

Summary
हम खुबानी की परागण आवश्यकताओं को स्थापित करने के लिए एक पद्धति प्रस्तुत करते हैं(प्रूनस आर्मेनियाका एल.) ऐसी खेती जो पीसीआर विश्लेषण द्वारा एस-जीनोटाइप की पहचान के साथ फ्लोरेसेंस माइक्रोस्कोपी द्वारा आत्म-(इन) अनुकूलता के निर्धारण को मिलाकर है।
Abstract
रोसासी में आत्म-असंगति एक गेमटोफाइटिक सेल्फ-इनकंकेटिबिलिटी सिस्टम (जीएसआई) द्वारा निर्धारित की जाती है जो मुख्य रूप से मल्टीलेलिक लोकस एस द्वारा नियंत्रित होती है। खुबानी में, आत्म-और अंतर-और अंतर-अनुकूलता संबंधों का निर्धारण तेजी से महत्वपूर्ण है, क्योंकि एक महत्वपूर्ण संख्या में नई खेती जारी करने के परिणामस्वरूप अज्ञात परागण आवश्यकताओं के साथ खेती में वृद्धि हुई है। यहां, हम एक ऐसी पद्धति का वर्णन करते हैं जो पीसीआर विश्लेषण द्वारा एस-जीनोटाइप की पहचान के साथ हाथ-परागण और माइक्रोस्कोपी द्वारा आत्म-(इन)अनुकूलता के दृढ़ संकल्प को जोड़ती है। स्व-(में) अनुकूलता निर्धारण के लिए, प्रत्येक खेती से गुब्बारे के चरण में फूल क्षेत्र में एकत्र किए गए थे, प्रयोगशाला में हाथ से परागण, निश्चित, और फ्लोरेसेंस माइक्रोस्कोपी के तहत पराग ट्यूब व्यवहार के अवलोकन के लिए एनीलिन नीले रंग के साथ दाग। खेती के बीच असंगति संबंधों की स्थापना के लिए, प्रत्येक खेती से डीएनए युवा पत्तियों से निकाला गया था और पीसीआरद्वारा एस-एलील्स की पहचान की गई थी । यह दृष्टिकोण असंगति समूहों की स्थापना और खेती के बीच असंगति संबंधों को स्पष्ट करने की अनुमति देता है, जो नए बगीचों के डिजाइन में उपयुक्त परागणकों का चयन करने और प्रजनन कार्यक्रमों में उपयुक्त माता-पिता का चयन करने के लिए एक मूल्यवान जानकारी प्रदान करता है।
Introduction
आत्म-असंगति आत्म-परागण को रोकने और आउटक्रॉसिंग1को बढ़ावा देने के लिए फूलों के पौधों की एक रणनीति है। रोसासी में, यह तंत्र एक गेमटोफाइटिक सेल्फ-इनकंकेटिबिलिटी सिस्टम (जीएसआई) द्वारा निर्धारित किया जाता है जो मुख्य रूप से मल्टीलेलिक लोकस एस2 द्वारानियंत्रितहोता है। शैली में, RNase जीन एस-एसtylar निर्धारक, एक RNase3,जबकि एक एफ बॉक्स प्रोटीन, जो एसपराग निर्धारक निर्धारित करता है, SFB जीन4द्वारा संहिताबद्ध है । आत्म-असंगति बातचीत ओव्यूल5,,6के निषेचन को रोकने वाली शैली के साथ पराग ट्यूब विकास के अवरोध के माध्यम से होती है।
खुबानी में, पिछले दो दशकों में दुनिया भर में एक किस्म का नवीकरण किया गया है7,8। विभिन्न सार्वजनिक और निजी प्रजनन कार्यक्रमों से नई खेती की एक महत्वपूर्ण संख्या की इस शुरूआत के परिणामस्वरूप अज्ञात परागण आवश्यकताओं के साथ खुबानी खेती की वृद्धि हुई है8।
खुबानी में परागण आवश्यकताओं को निर्धारित करने के लिए विभिन्न तरीकों का उपयोग किया गया है। क्षेत्र में, बंदी वृक्षों या नपुंसक फूलों में नियंत्रित परागण द्वारा आत्म-(इन) अनुकूलता स्थापित की जा सकती है और बाद में फल सेट9,10, 11,,,12के प्रतिशत को रिकॉर्ड किया जा11सकताहै। इसके अलावा, प्रयोगशाला में फूलों की अर्ध-इन वीवो संस्कृति और फ्लोरेसेंस माइक्रोस्कोपी,,8,,13, 14, 15, 16,,14,17के तहत पराग ट्यूब व्यवहार के विश्लेषण द्वारानियंत्रितपरागण किए गए हैं।17 हाल ही में, पीसीआर विश्लेषण और अनुक्रमण जैसी आणविक तकनीकों ने आरएनएनई और एसएफबी जीन18, 19,19के अध्ययन के आधार पर असंगति संबंधों के लक्षण वर्णन की अनुमति दी है। खुबानी में, 33 एस-एलील्स(एस1,से एस20,एस,22 से एस S30,एस52, एस53, एसवी,22 एस, एसएक्स)की सूचना दी गई है,जिसमें स्व-अनुकूलता(एससी)12, 18, 20,21,,2122,,2023, 24से संबंधित एक एलील शामिल हैं।,,24 अब तक, एस- जीनोटाइप 8 ,9, 17 , 25 , 26 ,,,278केअनुसार इस प्रजाति में,1726असंगति समूहों को स्थिर किया गया है ।26 एक ही एस-एलील्सके साथ खेती अंतर-असंगत हैं, जबकि कम से कम एक अलग एस-एलीलके साथ खेती और, नतीजतन, विभिन्न असंगत समूहों में आवंटित, अंतर-संगत हैं।
खुबानी खेती की परागण आवश्यकताओं को परिभाषित करने के लिए, हम एक पद्धति का वर्णन करते हैं जो खुबानी खेती में पीसीआर विश्लेषण द्वारा एस-जीनोटाइप की पहचान के साथ फ्लोरेसेंस माइक्रोस्कोपी द्वारा आत्म-(इन)अनुकूलता के दृढ़ संकल्प को जोड़ती है। यह दृष्टिकोण असंगति समूहों की स्थापना और खेती के बीच असंगति संबंधों को स्पष्ट करने की अनुमति देता है ।
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. आत्म-(में) अनुकूलता दृढ़ संकल्प
- खेत में फूलों का नमूना लिया। अवांछित पिछले परागण से बचने के लिए खुबानी28के लिए बीबीसीएच पैमाने पर चरण 58 के अनुरूप गुब्बारे के चरण(चित्रा 1ए)पर फूलों को इकट्ठा करना आवश्यक है।
-
प्रयोगशाला में स्वाध्याय और क्रॉस-परागण
- गुब्बारे के चरण में फूलों के एंथर्स को हटा दें और प्रयोगशाला के तापमान पर सूखने के लिए उन्हें कागज के टुकड़े पर रखें।
- 24 घंटे के बाद, एक बारीक जाल (0.26 मिमी) (चित्र 1बी)का उपयोग करके परागकणों को छलनी करें।
- प्रत्येक आत्म-परागण और क्रॉस-परागण के लिए एक ही गुब्बारे के चरण में 30 फूलों के एक समूह को कम करें और प्रयोगशाला के तापमान(चित्र 1सी)पर पानी में फूलों के फोम पर पिस्टिल रखें।
- हाथ से एक ही खेती 24 घंटे के फूलों से पराग के साथ एक तूलिका की मदद से पिस्टिलों को परागण करें। इसके अलावा, नियंत्रण के रूप में एक संगत परागणाइजर के फूलों से पराग के साथ प्रत्येक खेती के पिस्टिल्स का एक और सेटपरागण (चित्रा 1डी)।
