

Summary
细菌细胞的空间高度组织。为了跟随这个组织随着时间的推移在缓慢生长的Myxococcus xanthus细胞, 建立了荧光活细胞成像的一个高时空分辨率的几个世代。利用该方法可以确定重要蛋白质在染色体分离和细胞分裂中的时空动力学。
Abstract
细菌细胞的荧光活细胞成像是分析中心细胞周期事件的蛋白质和染色体的时空动态的关键方法。然而, 在缓慢生长的细菌中, 这些分子的成像是一个挑战, 由于漂白的显影和毒性在图像采集过程中。在这里, 我们描述了一个简单的协议, 以规避这些限制的情况下, Myxococcus xanthus (它的世代时间 4-6 小时)。为此, m. xanthus细胞生长在一个富含营养的琼脂垫上, 温度控制湿润环境。在这些条件下, 我们通过跟踪单个细胞的生长来确定单个细胞的加倍时间。此外, 关键的细胞过程, 如染色体分离和细胞分裂可以通过荧光活细胞成像的细胞, 包括相关荧光标记蛋白, 如 ParB YFP, FtsZ GFP, mCherry PomX 超过多个细胞周期。随后, 获取的图像被处理生成蒙太奇和/或电影。
Introduction
细菌细胞在空间上高度组织与许多蛋白质本地化非对称在细胞舱1,2,3,4。这种局部化通常是高度动态的, 随着时间的推移, 对细胞周期提示或外部信号发生变化。同样地, 细菌染色体在空间上高度组织与各自的基因座被安置到特定亚细胞位置在分离过程之前和期间5。这种动态空间组织对于生长、分裂、细胞周期调控、分化、运动、信号传导以及染色体组织和分离等都具有重要意义;因此, 它主要影响细菌功能的所有方面。
这些细胞过程的时空动力学正在分析各种不同的细菌物种, 大肠杆菌,枯草芽孢杆菌, 霍乱弧菌, Caulobacter crescentus作为重要模型有机体。然而, 这四种物种只涵盖了巨大的细菌多样性的小范围, 也许不奇怪的是, 由于这些物种之间的大系统发育距离, 细胞组织和极化机制在这些不同的细菌。这就提高了研究其他细菌种类的必要性, 以便最终提取细菌细胞时空动力学的一般原理。
革兰阴性三角洲-proteobacterium m. xanthus是研究细菌6中的社会行为和合作的模型有机体。m. xanthus是一个严格的好氧生物, 在营养物质的存在下, 它形成了细胞以高度协调、蜂拥的方式向外扩散, 并捕食其他微生物7的菌落。为了应对营养饥饿, 细胞启动了一个发育计划, 结果形成了由数以千计的细胞组成的果体, 在其中, 杆状运动细胞分化为球状双胞孢子8。这两种类型的行为,即蜂拥和果体形成, 只有被放置在固体表面的细胞执行。此外, 在这两种营养条件下, 细胞都参与了涉及直接细胞接触的过程, 包括交换外部膜脂蛋白, 在接受者9、10 中可能刺激运动或作为毒素作用., 交换 LPS11, 刺激运动的多糖对相邻的细胞12, 细胞间信号传递的单元格表面定位信号蛋白13,14。
最近, m. xanthus也成为研究运动的机制和它的章程15, 细胞分裂16,17,18和染色体组织19的模型有机体 ,20,21。在m. xanthus细胞周期的关键步骤已详细分析了荧光显微镜下使用的快照图像或短时间推移记录的菌株携带相关的荧光标记蛋白16, 17,18,19,20。理想情况下, 许多细胞应遵循单细胞分辨率的荧光活细胞成像至少一个完整的细胞周期, 以获得可靠的定量数据的细胞周期参数。然而, 这是一个挑战, 在 m. xanthus的情况下, 由于其相对长的时间 4-6 小时在标准实验室条件下, 由于漂白显影和毒性在图像采集。
在这里, 我们描述了一个协议, 遵循m xanthus细胞与单细胞分辨率的荧光活细胞成像至少24小时, 并涵盖几个细胞周期。重要的是, 在整个协议期间, 细胞保持在琼脂垫上, 并密切接触, 允许接触依赖的活动至关重要的社会生活方式的 m. xanthus。该协议还允许用户在高时间分辨率和单细胞分辨率下监视形状、大小、除法和荧光探针, 从而能够量化细胞间的变异性和细胞周期事件的相关性。
Protocol
1. xanthus菌株的制备和生长
注: 见表 1和表 2。
- 准备 1% casitone 肉汤 (铁通) 生长培养基 1% (w/v) 胰酪蛋白 (如Bacto casitone), 10 毫米三盐酸 ph 值 8.0, 1 毫米 2PO 4 ph 7.6, 8 毫米 MgSO422, 补充卡那霉素 (50 µg/毫升) 或土霉素 (10 µg/毫升)。添加庆大霉素 (10 µg/毫升), 以减少与其他细菌污染的风险, 因为m. xanthus细胞自然抗它。
- 接种5毫升1% 铁通含有相关抗生素与一个新鲜的野生类型的殖民地 (DK1622) 23, SA4420 (δmglA)24, SA4797 (δmglA, ΔpomX/PpomZ mCherry-pomX)16, SA8241 (δmglA, ftsZ+/pnatftsZ-gfp), 或 SA4749 (δmglA, parB+/pnatparB-yfp) 在1天的早晨。
