

Summary
We presenteren een methodologie om de bestuivingsvereisten van abrikoos(Prunus armeniëca L.) cultivars vast te stellen die de bepaling van zelf-(in)compatibiliteit door fluorescentiemicroscopie combineren met de identificatie van het S-genotype door PCR-analyse.
Abstract
Zelf-incompatibiliteit in Rosaceae wordt bepaald door een Gametophytic Self-Incompatibility System (GSI) dat voornamelijk wordt gecontroleerd door de multiallelic locus S. In abrikoos wordt de bepaling van zelf- en inter-(in)compatibiliteitsrelaties steeds belangrijker, omdat het vrijkomen van een belangrijk aantal nieuwe cultivars heeft geleid tot de toename van cultivars met onbekende bestuivingsvereisten. Hier beschrijven we een methodologie die de bepaling van zelf-(in)compatibiliteit met handbestuivingen en microscopie combineert met de identificatie van het S-genotypedoor PCR-analyse. Voor zelf-(in)compatibiliteitsbepaling werden bloemen in het ballonstadium van elke cultivar verzameld in het veld, met de hand bestoven in het laboratorium, vast, en gekleurd met anilineblauw voor de observatie van pollenbuisgedrag onder de fluorescentiemicroscopie. Voor het tot stand brengen van onverenigbaarheidsrelaties tussen cultivars werd DNA van elke cultivar uit jonge bladeren gehaald en werden S-allelesgeïdentificeerd door PCR. Deze aanpak maakt het mogelijk om incompatibiliteitsgroepen vast te stellen en incompatibiliteitsrelaties tussen cultivars op te helderen, wat een waardevolle informatie biedt om geschikte bestuivers te kiezen bij het ontwerpen van nieuwe boomgaarden en om geschikte ouders in fokprogramma's te selecteren.
Introduction
Zelf-incompatibiliteit is een strategie van bloeiende planten om zelfbestuiving te voorkomen en outcrossing te bevorderen1. In Rosaceae wordt dit mechanisme bepaald door een Gametophytic Self-Incompatibility System (GSI) dat voornamelijk wordt gecontroleerd door de multiallelic locus S2. In de stijl codeert het RNase-gen de S-stylar determinant, een RNase3, terwijl een F-box eiwit, dat de S-pollendeterminant bepaalt, wordt gecodificeerd door het SFB-gen 4. De zelf-onverenigbaarheid interactie vindt plaats door de remming van pollen buis groei langs de stijl het voorkomen van de bevruchting van de ovule5,6.
In abrikoos, een ras vernieuwing heeft plaatsgevonden wereldwijd in de afgelopen twee decennia7,8. Deze introductie van een belangrijk aantal nieuwe cultivars, uit verschillende publieke en private veredelingsprogramma's, heeft geresulteerd in de toename van abrikozencultivanten met onbekende bestuivingsvereisten8.
Verschillende methoden zijn gebruikt om de bestuivingsvereisten in abrikoos te bepalen. In het veld kan zelf-(in)compatibiliteit worden vastgesteld door gecontroleerde bestuivingen in gekooide bomen of in ontlaste bloemen en vervolgens het percentage fruitset9,10,,11,12registreren . Bovendien zijn in het laboratorium gecontroleerde bestuivingen uitgevoerd door een semi-in vivo bloementeelt en analyse van het gedrag van de pollenbuis onder fluorescentiemicroscopie8,13,14,15,16,17. Onlangs hebben moleculaire technieken, zoals PCR-analyse en sequencing, de karakterisering van onverenigbaarheidsrelaties mogelijk gemaakt op basis van de studie van de RNase- en SFB-genen 18,19. In abrikoos zijn drieëndertig S-allelen gemeld (S1 tot S20, S22 tot S30, S52, S53, Sv, Sx), inclusief één allel in verband met zelfcompatibiliteit (Sc)12,18,20,21,22,23,24. Tot nu toe zijn 26 onverenigbaarheidsgroepen bij deze soort ge stablished volgens het S-genotype S8,9,,17,25,26,27. Cultivars met Sdezelfde S-allelen zijn onderling onverenigbaar, terwijl cultivars met ten minste één verschillend S-allelen bijgevolg in verschillende onverenigbare groepen intercompatibel zijn.