- 72 घंटे के बाद, 4 डिग्री सेल्सियस29पर कम से कम 24 घंटे के लिए इथेनॉल/एसिटिक एसिड (3:1) के एक स्थिर समाधान में पिस्टिल्स को ठीक करें। फिर फिक्सेटिव को त्यागें और यह सुनिश्चित करते हुए 75% इथेनॉल जोड़ें कि नमूने पूरी तरह से समाधान में डूबे हुए हैं। इस घोल,में 8 ,17, 30 , 31,,32का उपयोग होने तक नमूनों को4डिग्री सेल्सियस पर संरक्षित किया जा8सकता32है ।
-
इन विट्रो पराग अंकुरण के माध्यम से पराग व्यवहार्यता का मूल्यांकन
- अंकुरण माध्यम तैयार करने के लिए, वजन 25 ग्राम सुक्रोज, 0.075 ग्राम बोरिक एसिड (एच3बीओ3)और 0.075 ग्राम कैल्शियम नाइट्रेट (सीए (नंबर3)2)33।
- आसुत पानी के 250 एमएल में माध्यम के घटकों को जोड़ें और पूरी तरह से भंग करें।
- 2 ग्राम एगर उठे और घूमता द्वारा मिश्रण जोड़ने के माध्यम को जमना।
- पीएच मीटर का उपयोग करके माध्यम के पीएच की जांच करें और मूल्य को नाओएच या एचसीएल समाधान के साथ 7.0 तक समायोजित करें।
- माध्यम को स्टरलाइज करने के लिए मिश्रण को ऑटोक्लेव करें।
- ऑटोक्लेविंग के बाद, माध्यम को ठंडा करें और इसे बाँझ लेमिनार प्रवाह हुड में पेट्री व्यंजनों में वितरित करें।
- जमते पराग अंकुरण माध्यम में नियंत्रित परागण के लिए उपयोग की जाने वाली उन्हीं खेती के पराग कणों को तितर - बितर करें और 24 घंटे 6 के बाद उन्हें माइक्रोस्कोप केनीचेदेखें ।
नोट: लैमिनार प्रवाह हुड को स्टरलाइज करने के लिए, सतह को 70% इथेनॉल के साथ साफ करें और 10 मिनट के दौरान यूवी लैंप पर स्विच करें। - पेट्री डिश को 4 डिग्री सेल्सियस पर फ्रिज में तब तक स्टोर करें जब तक कि इस्तेमाल न हो जाए।
-
माइक्रोस्कोपी अवलोकन
- पिस्टिल्स को डिस्टिल्ड वॉटर के साथ 1 एच के लिए तीन बार धोएं और उन्हें 4 डिग्री सेल्सियस पर 5% सोडियम सल्फेट में छोड़ दें। 24 घंटे के बाद, ऊतकों को नरम करने के लिए सोडियम सल्फेट में 10 मिनट केदौरान उन्हें 1 किलो/सेमी 2 पर ऑटोक्लेवकरें ३४।
- ऑटोक्लेवेड पिस्टिल को एक ग्लास स्लाइड पर रखें और, स्केलपेल की मदद से, पराग ट्यूबों का बेहतर दृश्य प्राप्त करने के लिए अंडाशय के चारों ओर ट्राइहोम हटा दें। इसके बाद पिस्टिलों को कवर ग्लास से स्क्वैश करें।
- तैयार करें 0.1% (v/v) एनलीन ब्लू दाग: 0.1 एन पोटेशियम फॉस्फेट ट्राइबेसिक (के 3 पीओ 4) के 100 मिलील में एनलीन ब्लू का0.1एमएल मिलाएं।4 पराग ट्यूब वृद्धि के दौरान कैलिओस जमाव को दाग देने की तैयारी के ऊपर एनीलिन नीले रंग की एक बूंद लागू करें।
- 340-380 बैंडपास और 425 लॉन्गपास फिल्टर का उपयोग करके यूवी एपिफ्लोरेसेंस के साथ एक माइक्रोस्कोप द्वारा शैली के साथ पराग ट्यूबों का निरीक्षण करें।
2. डीएनए निष्कर्षण
- वसंत ऋतु में खेत में 2-3 पत्तियों का नमूना। युवा चरणों में पत्तियों का नमूना लेने की सिफारिश की जाती है क्योंकि प्राप्त डीएनए पुरानी पत्तियों की तुलना में उच्च गुणवत्ता और फेनोलिक यौगिकों के निचले स्तर का होता है।
- व्यावसायिक रूप से उपलब्ध किट (सामग्रीकी तालिका देखें) में वर्णित चरणों के बाद जीनोमिक डीएनए निकालें।
- यूवी-विस स्पेक्ट्रोफोटोमीटर (260 एनएम) का उपयोग करके डीएनए सांद्रता की मात्रा और गुणवत्ता का विश्लेषण करें।
3. Sएस-एलील पहचान
-
पीसीआर प्रतिक्रियाओं की स्थापना
- प्रत्येक डीएनए निकासी नमूने के आसुत पानी में 50 एनजी/μL कमजोर पड़ने की तैयारी करें।
- पीसीआर रिएजेंट्स को धीरे-धीरे गल कर उन्हें बर्फ पर रखें। जरूरत होने तक डीएनए पॉलीमरेज को फ्रीजर में छोड़ दें।
- प्राइमर के विभिन्न संयोजनों का उपयोग करके प्रवर्धन प्रतिक्रियाओं को तैयार करें। टेबल 1में घटकों को मिलाकर पीसीआर रिएक्शन मिक्स बनाएं । भंवर पीसीआर प्रतिक्रिया अच्छी तरह से मिश्रण और पीसीआर प्लेट के प्रत्येक कुएं के लिए प्राइमर के विभिन्न संयोजनों के लिए संकेत दिया मात्रा वितरित। फिर, प्रत्येक कुएं में डीएनए कमजोर पड़ने का 1 माइक्रोल जोड़ें।
- पीसीआर प्लेट को थर्मोसाइकिलर में रखें और टेबल 1 में दिखाए गए संबंधित पीसीआर कार्यक्रम को चलाएं।
- प्रवर्धित टुकड़ों का विश्लेषण करें। पीसीआर प्रवर्धित टुकड़ों का विश्लेषण करने के लिए मुख्य रूप से दो अलग-अलग तरीके हैं: फ्लोरोसेंट-लेबल वाले प्राइमर के साथ केशिका इलेक्ट्रोफोरेसिस (सीई) या बिना लेबल वाले प्राइमर के साथ एगर उठे जेल इलेक्ट्रोफोरेसिस के एम्प्लिकॉन की कल्पना के रूप में।
-
केशिली इलेक्ट्रोफोरेसिस
- लोडिंग बफर तैयार करने के लिए, लेबल आकार मानक के 0.45 माइक्रोन के साथ डिएकोनाइज्ड फॉर्ममाइड के 35 माइक्रोन मिलाएं। भंवर अभिवाची मिश्रण करने के लिए अभिवाक है, और फिर पाठक प्लेट के कुएं में 35.5 μL बांटना।
- पीसीआर उत्पाद के 1 माइक्रोन को कुएं में जोड़ें। इसके अलावा, पानी वाष्पीकरण को रोकने के लिए खनिज तेल की एक बूंद जोड़ें।
- सेपरेशन प्लेट को सेपरेशन बफर जोड़ते हुए तैयार करें।
- जीन एनालाइजर (सामग्री की तालिकादेखें) के साथ शामिल वाणिज्यिक सॉफ्टवेयर का उपयोग करें। एक नया नमूना प्लेट बनाएं और प्लेट पर सभी कुओं के लिए नमूना नाम बचाएं।
- विश्लेषण की विधि का चयन करें। इस मामले में, नमूनों को 120 एस के लिए 90 डिग्री सेल्सियस पर, 30 एस के लिए 2.0 केवी पर इंजेक्ट करें, और 35 मिनट के लिए 6.0 केवी पर अलग करें।
- जीन एनालाइजर में दो प्लेट डालें। आसुत पानी के साथ केशिका सरणी भरें।
- पेटेंट लीनियर पॉलीएक्रेलैमाइड (एलपीए) जेल लोड करें। अंत में, रनपर क्लिक करें ।
-
जेल इलेक्ट्रोफोरेसिस
- 1x TAE (Tris-एसीटेट-एडटा) इलेक्ट्रोफोरेसिस रनिंग बफर (40 mM Tris, 20 mM एसिटिक एसिड, और पीएच 8.0 पर 1 m EDTA) के 150 एमएल में आणविक जीव विज्ञान ग्रेड के 1.5 ग्राम जोड़ने वाले 1%एगर उठे जेल तैयार करें। 2-3 मिनट के लिए माइक्रोवेव हीटिंग से एगर उठे को भंग करें।