- 并用重悬一个xanthus殖民地在500µL 1% 铁通补充抗生素在一个无菌管和转移整个悬浮液50毫升锥形瓶含有5毫升1% 铁通。
注: 使用锥形瓶的10倍的数量的文化, 以保证足够的 aeriation 和最佳增长。
- 并用重悬一个xanthus殖民地在500µL 1% 铁通补充抗生素在一个无菌管和转移整个悬浮液50毫升锥形瓶含有5毫升1% 铁通。
- 生长细胞为八代 (大约 40-48 h 以世代时间 4-6 h) 在32°c, 震动在 220 rpm, 在黑暗中。维持指数生长阶段的细胞 (OD550 < 1.2), 防止它们到达固定阶段。如果需要, 将细胞稀释成新鲜的1% 铁通培养基, 其中含有相关的抗生素到550的外径 0.1-0.2。
注: 单个细胞显微镜的最佳 OD550为 0.5-0.7。在这个 OD550中, 每个图像都有足够数量的单元格, 以便对细胞参数进行量化和统计分析。
2. 显微镜样品的制备
注意: 通过显微镜观察的细胞被放置在显微镜盖玻片上, 然后由含有营养的琼脂糖垫覆盖。盖玻片粘在塑料或金属框架上, 以提供机械支撑。为准备显微术, 应提前准备1% 个大垫子, 如步骤 2.1-2.3 所述。请参阅本表中所使用的特定产品的材料。
- 准备500毫升的 TPM 缓冲器 (10 毫米三盐酸 pH 7.6, 1 毫米的 MgSO2 毫升4 pH 7.6, 8 毫米4) 和高压釜或过滤器消毒使用瓶顶过滤器。
注: 无菌缓冲器可在室温下贮存数月。 - 准备1% 琼脂糖显微镜溶液, 含0.2% 铁通 (混合1克琼脂糖与80毫升的 TPM 缓冲器和20毫升的1% 铁通培养基)。在微波炉加热直到琼脂糖融化。
注: 0.2% 铁通足以使细胞生长和防止饥饿。在显微介质中较高浓度的铁通会产生高的背景荧光。 - 填充一个培养皿与熔融琼脂糖的厚度为0.5 厘米 (11.5 厘米 x 11.5 厘米方培养皿, 约60毫升的熔融琼脂糖), 让它降温到室温。
注: 琼脂糖垫可在潮湿环境下贮存4摄氏度, 长达2天。- 预热1% 琼脂糖/TPM/0.2% 铁通垫32°c 至少15分钟前使用。
注: 要准备细胞显微镜, 请按照步骤 2.4-2.8。
- 预热1% 琼脂糖/TPM/0.2% 铁通垫32°c 至少15分钟前使用。
- 将无菌玻璃盖玻片 (60 毫米 x 22 毫米, 厚度: 0.7 毫米) 放在中间有孔的塑料或金属框架上 (图 1a);这个框架作为一个机械支持的薄盖玻片, 有助于减少漂移在显微镜下。用磁带将盖玻片固定在框架上。
- 要准备框架, 切出一个75毫米 x 25 毫米框架从1毫米厚的金属板, 然后切出一个适当大小的孔 (在这个实验中20毫米 x 30 毫米) 在中间。
- 在盖玻片上添加 10-20 µL 指数生长的m xanthus细胞。
- 添加荧光0.5 µm 微球作为基准标记的细胞, 以简化跟踪的细胞或蛋白质的时间推移记录。
- 在 TPM 缓冲器中稀释微球 1:100, 贮存在4摄氏度, 长达数月。在使用前彻底摇动, 并将稀释微球的 5-10 µL 添加到细胞中。
注: 这里使用的是荧光在所有共同的蓝色, 绿色, 黄色和红色荧光通道的微球。
- 在 TPM 缓冲器中稀释微球 1:100, 贮存在4摄氏度, 长达数月。在使用前彻底摇动, 并将稀释微球的 5-10 µL 添加到细胞中。
- 切出一个小垫约大小的盖玻片的大预热1% 琼脂糖/TPM/0.2% 铁通垫, 并把它放在细胞顶部 (图 1B)。放置一个盖玻片在1% 琼脂糖/TPM/0.2% 铁通琼脂糖垫上, 以防止蒸发和维持在潮湿的环境中的细胞。
注: 盖玻片仅能防止2小时的显著蒸发。对于较长的延时记录, 1% 琼脂糖/TPM/0.2% 铁通垫和盖玻片三明治应密封与石蜡膜, 以防止蒸发。 - 孵育显微镜样品在32°c 15-20 分钟, 让细胞附着在琼脂糖垫底部。然后开始时间失效显微记录。
3. 显微镜设置和定时采集
注: 此处描述的协议是为倒置 widefield 显微镜开发的, 具有自动对焦、100 x/1.30 NA 油 PH3 目标、X、Y 机动化阶段、sCMOS 相机、光源、绿色荧光、红色荧光或黄色荧光滤镜蛋白质和温度控制孵化室。这个会议厅保持细胞免受光照和恒温。
- 预热的孵化室和显微镜到32°c 1-2 小时前开始显微镜。
注: 根据显微镜的设置, 加热可能需要更长的时间。预加热是降低漂移、稳定自动对焦控制系统的关键。 - 打开显微镜, 启动显微镜控制软件。选择正确的目标和正确的镜子和过滤器获取相衬图像, 以及绿色荧光, 红色荧光, 或黄色荧光蛋白的图像。
