووفتي) لتحليل نمو الخلايا المفردة والتعريب البروتين داخل الخلايا الفردية تلقائياً. ومع ذلك، هذه البرامج فقط مناسبة تماما لتحليل خلايا مفردة أو مجموعات صغيرة من الخلايا. وهكذا، فإنه لا يزال يشكل تحديا لتحليل البيانات التي تم إنشاؤها للتسجيلات ح 24 الموصوفة هنا تلقائياً.
وباختصار، وصفناها بروتوكولا سهلة الاستخدام واستنساخه لإجراء تصوير خلية يعيش مع تزايد بطيء م. إكسانثوس البكتيريا. ونحن تبين أن منصات بسيطة تكمل المغذيات [اغروس] كافية لدعم النمو لمالا يقل عن 24 ساعة، وتسمح بمراقبة وتحليل البروتين التعريب والنمو مع القرار خلية واحدة على مدى عدة أجيال.
Disclosures
الكتاب يعلن أن لديهم لا تضارب المصالح المالية.
Acknowledgments
وأيد هذا العمل بمجلس البحوث الألمانية (DFG) في إطار ترانسريجيو 174 "الزمانية المكانية ديناميات الخلايا البكتيرية" و "جمعية ماكس بلانك".
Materials
| Name | Company | Catalog Number | Comments |
| DMI6000B with AFC | Leica microsystems | 11888945 | Automated inverted widefield fluorescence microscope with adaptive focus control |
| Universal mounting frame | Leica microsystems | 11532338 | Stage holder for different sample sizes |
| HCX PL FLUOTAR 100x/1.30 oil PH3 | Leica microsystems | 11506197 | Phase contrast objective |
| Orca Flash 4.0 camera | Hamamatsu | 11532952 | 4.0 megapixel sCMOS camera for picture aquisition |
| Filter set TXR ET, k | Leica microsystems | 11504170 | Fluorescence filter set, Ex: 560/40 Em: 645/75 |
| Filter set L5 ET, k | Leica microsystems | 11504166 | Fluorescence filter set, Ex: 480/40 Em: 527/30 |
| Filter set YFP ET, k | Leica microsystems | 11504165 | Fluorescence filter set, Ex: 500/20 Em: 535/30 |
| ProScan III | Prior | H117N1, V31XYZEF, PS3J100 | Microscope automation controller with interactive control center |
| EL 6000 light source | Leica microsystems | 11504115 | External fluorescence light source |
| Incubator BLX Black | Pecon | 11532830 | Black incubation chamber surrounding the microscope |
| Tempcontrol 37-2 digital | Leica microsystems | 11521719 | Automated temperature control for incubation chamber |
| Gentmycin sulphate | Carl Roth | 0233.4 | Gentamycin |
| Oxytetracylin dihydrate | Sigma Aldrich | 201-212-8 | Oxytetracyclin |
| Kanamycin sulphate | Carl Roth | T832.3 | Kanamycin |
| Filtropur BT25 0.2 bottle top filter | Sarstedt | 831,822,101 | Bottle top filter for sterilization of buffers |
| Deckgläser | VWR | 630-1592 | Glass cover slip (60 x 22 mm, thickness: 0.7 mm) |
| Seakem LE agarose | Lonza | 50004 | Agarose for microscopy slides |
| Leica Metamorph AF | Leica microsystems | 11640901 | Microscope control software and software for picture analysis |
| Tetraspeck Microsperes, 0.5 µm | ThermoFisher | T7281 | Fluorescent microspheres |
| petri dish | Greiner Bio-one | 688102 | 120 mm x 120 mm x 17 mm squared petri dish for agarose pads |
| BD Bacto Casitone | Becton Dickinson | 225930 | Casitone |
| Parafilm M | VWR | 291-1213 | Parafilm |
| Tris(hydroxymethyl)-aminomethane | Carl Roth | AE15.2 | Tris |
| Magnesium sulphate heptahydrate | Carl Roth | P027.2 | Magnesium sulphate |
| Potassium dihydrogen phosphate p.a. | Carl Roth | 3904.1 | Potassium dihydrogen phosphate |
| 1% CTT medium: 1 % (w/v) BD Bacto™ casitone, 10 mM Tris-HCl ph 8.0, 1 mM potassium phosphate buffer pH 7.6, 8 mM MgSO4 | Cultivation medium for M.xanthus | ||
| TPM buffer: 10 mM Tris-HCl ph 8.0, 1 mM potassium phosphate buffer pH 7.6, 8 mM MgSO4 | Buffer for preparation of microscopy slides for M.xanthus |
References
- Shapiro, L., McAdams, H. H., Losick, R. Why and how bacteria localize proteins. Science. 326 (5957), 1225-1228 (2009).
- Treuner-Lange, A., Søgaard-Andersen, L. Regulation of cell polarity in bacteria. J Cell Biol. 206 (1), 7-17 (2014).
- Laloux, G., Jacobs-Wagner, C. Spatiotemporal control of PopZ localization through cell cycle-coupled multimerization. J Cell Biol. 201, 827-841 (2013).
- Rudner, D. Z., Losick, R.
- Badrinarayanan, A., Le, T. B. K., Laub, M. T. Bacterial chromosome organization and segregation. Annu Rev Cell Dev Biol. 31 (1), 171-199 (2015).