- डीएनए की कल्पना करने के लिए, एक न्यूक्लिक एसिड दाग के 4 माइक्रोन जोड़ें (सामग्री की तालिका देखें) और धीरे-धीरे मिलाएं।
- एक जेल ट्रे में सीढ़ी, नियंत्रण और नमूनों के लिए पर्याप्त कुओं के साथ एक जेल कंघी जोड़ें। फिर, धीरे-धीरे मिश्रण को जेल ट्रे के बीच में डालें और बुलबुले से बचें।
- जेल को कमरे के तापमान पर 30-45 मिनट तक ठंडा होने दें जब तक कि जेल पूरी तरह से जम न जाए। इलेक्ट्रोफोरेसिस कक्ष में जेल का परिचय दें, जेल कंघी को हटा दें और जेल को कवर करने के लिए पर्याप्त 1x टीएई बफर के साथ कक्ष को भरें।
नोट: जेल के प्लेसमेंट की जांच करें। कुओं को नकारात्मक ध्रुव के करीब रखा जाना चाहिए क्योंकि नकारात्मक रूप से आवेशित डीएनए कैथोड की ओर प्रवास करता है। - पीसीआर उत्पादों में 5 माइक्रोन लोडिंग बफर (0.1% (v/v) ब्रोमोफेनॉल ब्लू) जोड़ें और अच्छी तरह से मिलाएं।
- बैंड के आकार का अनुमान लगाने के लिए, डीएनए आणविक वजन सीढ़ी के 5 माइक्रोन लोड (सामग्री की तालिकादेखें)।
- नमूनों को जेल के अतिरिक्त कुओं में लोड करें।
- एक बार सभी नमूनों और डीएनए आणविक वजन सीढ़ी लोड कर रहे हैं, 1-1.5 घंटे के लिए ९० वी पर जेल चलाने के लिए, जब तक नीले रंग की लाइन लगभग ७५% जेल की लंबाई पर है ।
- न्यूक्लिक एसिड के लिए एक ट्रांसिल्यूमिनेटर में बैंड की कल्पना करें।
-
केशिली इलेक्ट्रोफोरेसिस
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
खुबानी में परागण अध्ययन एंथेसिस(चित्रा 1ए)से एक दिन पहले देर से गुब्बारे के चरण में फूलों के उपयोग की आवश्यकता होती है । इस चरण को पराग और पिस्टिल संग्रह दोनों के लिए सबसे अनुकूल माना जाता है, क्योंकि पुष्प संरचनाएं लगभग परिपक्व हैं, लेकिन एंथर डीहिसेंस अभी तक नहीं हुई है। यह न केवल एक ही फूल से बल्कि अन्य फूलों से भी पराग के अवांछित पराग के हस्तक्षेप को रोकता है, क्योंकि बंद पंखुड़ियां बाहरी पराग ले जाने वाले कीड़ों के आगमन में बाधा डालती हैं। पराग कणों को आसानी से एक ठीक जाल(चित्रा 1बी)के माध्यम से प्राप्त किया जाता है, जो पहले कमरे के तापमान पर या मामूली अतिरिक्त गर्मी के साथ 24 घंटे के लिए कागज के टुकड़े पर रखा जाता था। इसी तरह, चिमटी या नाखूनों(चित्रा 1सी)की मदद से पंखुड़ियों, सेपल्स और पुंकेसर को हटाने के बाद गुब्बारे के चरण में फूलों से पिस्टिल प्राप्त किए जाते हैं। पिस्टिल्स को एक ठीक ब्रश(चित्रा 1डी) के साथ आत्म-और क्रॉस-परागण किया जा सकताहै।
खुबानी के हर्मेफ्रोडिटिक फूलों में पांच गहरे लाल सेपल्स, पांच सफेद पंखुड़ियां(चित्रा 1ए),एक एकल पिस्टिल(चित्रा 2ए)और 25-30 पुंकेसर होते हैं। पिस्टिल में तीन मुख्य संरचनाएं हैं: कलंक, शैली और अंडाशय। अंडाशय में दो अंडाशय होते हैं, और फलों की स्थापना के लिए उनमें से कम से कम एक का निषेचन आवश्यक है। परागण के दौरान, कीड़े, मुख्य रूप से मधुमक्खियां, पराग कणों को कलंक(चित्रा 1ए)में स्थानांतरित करती हैं, जहां वे परागण के बाद 24 घंटे के भीतर(चित्रा2बी)अंकुरित होते हैं। प्रत्येक अंकुरित पराग कण से एक पराग ट्यूब का उत्पादन होता है, जो 3-4 दिनों के बाद अंडाशय तक पहुंचने के लिए पिस्टिल संरचनाओं के माध्यम से बढ़ता है और लगभग 7 दिनों के बाद दो अंडाशय में से एक को उपजाऊ बनाता है। आत्म-असंगत खेती में, जिसमें पराग कण का एस एलील दो पिस्टिलों में से एक के समान है, पराग ट्यूब ऊपरी शैली में बढ़ना बंद कर देता है, निषेचन कोरोकता है (चित्रा 2सी)। हालांकि, एक अलग एस एलील के साथ एक संगत खेती से पराग ट्यूब, शैली(चित्रा 2डी)के माध्यम से विकसित हो सकते हैं, अंडाशय(चित्रा 2ई)तक पहुंचते हैं और दो अंडाशय में से एक को उर्वरित कर सकते हैं।
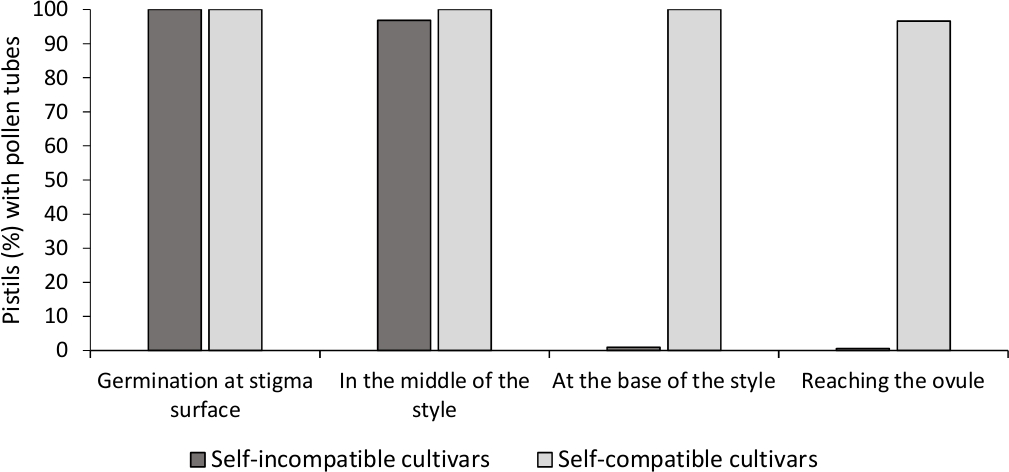
इन विट्रो पराग अंकुरण के विश्लेषण से यहां विश्लेषण की गई सभी खेती में अच्छी पराग व्यवहार्यता दिखाई दी, क्योंकि अधिकांश पराग ट्यूब संस्कृति माध्यम में 24 घंटे के बाद पराग कण की लंबाई से अधिक लंबी थीं। सभी परागणों से पिस्टिन में कलंक की सतह(चित्र 2बी)पर अंकुरित पराग कण देखे गए, जो पर्याप्त परागण(चित्र 3) का संकेत देते हैं।
प्रत्येक खेती के लिए आत्म-(इन) अनुकूलता निर्धारित करने के लिए, फ्लोरेसेंस माइक्रोस्कोपी के तहत प्रयोगशाला नियंत्रित स्थितियों में किए गए स्वयं और क्रॉस-परागण में परागण में परागणणणकता देखी गई। सभी पिस्टिलों की जांच में शैली के साथ पराग ट्यूब विकास दर्ज किया गया था। खेती को आत्म-असंगत माना जाता था जब पराग ट्यूब विकास को शैली के साथ सबसे अधिक परागणित पिस्टिल्स(चित्रा 2सी, चित्रा 3)और आत्म-संगत में गिरफ्तार किया गया था जब कम से कम एक पराग ट्यूब अधिकांश पिस्टिल्स में शैली के आधार तक पहुंच गया(चित्रा 2ई,चित्र 3)।
पीसीआर विश्लेषण द्वारा एस-लोकसके अध्ययन ने प्रत्येक खेती के एस-जीनोटाइपकी विशेषता की अनुमति दी। सबसे पहले, Sएस-एलील्स की पहचान प्राइमर एसआरसी-एफ/एसआरसी-आर (टेबल 2)का उपयोग करके पहले एस-आरएनएज़ इंट्रॉन के प्रवर्धन द्वारा की गई थी । प्रवर्धित टुकड़ों के आकार का विश्लेषण केशिका इलेक्ट्रोफोरेसिस(चित्रा 4ए)द्वारा किया गया था और उनके संबंधित असंगति समूह (आईजी) में विश्लेषण किए गए जीनोटाइप को वर्गीकृत करने के लिए उपयोग किया गया था। (टेबल3)।