注: 显微镜通常提供一个用于显微镜控制和图像采集的首选软件。这里有一个商业上可用的软件 (见材料表) 用于控制显微镜和图像采集。 - 添加一滴高品质浸入油到物镜的目标和底部的样品预孵化32摄氏度。将目标置于尽可能低的 Z 位置, 以避免在将样品放置到显微镜阶段时损害物镜。将金属框架与样品放到显微镜台上, 并将其与 "孔边" 朝向目标。将样品牢固地固定在舞台支架上。
- 通过在 Z 方向上移近目标来聚焦细胞。当油滴在样品的底部和物镜接触时, 移动舞台会慢一些。将舞台移动到 X/Y 方向, 直到在视图区域中显示多个单个单元格时, 当单元格处于焦点平面时。确保在视图区域中至少有一个荧光微球, 以便以后对齐获得的图像。
注: 在最佳条件下, 应达到每个区域的 15-30 个细胞密度 (2048 x 2048 像素或 133.1 x 133.1 µm)。 - 打开显微镜控制软件的多维采集向导, 建立一个时间失效实验, 允许显微镜在需要时在多个波长和舞台位置获取图像。
- 在主选项卡激活Timelapse和多个波长。其他选项卡将显示在窗口的左侧。
- 单击 "保存" 选项卡并选择 "目录" 以选择计算机硬盘上的空文件夹以保存获取的图像。如果文件存在, 请激活增量基名称, 以确保连续的数据集不覆盖前面的 dataset。然后给实验一个名称的日期和应变名称或标题的实验。
- 单击 " Timelapse " 选项卡以调整时间推移参数。将工期设置为24小时, 并将时间间隔设置为20分钟。时间点的数量将自动更改。
注: 最佳时间间隔取决于实验和要分析的细胞功能。频繁的图像收购可能会导致漂白。因此, 必须从经验上发现, 在时间分辨率和漂白之间进行权衡。在 4-6 小时的加倍时间, 图像可以很容易地获得的间隔5分钟 (甚至更小的间隔, 如果需要) 的相位对比显微镜。如果在24小时的时间路线的荧光显微镜是期望的图像应该被记录在大约 15-30 分钟的间隔。 - 单击 "波长" 选项卡. 通过更改数字, 选择每个时间点为每个图像获取的波长数。
注意: 对于每个波长, 一个新的标签将出现在多维采集向导的左侧, 波长将从上到下依次获取。对于每个波长, 可以分别修改采集设置。 - 单击顶部的第一个波长选项卡。在 "光照" 下拉列表中选择 "相位对比度"。选择100毫秒以进行曝光, 并在 "获取" 下拉列表中选择每个时间点。在下拉列表中选择 "从不", 停用自动公开。
- 对每一个需要在每个时间点获取的波长重复步骤3.5.5。在这里描述的实验性的设置和荧光标记的蛋白质, 使用以下参数来暴露: 250 ms 为 mCherry 融合蛋白, 200 ms YFP 融合蛋白, 1000 ms 的 GFP 融合蛋白。
注意: 应通过改变每个波长的灯强度和图像采集时间, 预先确定每种应变和荧光蛋白的最佳光照设置。太长的图像采集时间会增加光毒性效果, 最终导致生长骤停和细胞死亡。因此, 应该实现图像质量与细胞生存能力之间的权衡。 - 从多个阶段的位置获取图像, 以增加在同一实验中记录的细胞数。
- 要从多个舞台位置获取图像, 请在主选项卡中激活多个舞台位置。然后单击 "舞台" 选项卡, 然后单击 "实时" 按钮以查看视图字段。
- 在 X/Y 方向上移动舞台, 直到感兴趣的区域 (ROI) 位于视图字段中。通过单击 "舞台" 选项卡中的 "+" 来保存 X 和 Y 坐标. 在 x/y 方向上再次移动舞台, 直到找到新的 ROI 并通过单击 "+" 再次保存坐标。继续, 直到保存所需的区域数。
注: 在荧光图像采集的情况下, 请确保感兴趣的区域 (ROIs) 不太靠近, 以最小化毒性。
- 通过单击不同的保存的 X 和 Y 位置, 再检查单元格的焦点, 然后单击AFC 保持以在实验过程中保持保存的 Z 位置常数, 然后启动硬件自动对焦。
- 通过单击多维采集向导中的 "获取", 在显微镜控制软件中启动定时记录。
注意: 每个波长都将显示一个窗口, 并且会显示一个额外的窗口, 显示获得的时间点数和时间直到下一次图片获取。 - 检查在时间推移录音中的前几个时间点之后, 单元格仍然处于焦点, 以最大化图像的质量, 并在需要时重新对焦。
4. 定时电影和图像对准的生成
注: 几种商用和免费软件包可用于图像采集和图像分析。我们使用一个商业上可用的软件 (见材料表) 与多个预安装的插件和其他工具。
- 在安装了图像分析/处理软件的计算机上, 将单个图像保存为定时录制。
- 通过单击 "查看多维数据" 来启动软件并以堆栈打开图像|选择基文件 |选择目录。使用多维数据打开文件夹。检查数据集并单击 "视图";数据集将显示为从时间点一到结尾的单个图像。激活波长 (用于创建堆栈), 选择堆栈中应该存在的所有图像, 然后单击 "加载图像"。对所有波长重复此步骤, 并保存已完成的堆栈。
- 选使用文件打开影片所需的所有图像|打开。