- Munoz-Dorado, J., Marcos-Torres, F. J., Garcia-Bravo, E., Moraleda-Munoz, A., Perez, J. Myxobacteria: Moving, Killing, Feeding, and Surviving Together. Front Microbiol. 7, 781 (2016).
- Berleman, J. E., Kirby, J. R. Deciphering the hunting strategy of a bacterial wolfpack. FEMS Microbiol Rev. 33 (5), 942-957 (2009).
- Konovalova, A., Petters, T., Søgaard-Andersen, L.
- Nudleman, E., Wall, D., Kaiser, D. Cell-to-cell transfer of bacterial outer membrane lipoproteins. Science. 309, 125-127 (2005).
- Vassallo, C. N., et al. Infectious polymorphic toxins delivered by outer membrane exchange discriminate kin in myxobacteria. eLife. 6, 29397 (2017).
- Vassallo, C., et al. Cell rejuvenation and social behaviors promoted by LPS exchange in myxobacteria. Proc Natl Acad Sci USA. 112 (22), 2939-2946 (2015).
- Li, Y., et al. Extracellular polysaccharides mediate pilus retraction during social motility of Myxococcus xanthus. Proc. Natl. Acad. Sci. USA. 100, 5443-5448 (2003).
- Kim, S. K., Kaiser, D. Cell alignment required in differentiation of Myxococcus xanthus. Science. 249, 926-928 (1990).
- Lobedanz, S., Søgaard-Andersen, L. Identification of the C-signal, a contact dependent morphogen coordinating multiple developmental responses in Myxococcus xanthus. Genes Dev. 17, 2151-2161 (2003).
- Schumacher, D., Søgaard-Andersen, L. Regulation of cell polarity in motility and cell division in Myxococcus xanthus. Annu Rev Microbiol. 71 (1), 61-78 (2017).
- Schumacher, D., et al. The PomXYZ proteins self-organize on the bacterial nucleoid to stimulate cell division. Dev Cell. 41 (3), 299-314 (2017).
- Treuner-Lange, A., et al. PomZ, a ParA-like protein, regulates Z-ring formation and cell division in Myxococcus xanthus. Mol Microbiol. 87 (2), 235-253 (2013).
- Iniesta, A. A., Garcia-Heras, F., Abellon-Ruiz, J., Gallego-Garcia, A., Elias-Arnanz, M. Two systems for conditional gene expression in Myxococcus xanthus inducible by isopropyl-beta-D-thiogalactopyranoside or vanillate. J Bacteriol. 194 (21), 5875-5885 (2012).
- Harms, A., Treuner-Lange, A., Schumacher, D., Søgaard-Andersen, L. Tracking of chromosome and replisome dynamics in Myxococcus xanthus. reveals a novel chromosome arrangement. PLoS Genet. 9 (9), 1003802 (2013).
- Iniesta, A. A. ParABS system in chromosome partitioning in the bacterium Myxococcus xanthus. PLoS One. 9 (1), 86897 (2014).
- Lin, L., Osorio Valeriano, M., Harms, A., Søgaard-Andersen, L., Thanbichler, M. Bactofilin-mediated organization of the ParABS chromosome segregation system in Myxococcus xanthus. Nat Commun. 8 (1), 1817 (2017).
- Hodgkin, J., Kaiser, D. Cell-to-cell stimulation of movement in nonmotile mutants of Myxococcus. Proc Natl Acad Sci U S A. 74 (7), 2938-2942 (1977).
- Kaiser, D. Social gliding is correlated with the presence of pili in Myxococcus xanthus. Proc Natl Acad Sci USA. 76 (11), 5952-5956 (1979).
- Miertzschke, M., et al. Structural analysis of the Ras-like G protein MglA and its cognate GAP MglB and implications for bacterial polarity. EMBO J. 30 (20), 4185-4197 (2011).
- Hodgkin, J., Kaiser, D. Genetics of gliding motility in Myxococcus xanthus. (Myxobacterales): Two gene systems control movement. Mol Gen Genet. 171, 177-191 (1979).
- Shi, X., et al. Bioinformatics and experimental analysis of proteins of two-component systems in Myxococcus xanthus. J Bacteriol. 190 (2), 613-624 (2008).
- Bi, E. F., Lutkenhaus, J. FtsZ ring structure associated with division in Escherichia coli. Nature. 354 (6349), 161-164 (1991).
- Lutkenhaus, J., Pichoff, S., Du, S. Bacterial cytokinesis: From Z ring to divisome. Cytoskeleton. 69 (10), 778-790 (2012).
- McLoon, A. L., et al. MglC, a Paralog of Myxococcus xanthus GTPase-Activating Protein MglB, Plays a Divergent Role in Motility Regulation. J Bacteriol. 198 (3), 510-520 (2015).
- Gomez-Santos, N., et al. Comprehensive set of integrative plasmid vectors for copper-inducible gene expression in Myxococcus xanthus. Appl Environ Microbiol. 78 (8), 2515-2521 (2012).
- Treuner-Lange, A., et al. The small G-protein MglA connects to the MreB actin cytoskeleton at bacterial focal adhesions. J Cell Biol. 210 (2), 243-256 (2015).