एस 1 और एस 7 Sया एस6 और एस6 9 जैसे एलील्स केकुछ जोड़े, पहले इंस्टट्रॉन के लिए समान टुकड़ा आकार दिखाए गए। S9 इस प्रकार, इन एलील्स का भेदभाव प्राइमर प्रू-सी 2/प्र्यूसी4आर, एसएचएलएम1/एसएचएलएम 2 और एसएचएलएम 3/एसएचएलएम4 (टेबल 2)के साथ RNase के दूसरे इंट्रोन के एक क्षेत्र को बढ़ाना द्वारा किया गया था । PruC2/PruC4R प्राइमर संयोजन एस6 और एस9 के बीच अंतर करने के लिए इस्तेमाल किया गया था ।9 एस6के लिए, 1300 बीपी का एक टुकड़ा परिलक्षित किया गया था जबकि एस 9 एली(चित्रा 4 बी, तालिका 3)के लिए लगभग 700 बीपी का9 एक टुकड़ा देखा गया था। विशिष्ट प्राइमर SHLM1/SHLM2 और SHLM3/SHLM4 एस1 allele में लगभग ६५० बीपी का एक टुकड़ा परिलक्षित और एस7 allele में ४१३ बीपी(चित्रा 4सी,तालिका 3)।
एसएफबी जीन के V2 और HVb चर क्षेत्रों को बढ़ाना करने वाले प्राइमर AprFBC8-(F/R) का उपयोग एससी और एस8 एलील्स को अलग करने के लिए कियाजाता था क्योंकि दोनों एलील्स समान RNase अनुक्रम दिखाते हैं । एस8 एलील ने लगभग 150 बीपी का पीसीआर-टुकड़ा दिखाया जबकि 500 बीपी का टुकड़ा एससी एलील(चित्रा 4डी,टेबल 3)से मेल खाता था। एक Sबार एस-जीनोटाइप सभी खेती के लिए निर्धारित किया गया था, आत्म असंगत खेती उनके Sएस-alleles(तालिका 3)के आधार पर उनके इसी असंगति समूहों को सौंपा गया था ।
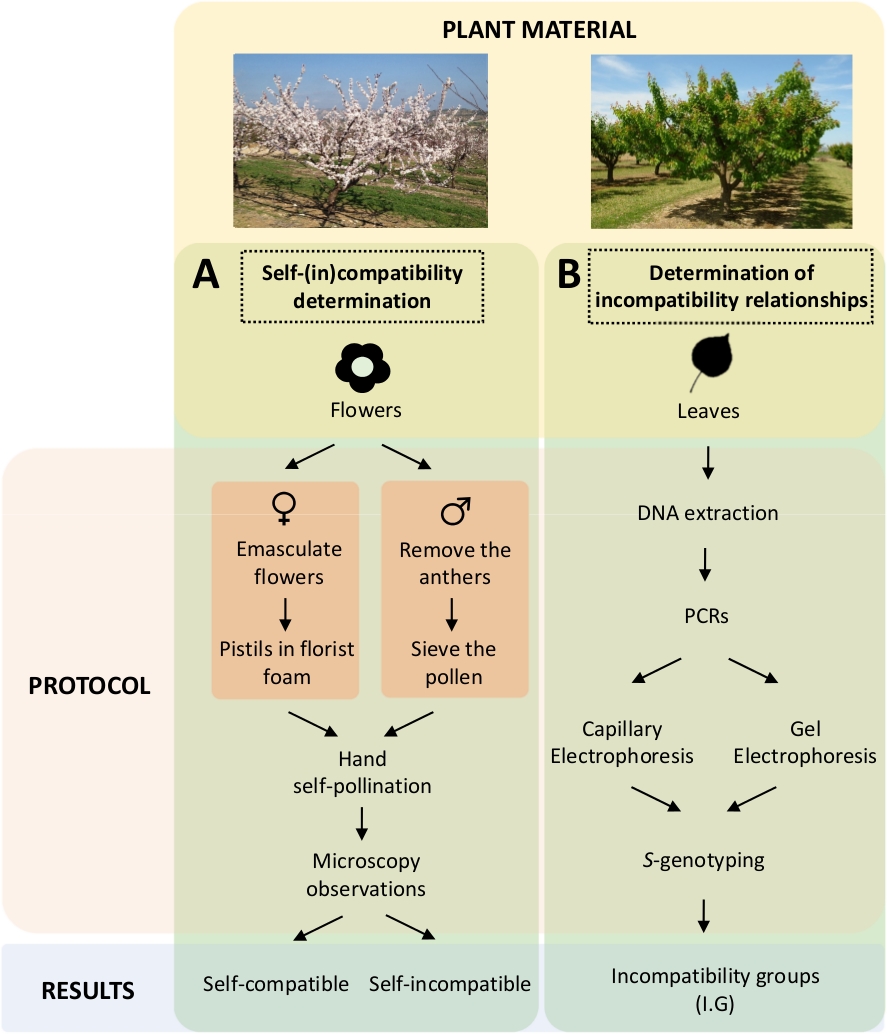
इस दृष्टिकोण के लिए प्रयोगशाला में नियंत्रित आत्म-और क्रॉस-परागण द्वारा प्रत्येक खेती की आत्म-(इन) अनुकूलता का निर्धारण करने की आवश्यकता है(चित्रा 5ए)आनुवंशिक विश्लेषण द्वारा एस-जीनोटाइप(चित्रा 5बी)द्वारा एस-जीनोटाइपके लक्षण वर्णन के साथ। नतीजतन, प्रत्येक खेती की परागण आवश्यकताओं और खुबानी खेती के बीच असंगति संबंधों को निर्धारित किया जा सकता है ।

चित्रा 1. खुबानी में आत्म-(में) अनुकूलता के निर्धारण के लिए प्रायोगिक स्थापित।
(A)खेत में गुब्बारे के मंच पर फूल (काले तीर) । (ख)एक बारीक जाल का उपयोग कर परागकणों की छलनी । (ग)पानी में फूलों के फोम पर रखे पिस्टिल। (घ)तूलिका की मदद से पिस्टिलों का हाथ से परागण। कृपया इस आंकड़े का एक बड़ा संस्करण देखने के लिए यहां क्लिक करें ।

चित्रा 2। खुबानी फूलों में गेमोफाइटिक असंगति संबंधों का आरेखीय प्रतिनिधित्व।
(क)गेमोफाइटिक सेल्फ-असंगति (जीएसआई) में, संगत और असंगत पराग दोनों कण कलंक पर अंकुरित होते हैं । पराग कण मूल जीनोटाइप के दो एस-एलील्समें से एक होता है, इस मामले में या तो एस1 या एस2। यदि पराग कण का एस-एलीलपिस्टिल के दो एस-एलील्समें से एक से मेल खाता है, तो इस मामले में एस1एस3,पराग ट्यूब वृद्धि शैली के ऊपरी एक तिहाई में बाधित होती है। (ख)कलंक की सतह पर परागकणों का अंकुरण। (ग)पराग ट्यूब एक असंगत व्यवहार का संकेत शैली में गिरफ्तार कर लिया । (घ)पराग ट्यूब शैली के साथ बढ़ रहा है । (ई)एक संगत व्यवहार का संकेत शैली के आधार पर पराग ट्यूब। स्केल बार, 100 माइक्रोन। कृपया इस आंकड़े का एक बड़ा संस्करण देखने के लिए यहां क्लिक करें ।

चित्र 3। पराग अंकुरण और पराग ट्यूब विकास के प्रतिनिधि परिणाम स्वयं के लिए शैली के माध्यम से संगत और आत्म-परागण के बाद आत्म-असंगत खेती।
पराग कणों के साथ पिस्टिल का प्रतिशत कलंक की सतह पर अंकुरित होता है, आधे रास्ते में पराग ट्यूबों के साथ शैली के आधार पर, और अंडाउल तक पहुंचता है। कृपया इस आंकड़े का एक बड़ा संस्करण देखने के लिए यहां क्लिक करें ।

चित्र 4. एस-एलील्स की पहचान के लिए पांच Sप्राइमर जोड़ी संयोजनों का उपयोग करके पीसीआर टुकड़ा प्रवर्धन।
(A)एसआरसी के लिए जीन एनालाइजर आउटपुट-(एफ/आर) प्राइमर एस-एलील्स के अनुरूप आरएनएएस फर्स्ट Sइंट्रॉन क्षेत्र के दो प्रवर्धित टुकड़ों के आकार को दिखाते हैं । (ख) एस6 और एस9 एलील्स की पहचान के लिए प्राइमर PruC2/PruC4R का उपयोग कर पीसीआर प्रवर्धन ।9 (ग)पीसीआर उत्पादों एस1 allele और SHLM3 और SHLM4 के भेदभाव के लिए विशिष्ट प्राइमर SHLM1 और SHLM2 का उपयोग कर प्राप्त करने के लिए एस7 allele भेद । (घ) एससी और एस8 एलील्स की पहचान के लिए AprFBC8-(F/R) प्राइमर के साथ पीसीआर प्रवर्धन । एममैं:1 केबी डीएनए सीढ़ी। एमद्वितीय:100 बीपी डीएनए सीढ़ी। कृपया इस आंकड़े का एक बड़ा संस्करण देखने के लिए यहां क्लिक करें ।

चित्रा 5। खुबानी खेती में स्व-और अंतर-(इन) अनुकूलता संबंधों को स्पष्ट करने के लिए प्रायोगिक डिजाइन की योजना।
(A)प्रयोगशाला में नियंत्रित परागण द्वारा आत्म-(इन) अनुकूलता निर्धारण का कार्यप्रवाह। (ख)आणविक दृष्टिकोणों द्वारा एस-एलीलपहचान का कार्यप्रवाह। कृपया इस आंकड़े का एक बड़ा संस्करण देखने के लिए यहां क्लिक करें ।
| पीसीआर मास्टर मिक्स | थर्मोसाइकिलर की स्थिति | ||||||
| घटक | अंतिम एकाग्रता | 15 माइक्रोल प्रतिक्रिया | साइकिल चरण | तापमान | समय | चक्र | |
| 10x एनएच4 रिएक्शन बफर | 10x | 1.5 माइक्रोन | प्रारंभिक विकृति | 94 डिग्री सेल्सियस | 3 मिनट | 1 | |
| 50 एमएमएम एमजीसीएल2 समाधान | 25 एमएमएम | 1.2 माइक्रोल | डेन्चरिंग | 94 डिग्री सेल्सियस | 1 मिनट | 35 | |
| 100 एमएमएम डीएनटीपी | 2.5 mM | 0.6 माइक्रोन | एनीलिंग | 55 डिग्री सेल्सियस | 1 मिनट | ||
| प्राइमर एसआरसी-एफ | 10 माइक्रोन | 0.6 माइक्रोन | एक्सटेंशन | 72 डिग्री सेल्सियस | 3 मिनट | ||
| प्राइमर एसआरसी-आर | 10 माइक्रोन | 0.6 माइक्रोन | अंतिम विस्तार | 72 डिग्री सेल्सियस | 5 मिनट | 1 | |
| 500 यू Taq डीएनए बहुलक | 0.5 यू | 0.2 माइक्रोल | 4 डिग्री सेल्सियस | पकड़ | |||
| एच2ओ | 8.3 माइक्रोन | ||||||
| घटक | अंतिम एकाग्रता | 25 माइक्रोल प्रतिक्रिया | साइकिल चरण | तापमान | समय | चक्र | |