注: 建议在计算能力有限的情况下, 一次用一个获得的波长打开图像, 以不减慢计算机的速度。如果时间推移录制的某些部分 (例如, 开始、结束或多个时间点) 应跳过, 则可以在已完成的影片中对其进行调整。 - 激活需要纠正漂移的图像堆栈。通过应用程序打开对齐工具|自动对齐..................... 使用 "源堆栈" 按钮选择堆栈, 然后单击 "应用"。
注: 自动对准将需要一些时间和计算能力, 但是一个很好的方法来纠正大栈漂移的显微镜设置。这种自动对准工作很好, 如果微球包括在内, 但也可能在没有它们的工作。 - 保存对齐的堆栈。
- 使用 ROIs。
注意: 荧光时间推移显微镜可以轻松地创建大量的数据文件, 它们占了大量的计算能力, 减慢了这些影片的下游处理速度。因此, 我们强烈建议识别 ROIs 并隔离单元格以使用较小的文件。- 选择矩形区域工具。通过在相位对比度图像上手动绘制 roi, 在感兴趣的单元格周围创建 roi。确保感兴趣的单元格是可见的, 并在整个时间推移电影的焦点。
- 打开同一数据集的第二个波长的延时电影。要将 ROI 从相对比图片转移到第二波长的荧光图像, 请使用带区域的传输区域工具|转移区域。选择相位对比数据集作为源图像和第二波长数据集作为目标图像。选择所有区域并按"确定"。
- 对为同一数据集获取的每个波长重复步骤4.6.2。
- 选择 ROI 并将其作为堆栈复制到编辑 |复制 |堆栈...或按Shift + Ctrl + D键。然后将重复的堆栈保存为文件 |保存到与原始数据相同的文件夹中。
- 重复步骤4.6.4 为同一数据集获取的每个波长的 ROI
- 若要以 "动画" 或 AVI 格式生成影片, 请通过堆栈打开 "制作影片" 功能|拍电影。使用 "源堆栈" 按钮选择时间间隔录制。选择输出格式、帧速率、帧数, 然后单击 "保存"。
Representative Results
m. xanthus是一种生长缓慢的细菌, 在固体表面上移动。为了测试实验的建立, 我们对运动 DK1622 细胞进行了时间推移实验。相位对比图像获得的间隔为5分钟为24小时 (图 2A, B)。大多数单元格按组排列。正如预期的, 细胞显示运动和主要移动组。我们进一步观察到, 细胞有时会逆转运动方向。这些发现表明, 在受试条件下的细胞在细胞运动时表现正常。然而, 即使细胞每5分钟被记录, 每个细胞的鉴定是困难的。此外, 由于细胞是运动性的, 许多细胞逃离或进入视野, 使得在较长时期内难以跟踪细胞。
为了通过活细胞成像追踪同一m xanthus细胞的几轮细胞周期, 单个菌株可以被删除的mglA基因, 这是必不可少的运动25。这可防止在成像协议过程中单元格从视图区域移出。帧内删除是由石et的描述生成的。26
正如预期的, 在相对比活细胞成像与非活动ΔmglA细胞 (图 3), 细胞没有显示积极的运动。在 microcolony 形成过程中, 我们能够遵循个体细胞的生长和分裂。根据在5分钟内以24小时为间隔获取图像的时间推移记录, 可以将 interdivision 时间 (两个细胞分裂事件之间的时间) 与单细胞分辨率进行量化。ΔmglA突变体细胞的分裂时间为 235 50 min (n = 97 细胞)。与大约4小时, interdivision 时间是相似的加倍的时间在悬浮培养被测量为重量细胞。这提供了证据, m. xanthus细胞在这些实验条件下生长最佳。
为了调查我们的设置是否允许细胞在长时间跟踪 YFP 标记的蛋白质时正常生长, 我们用m xanthus细胞进行荧光时移成像, 表达 YFP 标记蛋白。为此, 我们遵循 ParB YFP 作为复制起源 (ori) 的标志。ParB 是 ParABS系统的组分在m xanthus并且结合到接近ori的近端位置;因此, 起源复制和染色体分离可以遵循19,20,21。与图像采集 (相对比和荧光, 200 ms 采集时间在 YFP 通道) 每20分钟, 细胞增长, 分裂, 并显示增长, 即使在 24 h (图 4A)。在录音开始时, ParB-YFP 在大多数单元格的 subpolar 区域形成了两个簇 (图 4A)。在细胞分裂之前或之后不久, subpolar ParB-YFP 簇在老细胞极点复制。两个星团中的一个保持在旧的细胞极点, 而第二个拷贝移位到新的细胞极点, 达到其最终 subpolar 位置后约 40-60 分钟 (图 4A, B)。这些观测与以前使用薄琼脂19的短时间失效记录所产生的数据一致。我们的结论是, 这种实验性的建立允许荧光的时间推移显微镜跟踪染色体分离的几个细胞周期的缓慢增长的m. xanthus细胞, 没有扰动细胞生长或染色体分离机械。
在类似的实验中, 我们试图追踪细胞分裂的标记, 时间失效荧光显微镜。与几乎所有其他细菌相似, m. xanthus需要 FtsZ, 一种细菌蛋白象 GTPase, 为细胞分裂16,17,18。FtsZ 形成一个环状结构在 midcell, 所谓的 Z 圆环, 帮助招募所有其他蛋白质需要为细胞分裂27,28。在m xanthus, Z 圆环的形成和它的定位在 midcell 由三 PomXYZ 蛋白质16,17刺激。这三种蛋白质形成染色体相关的复合体, 通过 nucleoid 从细胞分裂的地方转移到两个女儿细胞中的 nucleoid。在染色体分离之前, nucleoid 的中部与 midcell 重合, PomXYZ 复杂的新兵 FtsZ 并刺激 Z 环的形成。
在这里, 我们首先遵循的非运动细胞表达ftsZ gfp。由于 FtsZ-gfp 整体显示的荧光信号比 ParB YFP 弱, 我们在 GFP 通道中增加了5倍到1秒的曝光时间。正如预期的那样, FtsZ-GFP 的强积累只在 midcell 观察到, 这种定位决定了细胞分裂收缩的位置 (图 5A)。FtsZ-GFP 主要地形成了一个群在 midcell 在更长的细胞。同样明显的是, 随着时间的推移, 这个簇的强度增加了。细胞分裂后, 我们观察到, FtsZ-GFP 在 midcell 的两个女儿细胞中重新积累约2小时后 (图 5B)。这与发现大约50% 的人口中的细胞显示 FtsZ 本地化在 midcell 基于快照分析16,17是一致的。
在第二个实验中, 我们遵循非运动的ΔmglA细胞为 24 h 表达mCherry-pomX。作为 PomXYZ 系统的一部分, PomX 有助于指导 Z 环的形成和定位, 从而刺激细胞分裂在 midcell16。mCherry-PomX 的荧光信号强度强, 在250毫秒的荧光通道中允许曝光时间. 重要的是, 所有细胞在实验过程中都生长在大小, 并显示细胞分裂事件, 形成 microcolonies 后24小时 (图 6A)。正如以前报告的16, 几乎所有的细胞都含有 mCherry PomX 簇。大多数这些地方在 midcell 和集群远离 midcell 移位在实验过程中 midcell。在细胞分裂过程中, mCherry-PomX 簇分裂, 每个子细胞接收一个簇。与 FtsZ-GFP 相反, mCherry PomX 在细胞周期的 midcell 80-90% 上进行局部定位, 并在细胞分裂后不久到达这个位置 (图 6B)。
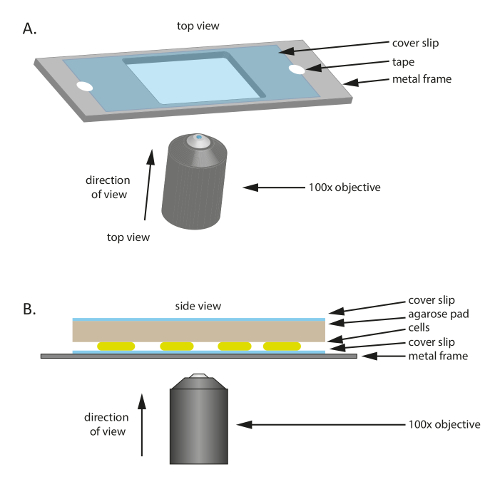
图 1: 在本研究中使用的实验设置示意图.(a) 金属或塑料框架用作样品的支持。盖玻片用胶带固定在金属框架上, 以减少试样的运动。(B) 实验样本设置的侧面视图。单元格安装在 (A) 中显示的盖玻片上。为细胞提供养分和湿度的琼脂糖垫放在细胞的顶端。琼脂糖垫被额外的盖玻片覆盖, 以减少蒸发。对于高质量的图像, 使用100X 油浸相对比目标。请单击此处查看此图的较大版本.

图 2: xanthus细胞的相位对比时间-失效显微术.细胞被跟随了24小时, 并且图片被获取每5分钟. (A) 显示同一视野的代表性图像每5分钟。彩色箭头表示单个细胞运动的方向性。同一颜色在一段时间内标记同一个单元格。数字表示时间 (分钟)。缩放栏: 5 µm (B) 显示每小时后相同视野的图像。请注意, 显示了相同的视图字段, 但由于单元格正在移动, 因此单元格不断地进入和离开视图字段。数字表示时间 (小时)。刻度条: 5 µm PH 值: 相衬。请单击此处查看此图的较大版本.

图 3: 非运动性m xanthus细胞的相对比时间-失效显微术.ΔmglA细胞被跟随了 24 h. 图像在每小时显示后每5分钟和代表图像获得。选定的单元格分区收缩用橙色箭头标记。数字表示时间 (小时)。PH 值: 相衬。请单击此处查看此图的较大版本.
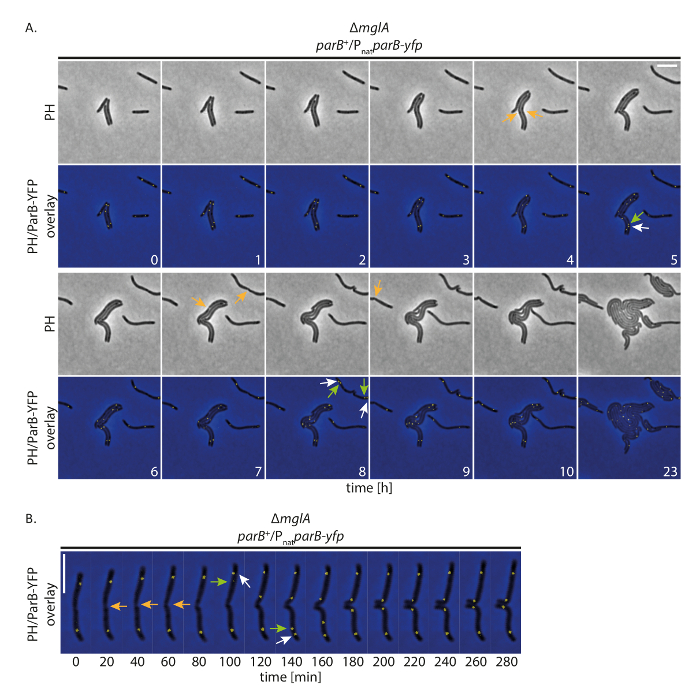
图 4: ParB-YFP 在非运动性m xanthus细胞中的荧光时间失效显微术。δmglA突变体的细胞,表达 parB yfp在本机parB的存在 (SA4749; ΔmglA;parBparByfp) 通过相对比和荧光显微术对24小时进行了跟踪. (A) 图像每小时获得20分钟和代表性图像, 直到 10 h 显示, 以及 24 h 之后的相同单元格. 图像显示在相对比 (PH) 和叠加的相位对比和 YFP 信号。选定的单元格分区用橙色箭头标记。白色和绿色箭头表示 ParB YFP 群集复制事件, 其中绿色箭头标记了转运群集。数字表示时间 (小时)。刻度条: 5 µm (B) 图像是在 (A) 中获得的, 但在更高的时间分辨率下显示。数字表示时间 (分钟)。箭和 (A) 一样。刻度条: 5 µm.请点击这里查看这个数字的大版本.