| 10x पीसीआर बफर | 10x | 2.5 माइक्रोन | प्रारंभिक विकृति | 94 डिग्री सेल्सियस | 2 मिनट | 1 | |
| 5x क्यू-सॉल्यूशन | 5x | 5 माइक्रोन | डेन्चरिंग | 94 डिग्री सेल्सियस | 10 एस | 10 | |
| 100 एमएमएम डीएनटीपी | 2.5 mM | 0.5 माइक्रोन | एनीलिंग | 55 डिग्री सेल्सियस | 2 मिनट | ||
| प्राइमर प्रूसी2 | 10 माइक्रोन | 0.2 माइक्रोल | एक्सटेंशन | 68 डिग्री सेल्सियस | 2 मिनट | ||
| प्राइमर C4R | 10 माइक्रोन | 0.2 माइक्रोल | डेन्चरिंग | 94 डिग्री सेल्सियस | 10 एस | 25 | |
| 250 यू टाक डीएनए पॉलीमरेज | 10 यू | 0.13 माइक्रोल | एनीलिंग | 58 डिग्री सेल्सियस | 2 मिनट | ||
| एच2ओ | 15.5 माइक्रोल | एक्सटेंशन* | 68 डिग्री सेल्सियस | 2 मिनट | |||
| अंतिम विस्तार | 72 डिग्री सेल्सियस | 5 मिनट | 1 | ||||
| 4 डिग्री सेल्सियस | पकड़ | ||||||
| * 10 एस के साथ 68% सी विस्तार कदम के लिए प्रत्येक चक्र जोड़ा। | |||||||
| घटक | अंतिम एकाग्रता | 25 माइक्रोल प्रतिक्रिया | साइकिल चरण | तापमान | समय | चक्र | |
| 10x पीसीआर बफर | 10x | 2.5 माइक्रोन | प्रारंभिक विकृति | 94 डिग्री सेल्सियस | 2 मिनट | 1 | |
| 5x क्यू-सॉल्यूशन | 5x | 5 माइक्रोन | डेन्चरिंग | 94 डिग्री सेल्सियस | 30 एस | 35 | |
| 100 एमएमएम डीएनटीपी | 2.5 mM | 0.5 माइक्रोन | एनीलिंग | 62 डिग्री सेल्सियस | 1.5 मिनट | ||
| प्राइमर SHLM1 | 10 माइक्रोन | 0.2 माइक्रोल | एक्सटेंशन | 72 डिग्री सेल्सियस | 2 मिनट | ||
| प्राइमर SHLM2 | 10 माइक्रोन | 0.2 माइक्रोल | अंतिम विस्तार | 72 डिग्री सेल्सियस | 5 मिनट | 1 | |
| 250 यू टाक डीएनए पॉलीमरेज | 10 यू | 0.13 माइक्रोल | 4 डिग्री सेल्सियस | पकड़ | |||
| एच2ओ | 15.5 माइक्रोल | ||||||
| घटक | अंतिम एकाग्रता | 20 माइक्रोल प्रतिक्रिया | साइकिल चरण | तापमान | समय | चक्र | |
| 5x पीसीआर बफर | 5x | 4 माइक्रोन | प्रारंभिक विकृति | 98 डिग्री सेल्सियस | 30 एस | 1 | |
| डीएनटीपी | 2.5 mM | 1.6 माइक्रोल | डेन्चरिंग | 98 डिग्री सेल्सियस | 10 एस | 35 | |
| प्राइमर SHLM3 | 10 माइक्रोन | 1 माइक्रोल | एनीलिंग | 51 डिग्री सेल्सियस | 30 एस | ||
| प्राइमर SHLM4 | 10 माइक्रोन | 1 माइक्रोल | एक्सटेंशन | 72 डिग्री सेल्सियस | 1 मिनट | ||
| 100 यू डीएनए पॉलीमरेज | 5 यू | 0.2 माइक्रोल | अंतिम विस्तार | 72 डिग्री सेल्सियस | 5 मिनट | 1 | |
| एच2ओ | 12.4 माइक्रोन | 4 डिग्री सेल्सियस | पकड़ | ||||
| घटक | अंतिम एकाग्रता | 25 माइक्रोल प्रतिक्रिया | साइकिल चरण | तापमान | समय | चक्र | |
| 10x पीसीआर बफर | 10x | 2.5 माइक्रोन | प्रारंभिक विकृति | 94 डिग्री सेल्सियस | 2 मिनट | 1 | |
| 100 एमएमएम डीएनटीपी | 2.5 mM | 2 माइक्रोन | डेन्चरिंग | 94 डिग्री सेल्सियस | 30 एस | 35 | |
| प्राइमर FBC8-F | 10 माइक्रोन | 1 माइक्रोल | एनीलिंग | 55 डिग्री सेल्सियस | 1.5 मिनट | ||
| प्राइमर FBC8-R | 10 माइक्रोन | 1 माइक्रोल | एक्सटेंशन | 72 डिग्री सेल्सियस | 2 मिनट | ||
| 250 यू टाक डीएनए पॉलीमरेज | 10 यू | 0.125 माइक्रोल | अंतिम विस्तार | 72 डिग्री सेल्सियस | 5 मिनट | 1 | |
| एच2ओ | 17.4 माइक्रोन | 4 डिग्री सेल्सियस | पकड़ | ||||
तालिका 1. इस प्रोटोकॉल में उपयोग किए जाने वाले विभिन्न प्राइमर संयोजनों के लिए प्रतिक्रिया और साइकिलिंग की स्थिति।
| प्राइमरों | अनुक्रम | संदर्भ |
| एसआरसी-एफ | 5'-सीटीटीसीसीटीटीटीटीटीटीटीटीटीटीसीटीटीसीटीजीसी-3' | 18 |
| एसआरसी-आर | 5'-GGCCATTTTTTGCCCCTTG-3' | 18 |
| प्रू-सी 2 | 5'-CTTTGGCAGTAATTAATTAAACC-3 ' | 35 |
| प्रू-सी4आर | 5'-GGATGTGGTACGATGATGCG-3' | 35 |
| SHLM1-F | 5'-GGTGGGATGATGATTAGTAGCCCC-3' | 17 |
| SHLM2-R | 5'-GGCTGCATAAGAGCTGTAGG-3' | 17 |
| SHLM3-F | 5'-टाटाटीसीटीटीटीटीटीजीसी-3' | 17 |
| SHLM4-R | 5'-CACTATGATAATTTTATG-3' | 17 |
| AprFBC8-F | 5'-CATGGAAAAAAAAAACTGACTTATGG-3' | 26 |
| AprFBC8-R | 5'-GCCTCTAATTTCATCATCTCTCTTAG-3' | 26 |
तालिका 2. प्रूनस आर्मेनियाकामें एस-जीनोटाइपलक्षण वर्णन के लिए इस प्रोटोकॉल, अनुक्रम और संदर्भ में उपयोग किए जाने वाले प्राइमर।
| Cultivar | एसआरसी-(F/R) (बीपी) | PruC2/PruC4R (बीपी) | SHLM1/SHLM2 (बीपी) | SHLM3/SHLM4 (बीपी) | AprFBC8-(F/R) (बीपी) | एस-जीनोटाइप | असंगति समूह (I.G) |
| वंडर कॉट8 | 420, 420 | 749, 1386 | एस6एस9 | आठवीं | |||
| जादू खाट8 | 334, 420 | 749 | एस2एस9 | Xx | |||
| गोल्डस्ट्राइक8 | 334, 420 | 749 | एस2एस9 | - | |||
| टी06917 | 334, 408 | 650 | एस1एस2 | मैं | |||
| टी12017 | 334, 408 | 650 | एस1एस2 | - | |||
| सी-6 | 334, 408 | 413 | एस2एस7 | Iv | |||
| कूपर खाट8 | 274, 408 | 650 | एस1एस3 | XVIII | |||
| एपिकेन | 358, 358 | 500 | एससीएससी | - | |||
| बर्गकोट 8 | 334, 358 | 500 | एस2एससी | - | |||
| स्प्रिंगब्लश 8 | 274, 358 | 150 | एस3एस8 | Xxi |
तालिका 3. इस प्रोटोकॉल और असंगति समूह असाइनमेंट में उपयोग किए जाने वाले पांच प्राइमर जोड़े के साथ खुबानी खेती की एस-जेनोटीपिंग। S एसआरसी-(एफ/आर), PruC2/PruC4R, SHLM1/SHLM2, SHLM3/SHLM4, और AprFBC8-(F/R) प्राइमर का उपयोग करके परिलक्षित एस-एलील्सके विभिन्न पॉलीमरेज चेन रिएक्शन उत्पाद आकार तालिका में दिखाए गए हैं ।
Subscription Required. Please recommend JoVE to your librarian.
Discussion
परंपरागत रूप से, अधिकांश वाणिज्यिक खुबानी यूरोपीय खेती स्वयं संगत३६थे । फिर भी, पिछले दशकों में प्रजनन कार्यक्रमों में माता-पिता के रूप में उत्तरी अमेरिकी आत्म-असंगत खेती के उपयोग के परिणामस्वरूप अज्ञात परागण आवश्यकताओं7,,8,,37के साथ नई आत्म-असंगत खेती की बढ़ती संख्या जारी हुई है। इस प्रकार, खुबानी खेती में आत्म और अंतर-अनुकूलता संबंधों का निर्धारण तेजी से महत्वपूर्ण है । यह उन क्षेत्रों में दबाव डाला जाता है जहां सर्दियों में ठंड कम हो रही है, फूलों के समय में उच्च वर्ष से वर्ष की विविधताएं कई मामलों में खेती और उनके परागकणों के फूलों में संयोग को रोक रही हैं, विशेष रूप से उच्च डरावना आवश्यकताओं के साथ खेती में38। यहां वर्णित पद्धति, हाथ परागण, माइक्रोस्कोपी और आनुवंशिक विश्लेषण के संयोजन से प्रत्येक खेती की आत्म (में) अनुकूलता का निर्धारण करने और इसकी संभावित परागणक खेती स्थापित करने के लिए बहुत उपयोगी रहा है ।
परागण आवश्यकताओं को बाग की स्थितियों में क्षेत्र-नियंत्रण प्रयोगों के माध्यम से निर्धारित किया जा सकता है11,,39. हालांकि, मौसम संबंधी प्रतिकूल परिस्थितियों सहित बाहरी कारकों के लिए प्रदर्शनी परागण विफलता10का कारण बन सकती है, जिसके परिणामस्वरूप आत्म-असंगति का गलत निदान हो सकता है। यहां वर्णित पद्धति पर्यावरणीय प्रभाव से बचने, प्रयोगशाला नियंत्रित स्थितियों में हाथ से परागणित फूलों की सूक्ष्मकॉपी टिप्पणियों द्वारा आत्म-(इन) अनुकूलता का मूल्यांकन करने की अनुमति देती है। इसके अलावा, यह दृष्टिकोण प्रति वर्ष अधिक संख्या में खेती का विश्लेषण करने की अनुमति देता है, क्योंकि क्षेत्र प्रयोगों में आवश्यक प्रत्येक खेती के लिए कई वयस्क पेड़ों के बजाय केवल थोड़ी संख्या में फूलों की आवश्यकता होती है40।
असंगति संबंधों को हाथ परागण और माइक्रोस्कोपी14के संयोजन से स्थापित किया जा सकता है । हालांकि, परागण केवल वसंत ऋतु में फूलों के मौसम के दौरान एक छोटी अवधि के लिए किया जा सकता है, और प्रयोगशाला के पास वयस्क पेड़ों की आवश्यकता होती है, क्योंकि एकत्र किए गए फूलों की उम्र बहुत कम होती है। इस प्रकार, असंगति संबंधों की संख्या है कि प्रत्येक मौसम में नियंत्रित हाथ परागण द्वारा विश्लेषण किया जा सकता है बहुत कम है । एस-लोकस द्वारा एन्कोड Sकिए गए जीन के लक्षण वर्णन ने एस-एलीलजीनोटाइपिंग18,,41के लिए पीसीआर आधारित विधियों के विकास को सक्षम बनाया है। यह दृष्टिकोण एस-एलीलपहचान को तेज करता है क्योंकि इसमें फूलों की आवश्यकता नहीं होती है, और प्रयोग किसी भी वनस्पति ऊतक42के साथ किए जा सकते हैं। यह उस अवधि को बढ़ाता है जिसके दौरान पौधे की सामग्री, आमतौर पर युवा पत्तियों को43एकत्र किया जा सकता है। इसके अलावा, पत्तियों को ल्योफिलाइज्ड या फ्रोजन किया जा सकता है, ताकि विश्लेषण वर्ष के किसी भी समय किया जा सके, परागण के विपरीत जो केवल फूलों के मौसम44के दौरान ताजे फूलों पर किया जा सकता है। एक अतिरिक्त लाभ यह है कि फूलों की उम्र में प्रवेश करने से पहले भी युवा पेड़ों से पत्तियों को एकत्र किया जा सकता है, नमूनों के संग्रह को सुविधाजनक बनाता है और परिणाम45के प्रारंभिक प्राप्त होते हैं।
आनुवंशिक विश्लेषण आत्म-असंगति एलील्स के बेहतर भेदभाव की अनुमति देता है क्योंकि यह21,,46के मिश्रित टुकड़े के आकार के सटीक परिणाम प्रदान करता है। आज,तक, खुबानी22,,,,12, 18, 20, 21, 22,,,1823,,2324,में तैंतीस एस-एलील्सकी पहचान की गई है,24जिसने एस-जीनोटाइप8,9,17,25, 26, 27पर आधारित 36 असंगति समूह स्थापित करनेकीअनुमति दी है।,,27 दूसरी ओर, इस पद्धति की एक खामी यह है कि एक ही सीमा आकार या उत्परिवर्तनों में विभिन्न एलील्स को गलत तरीके से एक ही एलील के रूप में पहचाना जा सकता है। इस प्रकार, आरएनएसई अनुक्रम के लिए एससी और एस8 एलील्स समान हैं लेकिन अनुसूचित जाति19के एसएफबी जीन में 358-बीपी सम्मिलन पाया गया है। इसी तरह, एलील्स एस1 और एस7 का पहला इंट्रॉन क्षेत्र समान है और प्राइमर एसआरसी-एफ/एसआरसी-आर का उपयोग करके अविवेच्य हैं ।7 इसके अलावा, एस6 और एस52 88 या एस20 और एस55,और एस 7,20 एस13 (EF062341) और एस46 17जैसे कई समरूपताएं पाई गई हैं क्योंकि इनमें से कुछ एलील्स को पीसीआर प्रवर्धन केदौरानआंशिक रूप से अनुक्रमित या विफलताओं से किया गया है और परिणामस्वरूप, उन्हें सही ढंग से प्रतिष्ठित करने के लिए आगे के कार्य की आवश्यकता है।7 S
एस-एलील्स की पहचान और उनके संबंधित असंगति समूह,,8,17,26,27में कृषि के आवंटन के माध्यम से असंगति संबंध स्थापित करने के लिए पीसीआर विश्लेषण और, एस-आरएनएएस अनुक्रमण पर्याप्त हैं । S हालांकि, इस पद्धति में विशेष खुबानी खेती के लिए आत्म-(इन) अनुकूलता के निर्धारण को रोकने की सीमा है। स्व-अनुकूलता (एससी) अन्य प्रूनस प्रजातियों47,बादाम (एसएफ)48, 49,या मीठी चेरी(एस4')’50, 51,51के रूप में विशेषएस-एलील्ससे जुड़ी हुई है।49 हालांकि, खुबानी में, अनुसूचित जाति एलील, जो अनुसूचित जाति21से जुड़ा हुआ है, गलती से एस8,एक आत्म-असंगत एलील19,22,और एस लोकस से जुड़े नहीं संभावित म्यूटेशन के रूप में पहचाना जा सकता है, क्योंकि एम-टिड्डी12,52, अनुसूचित जाति प्रदान करने वाले एम-टिड्डी12, 52की पहचान की गई है।,, हाल ही Mमें, एम-लोकस को एसएसआर मार्कर12का उपयोग करके जीनोटाइप किया गया है। इसलिए, खुबानी जीनोटाइप के लिए अनुसूचित जाति की आनुवंशिक पहचान को आगे अनुसंधान की आवश्यकता है और, एस लोकस से जुड़े कारकों के कारण गलतियों से बचने के लिए, इस काम में स्वयं का लक्षण वर्णन (में) अनुकूलता भी आत्म परागणित फूलों के पिस्टिल के माध्यम से पराग ट्यूबों के व्यवहार को फेनोटाइपिंग द्वारा निर्धारित किया गया है ।