图 5: FtsZ-GFP 在非运动性m xanthus细胞中的荧光时间失效显微术.δmglA突变体的细胞, 表达ftsZ-gfp在本地ftsZ的存在 (SA8241; ΔmglA;ftsZ通过相位对比和荧光显微术,ftsZ了 24 h。(A) 图像每小时获得20分钟和代表性图像, 直到 10 h 显示, 以及 24 h 之后的相同单元格. 图像显示在相对比 (PH) 和叠加的相位对比和 GFP 信号。选定的单元格分区用橙色箭头标记。白色箭头表示 FtsZ-GFP 簇在 midcell。数字表示时间 (小时)。刻度条: 5 µm (B) 图像是在 (A) 中获得的, 但在更高的时间分辨率下显示。数字表示时间 (分钟)。绿色和白色箭头分别在左和右单元格中标记 FtsZ GFP 簇。橙色箭头表示细胞分裂。刻度条: 5 µm.请点击这里查看这个数字的大版本.

图 6: mCherry-PomX 在非运动性m xanthus细胞中的荧光时间失效显微术.非运动ΔpomX细胞积累 mCherry-pomX (SA4797; δmglA; δpomX/PpomZ mCherry) 通过相对比和荧光显微镜每20分钟跟踪24小时. (A) 代表图像每小时, 直到10小时显示, 连同相同的细胞后, 24 h. 图像显示在相对比 (PH) 和叠加的相位对比和 mCherry 信号。选定的单元格分区用橙色箭头标记。白色和绿色箭头分别表示在拆分事件前后 mCherry PomX 簇。数字表示时间 (小时)。刻度条: 5 µm (B) 图像是在 (A) 中获得的, 在更高的时间分辨率下显示。箭和 (A) 一样。刻度条: 5 µm.请点击这里查看这个数字的大版本.
| 菌株 | 相关基因型1 | 参考 |
| DK1622 | Wildtype | 23 |
| SA4420 | ΔmglA | 24 |
| SA4749 | ΔmglA; parB+/attB::PnatparB yfp (pAH7) | 本研究 |
| SA4797 | ΔmglA;ΔpomX/attB::PpomZ mCherry pomX (pAH53) | 16 |
| SA8241 | ΔmglA; ftsZ+/mxan18-19::PnatftsZ-gfp (pDS150) | 本研究 |
| 在括号内的质粒包含表明的基因融合和被 intergated 在指定的地点在基因组。 在attB或mxan18-19基因间地区集成的质粒从 他们的原生启动子 (pnat) 或 pomZ 的本机启动子 (ppomZ)。 |
||
表 1: 本研究中使用的细菌菌株清单。
| 质 粒 | 相关特点 | 参考 |
| pAH7 | PnatparB-yfp;Mx8 attP;春节 | 19 |
| pAH53 | PpomZ mCherry pomX;Mx8 attP ;KmR | 16 |
| pDS150 1 | PnatftsZ-gfp ;mxan18-19 ;春节 | 本研究 |
| pMR3691 | 香草酸诱导基因表达的质粒 | 18 |
| pKA51 | PnatftsZ-gfp ;Mx8 attP;春节 | 17 |
| 1 pDS150: pDS150 是 pKA51 的衍生物, Mx8 attP的位置被mxan18-19基因间区域所取代。 为此mxan18-19基因间区域从 pMR3691 被放大了与引物 mxan18-19 批转 BsdRI (GCGATCATTGCGCGCCAGACGATAACAGGC) 和 Mxan18-19 BlpI (GCGGCTGAGCCCGCGCCGACAACCGCAACC) 和克隆成 pKA51。 |
||
表 2: 本研究中使用的质粒清单。
Discussion
荧光活细胞成像已成为研究细菌细胞时空动态的有力工具。然而, 诸如m. xanthus这样的运动和慢生长细菌的时间失效荧光显微术一直具有挑战性, 只在短时间内进行。在这里, 我们提出了一个易于使用和稳健的方法的活细胞成像的m. xanthus的时间推移荧光显微镜。这种方法允许用户遵循细胞和荧光标记蛋白质的几个回合的细胞周期与单细胞分辨率。
有几个先决条件, 影响生长缓慢的m. xanthus细胞的活细胞成像的成功, 包括: 1) 一个固体表面的细胞附着;2) 养分和氧气的供应;3) 恒定的湿度和温度;4) 实验条件的优化, 如曝光时间和图像采集频率。
在我们的实验设置中, 我们使用厚的琼脂糖垫补充养分。使用厚的琼脂糖垫, 而不是微流控装置来跟踪单个细胞具有一些根本的好处, 但也有一些缺点。首先, 琼脂糖垫不仅为m xanthus细胞的附着和运动提供了表面, 而且还有足够的养分促进生长至少24小时。第二, 通常用于研究荧光标记蛋白的细胞内定位的快照分析, 以前是在同一类型的琼脂糖垫16,17,29。因此, 从快照分析中得到的数据可以与此处描述的方法直接比较。第三, 琼脂糖垫可以很容易地修改和补充抗生素或其他补充物, 如丘索4和香草酸, 通常用于基因表达诱导18,30。最后, 由于细胞在实验过程中被允许形成 microcolonies, 这种设置还允许研究直接细胞间相互作用对特定参数的影响。这方面是特别重要的情况下, m. xanthus , 因为这种细菌显示几个接触依赖的相互作用。该方法的主要缺点是实验条件是在实验期间预设的。相比之下, 微流控装置通常允许在实验过程中通过添加抗生素31来改变实验条件。
免费的软件包 (例如, MicrobeJ, Oufti) 可以自动分析单个细胞的生长和蛋白质的定位。然而, 这些软件只适用于对单个细胞或小组细胞的分析。因此, 自动分析此处描述的 24 h 记录所生成的数据仍然是一个挑战。
总之, 我们描述了一个易于使用的和可重现的协议, 执行活细胞成像与缓慢增长的m. xanthus细菌。我们表明, 简单的营养补充琼脂糖垫足以维持至少24小时的增长, 并允许观察和分析蛋白质的定位和生长, 单细胞分辨率数代。
Disclosures
作者声明他们没有竞争的财政利益。
Acknowledgments
这项工作得到德国研究委员会 (DFG) 在 Transregio 174 "细菌细胞时空动态" 和普朗克社会的框架内的支持。
Materials
| Name | Company | Catalog Number | Comments |
| DMI6000B with AFC | Leica microsystems | 11888945 | Automated inverted widefield fluorescence microscope with adaptive focus control |