फ्लोरोसेंस माइक्रोस्कोपी के तहत नियंत्रित आत्म-परागण के पिस्टिल में परागण के व्यवहार के बाद के अवलोकन के साथ प्रयोगशाला की स्थितियों में हाथ से परागण द्वारा आत्म-परागण के Sनिर्धारण के संयोजन में वर्णित पद्धति और पीसीआर विश्लेषण द्वारा एस-जीनोटाइप की पहचान खुबानी खेती की परागण आवश्यकताओं को स्थापित करने की अनुमति देती है। यह उत्पादकों और प्रजनकों के लिए एक मूल्यवान जानकारी प्रदान करता है, क्योंकि यह खेती के बीच असंगति संबंधों को स्थापित करने के लिए नए बगीचों के डिजाइन में उपयुक्त परागणकारों का चयन करने के साथ ही उचित माता पिता का चयन करने के लिए खुबानी प्रजनन कार्यक्रमों में नए पार डिजाइन की अनुमति देता है ।
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
लेखकों के पास खुलासा करने के लिए कुछ नहीं है ।
Acknowledgments
इस शोध को मिनिस्टरियो डी सिन्सिया, इनोवासिओन वाई यूनीवर्सिडेस-यूरोपियन रीजनल डेवलपमेंट फंड, यूरोपियन यूनियन (AGL2016-77267-R, और AGL2015-74071-JIN द्वारा वित्त पोषित किया गया था); इंस्टीट्यूटो नैसिनल डी इन्वेस्टिगासिओन वाई टेक्नोलोगिया एगरिया वाई एलिमेंटेरिया (RFP2015-00015-00, RTA2017-00003-00); गोबिएर्नो डी अरगोन-यूरोपियन सोशल फंड, यूरोपियन यूनियन (ग्रुपो कंसोलिडाडो A12_17R), फंडासिओन बायोडाइवर्सिड, और एग्रोसेगुरो एसईए
Materials
| Name | Company | Catalog Number | Comments |
| Agarose D1 Low EEO | Conda | 8010.22 | |
| BIOTAQ DNA Polymerase kit | Bioline | BIO-21060 | |
| Bright field microscope | Leica Microsystems | DM2500 | |
| CEQ System Software | Beckman Coulter | ||
| DNeasy Plant Mini Kit | QIAGEN | 69106 | |
| dNTP Set, 4 x 25 µmol | Bioline | BIO-39025 | |
| GenomeLab DNA Size Standard Kit - 400 | Beckman Coulter | 608098 | |
| GenomeLab GeXP Genetic Analysis System | Beckman Coulter | ||
| GenomeLab Separation Buffer | Beckman Coulter | 608012 | |
| GenomeLab Separation Gel LPA-1 | Beckman Coulter | 391438 | |
| HyperLadder 100bp | Bioline | BIO-33029 | |
| HyperLadder 1kb | Bioline | BIO-33025 | |
| Image Analysis System | Leica Microsystems | ||
| Molecular Imager VersaDoc MP 4000 system | Bio-Rad | 170-8640 | |
| NanoDrop One Spectrophotometer | Thermo Fisher Scientific | 13-400-518 | |
| pH-Meter BASIC 20 | Crison | ||
| Phusion High-Fidelity PCR Kit | Thermo Fisher Scientific | F553S | |
| Power Pack P 25 T | Biometra | ||
| Primer Forward | Isogen Life Science | ||
| Primer Reverse | Isogen Life Science | ||
| Quantity One Software | Bio-Rad | ||
| Stereoscopic microscope | Leica Microsystems | MZ-16 | |
| Sub-Cell GT | Bio-Rad | ||
| SYBR Safe DNA Gel Stain | Thermo Fisher Scientific | S33102 | |
| T100 Thermal Cycler | Bio-Rad | 1861096 | |
| Taq DNA Polymerase | QIAGEN | 201203 | |
| Vertical Stand Autoclave | JP Selecta |
References
- Silva, N. F., Goring, D. R.
- Charlesworth, D., Vekemans, X., Castric, V., Glémin, S. Plant self-incompatibility systems: A molecular evolutionary perspective. New phytologist. 168, 61-69 (2005).
- Tao, R., et al. Identification of stylar RNases associated with gametophytic self-incompatibility in almond (Prunus dulcis). Plant and Cell Physiology. 38, 304-311 (1997).
- Ushijima, K., et al. Structural and transcriptional analysis of the self-incompatibility locus of almond: Identification of a pollen-expressed F-box gene with haplotype-specific polymorphism. The Plant cell. 15, 771-781 (2003).
- Bedinger, P. A., Broz, A. K., Tovar-Mendez, A., McClure, B. Pollen-Pistil Interactions and Their Role in Mate Selection. Plant Physiology. 173, 79-90 (2017).
- Guerra, M. E., Rodrigo, J. Japanese plum pollination: A review. Scientia Horticulturae. 197, 674-686 (2015).
- Zhebentyayeva, T., Ledbetter, C., Burgos, L., Llacer, G. Apricot. Fruit Breeding. Badenes, M. L., Byrne, D. , Springer. Boston, MA. 415-458 (2012).
- Herrera, S., Lora, J., Hormaza, J. I., Herrero, M., Rodrigo, J. Optimizing Production in the New Generation of Apricot Cultivars: Self-incompatibility, S-RNase Allele Identification, and Incompatibility Group Assignment. Frontiers in Plant Science. 9, 527 (2018).
- Egea, J., Burgos, L. Detecting Cross-incompatibility of Three North American Apricot Cultivars and Establishing the First Incompatibility Group in Apricot. Journal of the American Society for Horticultural Science. 121, 1002-1005 (1996).
- Rodrigo, J., Herrero, M. Effects of pre-blossom temperatures on flower development and fruit set in apricot. Scientia Horticulturae. 92, 125-135 (2002).
- Julian, C., Herrero, M., Rodrigo, J. Flower bud differentiation and development in fruiting and non-fruiting shoots in relation to fruit set in apricot (Prunus armeniaca). Trees. 24, 833-841 (2010).
- Muñoz-Sanz, J. V., Zuriaga, E., López, I., Badenes, M. L., Romero, C. Self-(in)compatibility in apricot germplasm is controlled by two major loci, S and M. BMC Plant Biology. 17, 82 (2017).