| Universal mounting frame | Leica microsystems | 11532338 | Stage holder for different sample sizes |
| HCX PL FLUOTAR 100x/1.30 oil PH3 | Leica microsystems | 11506197 | Phase contrast objective |
| Orca Flash 4.0 camera | Hamamatsu | 11532952 | 4.0 megapixel sCMOS camera for picture aquisition |
| Filter set TXR ET, k | Leica microsystems | 11504170 | Fluorescence filter set, Ex: 560/40 Em: 645/75 |
| Filter set L5 ET, k | Leica microsystems | 11504166 | Fluorescence filter set, Ex: 480/40 Em: 527/30 |
| Filter set YFP ET, k | Leica microsystems | 11504165 | Fluorescence filter set, Ex: 500/20 Em: 535/30 |
| ProScan III | Prior | H117N1, V31XYZEF, PS3J100 | Microscope automation controller with interactive control center |
| EL 6000 light source | Leica microsystems | 11504115 | External fluorescence light source |
| Incubator BLX Black | Pecon | 11532830 | Black incubation chamber surrounding the microscope |
| Tempcontrol 37-2 digital | Leica microsystems | 11521719 | Automated temperature control for incubation chamber |
| Gentmycin sulphate | Carl Roth | 0233.4 | Gentamycin |
| Oxytetracylin dihydrate | Sigma Aldrich | 201-212-8 | Oxytetracyclin |
| Kanamycin sulphate | Carl Roth | T832.3 | Kanamycin |
| Filtropur BT25 0.2 bottle top filter | Sarstedt | 831,822,101 | Bottle top filter for sterilization of buffers |
| Deckgläser | VWR | 630-1592 | Glass cover slip (60 x 22 mm, thickness: 0.7 mm) |
| Seakem LE agarose | Lonza | 50004 | Agarose for microscopy slides |
| Leica Metamorph AF | Leica microsystems | 11640901 | Microscope control software and software for picture analysis |
| Tetraspeck Microsperes, 0.5 µm | ThermoFisher | T7281 | Fluorescent microspheres |
| petri dish | Greiner Bio-one | 688102 | 120 mm x 120 mm x 17 mm squared petri dish for agarose pads |
| BD Bacto Casitone | Becton Dickinson | 225930 | Casitone |
| Parafilm M | VWR | 291-1213 | Parafilm |
| Tris(hydroxymethyl)-aminomethane | Carl Roth | AE15.2 | Tris |
| Magnesium sulphate heptahydrate | Carl Roth | P027.2 | Magnesium sulphate |
| Potassium dihydrogen phosphate p.a. | Carl Roth | 3904.1 | Potassium dihydrogen phosphate |
| 1% CTT medium: 1 % (w/v) BD Bacto™ casitone, 10 mM Tris-HCl ph 8.0, 1 mM potassium phosphate buffer pH 7.6, 8 mM MgSO4 | Cultivation medium for M.xanthus | ||
| TPM buffer: 10 mM Tris-HCl ph 8.0, 1 mM potassium phosphate buffer pH 7.6, 8 mM MgSO4 | Buffer for preparation of microscopy slides for M.xanthus |
References
- Shapiro, L., McAdams, H. H., Losick, R. Why and how bacteria localize proteins. Science. 326 (5957), 1225-1228 (2009).
- Treuner-Lange, A., Søgaard-Andersen, L. Regulation of cell polarity in bacteria. J Cell Biol. 206 (1), 7-17 (2014).
- Laloux, G., Jacobs-Wagner, C. Spatiotemporal control of PopZ localization through cell cycle-coupled multimerization. J Cell Biol. 201, 827-841 (2013).
- Rudner, D. Z., Losick, R.
- Badrinarayanan, A., Le, T. B. K., Laub, M. T. Bacterial chromosome organization and segregation. Annu Rev Cell Dev Biol. 31 (1), 171-199 (2015).
- Munoz-Dorado, J., Marcos-Torres, F. J., Garcia-Bravo, E., Moraleda-Munoz, A., Perez, J. Myxobacteria: Moving, Killing, Feeding, and Surviving Together. Front Microbiol. 7, 781 (2016).
- Berleman, J. E., Kirby, J. R. Deciphering the hunting strategy of a bacterial wolfpack. FEMS Microbiol Rev. 33 (5), 942-957 (2009).
- Konovalova, A., Petters, T., Søgaard-Andersen, L.
- Nudleman, E., Wall, D., Kaiser, D. Cell-to-cell transfer of bacterial outer membrane lipoproteins. Science. 309, 125-127 (2005).
- Vassallo, C. N., et al. Infectious polymorphic toxins delivered by outer membrane exchange discriminate kin in myxobacteria. eLife. 6, 29397 (2017).
- Vassallo, C., et al. Cell rejuvenation and social behaviors promoted by LPS exchange in myxobacteria. Proc Natl Acad Sci USA. 112 (22), 2939-2946 (2015).
- Li, Y., et al. Extracellular polysaccharides mediate pilus retraction during social motility of Myxococcus xanthus. Proc. Natl. Acad. Sci. USA. 100, 5443-5448 (2003).
- Kim, S. K., Kaiser, D. Cell alignment required in differentiation of Myxococcus xanthus. Science. 249, 926-928 (1990).
- Lobedanz, S., Søgaard-Andersen, L. Identification of the C-signal, a contact dependent morphogen coordinating multiple developmental responses in Myxococcus xanthus. Genes Dev. 17, 2151-2161 (2003).
- Schumacher, D., Søgaard-Andersen, L. Regulation of cell polarity in motility and cell division in Myxococcus xanthus. Annu Rev Microbiol. 71 (1), 61-78 (2017).
- Schumacher, D., et al. The PomXYZ proteins self-organize on the bacterial nucleoid to stimulate cell division. Dev Cell. 41 (3), 299-314 (2017).
- Treuner-Lange, A., et al. PomZ, a ParA-like protein, regulates Z-ring formation and cell division in Myxococcus xanthus. Mol Microbiol. 87 (2), 235-253 (2013).
- Iniesta, A. A., Garcia-Heras, F., Abellon-Ruiz, J., Gallego-Garcia, A., Elias-Arnanz, M. Two systems for conditional gene expression in Myxococcus xanthus inducible by isopropyl-beta-D-thiogalactopyranoside or vanillate. J Bacteriol. 194 (21), 5875-5885 (2012).
- Harms, A., Treuner-Lange, A., Schumacher, D., Søgaard-Andersen, L. Tracking of chromosome and replisome dynamics in Myxococcus xanthus. reveals a novel chromosome arrangement. PLoS Genet. 9 (9), 1003802 (2013).
- Iniesta, A. A. ParABS system in chromosome partitioning in the bacterium Myxococcus xanthus. PLoS One. 9 (1), 86897 (2014).
- Lin, L., Osorio Valeriano, M., Harms, A., Søgaard-Andersen, L., Thanbichler, M. Bactofilin-mediated organization of the ParABS chromosome segregation system in Myxococcus xanthus. Nat Commun. 8 (1), 1817 (2017).
- Hodgkin, J., Kaiser, D. Cell-to-cell stimulation of movement in nonmotile mutants of Myxococcus. Proc Natl Acad Sci U S A. 74 (7), 2938-2942 (1977).
- Kaiser, D. Social gliding is correlated with the presence of pili in Myxococcus xanthus. Proc Natl Acad Sci USA. 76 (11), 5952-5956 (1979).
- Miertzschke, M., et al. Structural analysis of the Ras-like G protein MglA and its cognate GAP MglB and implications for bacterial polarity. EMBO J. 30 (20), 4185-4197 (2011).
- Hodgkin, J., Kaiser, D. Genetics of gliding motility in Myxococcus xanthus. (Myxobacterales): Two gene systems control movement. Mol Gen Genet. 171, 177-191 (1979).
- Shi, X., et al. Bioinformatics and experimental analysis of proteins of two-component systems in Myxococcus xanthus. J Bacteriol. 190 (2), 613-624 (2008).
- Bi, E. F., Lutkenhaus, J. FtsZ ring structure associated with division in Escherichia coli. Nature. 354 (6349), 161-164 (1991).
- Lutkenhaus, J., Pichoff, S., Du, S. Bacterial cytokinesis: From Z ring to divisome. Cytoskeleton. 69 (10), 778-790 (2012).
- McLoon, A. L., et al. MglC, a Paralog of Myxococcus xanthus GTPase-Activating Protein MglB, Plays a Divergent Role in Motility Regulation. J Bacteriol. 198 (3), 510-520 (2015).
- Gomez-Santos, N., et al. Comprehensive set of integrative plasmid vectors for copper-inducible gene expression in Myxococcus xanthus. Appl Environ Microbiol. 78 (8), 2515-2521 (2012).
- Treuner-Lange, A., et al. The small G-protein MglA connects to the MreB actin cytoskeleton at bacterial focal adhesions. J Cell Biol. 210 (2), 243-256 (2015).