- Burgos, L., Berenguer, T., Egea, J. Self- and Cross-compatibility among Apricot Cultivars. HortScience. 28, 148-150 (1993).
- Rodrigo, J., Herrero, M. Evaluation of pollination as the cause of erratic fruit set in apricot "Moniqui". Journal of Horticultural Science. 71, 801-805 (1996).
- Milatović, D., Nikolić, D., Krška, B. Testing of self-(in)compatibility in apricot cultivars from European breeding programmes. Horticultural Science. 40 (2), 65-71 (2013).
- Milatović, D., Nikolić, D., Fotirić-Aksić, M., Radović, A. Testing of self-(in)compatibility in apricot cultivars using fluorescence microscopy. Acta Scientiarum Polonorum, Hortorum Cultus. 12 (6), 103-113 (2013).
- Herrera, S., Rodrigo, J., Hormaza, J. I., Lora, J. Identification of Self-Incompatibility Alleles by Specific PCR Analysis and S-RNase Sequencing in Apricot. Int J Mol Sci. 19, 3612 (2018).
- Romero, C., et al. Analysis of the S-locus structure in Prunus armeniaca L. Identification of S-haplotype specific S-RNase and F-box genes. Plant Molecular Biology. 56, 145-157 (2004).
- Halász, J., Pedryc, A., Hegedus, A. Origin and dissemination of the pollen-part mutated SC haplotype which confers self-compatibility in apricot (Prunus armeniaca). New Phytologist. 176, 792-803 (2007).
- Halász, J., Hegedus, A., Hermán, R., Stefanovits-Bányai, É, Pedryc, A. New self-incompatibility alleles in apricot (Prunus armeniaca L.) revealed by stylar ribonuclease assay and S-PCR analysis. Euphytica. 145, 57-66 (2005).
- Vilanova, S., Romero, C., Llacer, G., Badenes, M. L., Burgos, L. Identification of Self-(in)compatibility Alleles in Apricot by PCR and Sequence Analysis. Journal of the American Society for Horticultural Science. 130, 893-898 (2005).
- Feng, J., et al. Detection and transcript expression of S-RNase gene associated with self-incompatibility in apricot (Prunus armeniaca L.). Molecular Biology Reports. 33, 215-221 (2006).
- Zhang, L., et al. Identification of self-incompatibility (S-) genotypes of Chinese apricot cultivars. Euphytica. 160, 241-248 (2008).
- Wu, J., et al. Identification of S-haplotype-specific S-RNase and SFB alleles in native Chinese apricot (Prunus armeniaca L). Journal of Horticultural Science and Biotechnology. 84, 645-652 (2009).
- Szabó, Z., Nyéki, J. Blossoming, fructification and combination of apricot varieties. Acta Horticulturae. 293, 295-302 (1991).
- Halász, J., Pedryc, A., Ercisli, S., Yilmaz, K. U., Hegedűs, A. S-genotyping supports the genetic relationships between Turkish and Hungarian apricot germplasm. Journal of the American Society for Horticultural Science. 135, 410-417 (2010).
- Lachkar, A., et al. Identification of self-(in)compatibility S-alleles and new cross-incompatibility groups in Tunisian apricot (Prunus armeniaca L.) cultivars. The Journal of Horticultural Science and Biotechnology. 88, 497-501 (2013).
- Pérez-Pastor, A., Ruiz-Sánchez, M. C., Domingo, R., Torrecillas, A. Growth and phenological stages of Búlida apricot trees in South-East. Agronomie. 24, 93-100 (2004).
- Williams, J. H., Friedman, W. E., Arnold, M. L. Developmental selection within the angiosperm style: using gamete DNA to visualize interspecific pollen competition. Proceedings of the National Academy of Sciences of the United States of America. 96, 9201-9206 (1999).
- Julian, C., Herrero, M., Rodrigo, J. Anther meiosis time is related to winter cold temperatures in apricot (Prunus armeniaca L.). Environmental and Experimental Botany. 100, 20-25 (2014).
- Guerra, M. E., López-Corrales, M., Wünsch, A., Rodrigo, J. Lack of Fruit Set Caused by Ovule Degeneration in Japanese Plum. Journal of the American Society for Horticultural Science. 136 (6), 375-381 (2011).
- Guerra, M. E., Wünsch, A., López-Corrales, M., Rodrigo, J. Flower Emasculation as the Cause for Lack of Fruit Set in Japanese Plum Crosses. Journal of the American Society for Horticultural Science. 135 (6), 556-562 (2010).
- Hormaza, J. I., Pinney, K., Polito, V. S. Correlation in the tolerance to ozone between sporophytes and male gametophytes of several fruit and nut tree species (Rosaceae). Sexual Plant Reproduction. 9, 44-48 (1996).
- Alcaraz, M. L., Hormaza, J. I., Rodrigo, J. Pistil Starch Reserves at Anthesis Correlate with Final Flower Fate in Avocado (Persea americana). PLOS ONE. 8 (10), 78467 (2013).
- Tao, R., et al. Molecular typing of S-alleles through Identification, Characterization and cDNA cloning for S-RNases in Sweet Cherry. Journal of the American Society for Horticultural Science. 124, 224-233 (1999).
- Burgos, L., et al. The self-compatibility trait of the main apricot cultivars and new selections from breeding programmes. Journal of Horticultural Science. 72, 147-154 (1997).
- Hormaza, J. I., Yamane, H., Rodrigo, J. Apricot. Genome Mapping and Molecular Breeding in Plants, Volume 4 Fruits and Nuts. Kole, C. , Springer. New York, NY. 171-187 (2007).
- Benmoussa, H., Ghrab, M., Ben Mimoun, M., Luedeling, E. Chilling and heat requirements for local and foreign almond (Prunus dulcis Mill.) cultivars in a warm Mediterranean location based on 30 years of phenology records. Agricultural and Forest Meteorology. 239, 34-46 (2017).
- Rodrigo, J., Herrero, M., Hormaza, J. I. Pistil traits and flower fate in apricot (Prunus armeniaca). Annals of Applied Biology. 154, 365-375 (2009).
- Williams, R. R. Techniques used in fruit-set experiments. Towards Regulated Cropping. Williams, R. R., Wilson, D. , Grower Books. London. 57-61 (1970).
- Sutherland, B. G., Robbins, T. P., Tobutt, K. R. Primers amplifying a range of Prunus S-alleles. Plant Breeding. 123, 582-584 (2004).
- Murray, M. G., Thompson, W. F. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Research. 8, 4321-4325 (1980).
- Porebski, S., Bailey, L. G., Baum, B. R. Modification of a CTAB DNA Extraction Protocol for Plants Containing High Polysaccharide and Polyphenol Components. Plant Molecular Biology Reporter. 15 (1), 8-15 (1997).
- Rogers, S. O., Bendich, A. J. Extraction of DNA from milligram amounts of fresh, herbarium and mummified plant tissues. Plant Molecular Biology. 5 (2), 69-76 (1985).
- Hormaza, J. I. Molecular characterization and similarity relationships among apricot (Prunus armeniaca L.) genotypes using simple sequence repeats. Theoretical and Applied Genetics. 104, 321-328 (2002).
- Sonneveld, T., Tobutt, K. R., Robbins, T. P. Allele-specific PCR detection of sweet cherry self-incompatibility (S) alleles S1 to S16 using consensus and allele-specific primers. Theoretical and Applied Genetics. 107, 1059-1070 (2003).
- Hegedus, A., Lénárt, J., Halász, J. Sexual incompatibility in Rosaceae fruit tree species: molecular interactions and evolutionary dynamics. Biologia Plantarum. 56 (2), 201-209 (2012).
- Fernández i Martí, A., Gradziel, T. M., Socias i Company, R. Methylation of the Sf locus in almond is associated with S-RNase loss of function. Plant Molecular Biology. 86, 681-689 (2014).
- Company, R. S. i, Kodad, O., Martí, A. F. i, Alonso, J. M. Mutations conferring self-compatibility in Prunus species: From deletions and insertions to epigenetic alterations. Scientia Horticulturae. 192, 125-131 (2015).
- Boskovic, R., Tobutt, K. R. Correlation of stylar ribonuclease zymograms with incompatibility alleles in sweet cherry. Euphytica. 90, 245-250 (1996).
- Cachi, A. M., Wünsch, A. S-genotyping of sweet cherry varieties from Spain and S-locus diversity in Europe. Euphytica. 197 (2), 229-236 (2014).
- Zuriaga, E., et al. An S-locus Independent Pollen Factor Confers Self-Compatibility in "Katy" Apricot. PLoS ONE. 8 (1), 53947 (2013).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}