Om de bestuivingsvereisten van abrikozencultivivars te definiëren, beschrijven we een methodologie die de bepaling van zelf-(in)compatibiliteit door fluorescentiemicroscopie combineert met de identificatie van het S-genotype door PCR-analyse in abrikozencultivivars. Deze aanpak maakt het mogelijk om incompatibiliteitsgroepen vast te stellen en incompatibiliteitsrelaties tussen cultivars op te helderen.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. Zelf-(in)compatibiliteitsbepaling
- Proef de bloemen in het veld. Het is noodzakelijk om de bloemen te verzamelen op ballonstadium(figuur 1A), overeenkomend met fase 58 op de BBCH-schaal voor abrikoos28, om ongewenste eerdere bestuiving te voorkomen.
-
Zelf- en kruisbestuivingen in het laboratorium
- Verwijder de anthers van de bloemen op ballon stadium en leg ze op een stuk papier te drogen op laboratoriumtemperatuur.
- Na 24 uur zeef je de stuifmeelkorrels met behulp van een fijn gaas (0,26 mm) (figuur 1B).
- Emasculate een groep van 30 bloemen op dezelfde ballon stadium voor elke zelfbestuiving en kruisbestuiving en plaats de stampers op bloemist schuim in water op laboratoriumtemperatuur (Figuur 1C).
- Hand bestuiven de stampers met behulp van een penseel met stuifmeel van bloemen van dezelfde cultivar 24 uur na emasculatie. Bovendien bestuiven een andere set van stampers van elke cultivar met stuifmeel van bloemen van een compatibele bestuiver als controle(Figuur 1D).
- Na 72 uur bevestigt u de stampers in een fixatieve oplossing van ethanol/azijnzuur (3:1) voor ten minste 24 uur bij 4 °C29. Gooi vervolgens de fixatieve en voeg 75% ethanol ervoor te zorgen dat de monsters volledig worden ondergedompeld in de oplossing. Monsters kunnen in deze oplossing bij 4 °C worden bewaard tot gebruik8,17,30,31,32.
-
Evaluatie van de levensvatbaarheid van stuifmeel door in vitro pollen kiemkracht
- Ter voorbereiding van het kiemmiddel gewicht 25 g sacharose, 0,075 g boorzuur (H3BO3) en 0,075 g calciumnitraat (Ca(NR3)2)33.
- Voeg de componenten van het medium toe in 250 mL gedestilleerd water en los volledig op.
- Stollen het medium toe te voegen 2 g agarose en meng door werveling.
- Controleer de pH van het medium met behulp van een pH-meter en pas de waarde aan op 7.0 met NaOH- of HCl-oplossing.
- Autoclave het mengsel om het medium te steriliseren.
- Na autoclaving koel je het medium af en verdeel je het in petrischaaltjes in een steriele laminaire flow hood.
- Verstrooi de stuifmeelkorrels van dezelfde cultivars die worden gebruikt voor de gecontroleerde bestuivingen in het gestolde pollenkiemingsmedium en observeer ze onder de microscoop na 24 uur6.
OPMERKING: Om de laminaire stroomkap te steriliseren, reinigt u het oppervlak met 70% ethanol en schakelt u de UV-lamp gedurende 10 minuten in. - Bewaar de petrischaaltjes in een koelkast op 4 °C tot gebruik.
-
Microscopie-waarnemingen
- Was de stampers drie maal gedurende 1 uur met gedestilleerd water en laat ze in 5% natriumsulfiet bij 4 °C. Na 24 uur, autoclave ze op 1 kg/cm2 gedurende 10 min in natriumsulfiet om de weefsels te verzachten34.
- Plaats de autoclaved pistils over een glazen glijbaan en verwijder, met behulp van een scalpel, de trichomen rond de eierstok om een betere visualisatie van de pollenbuizen te krijgen. Dan, squash de stampers met een cover glas.
- Bereid 0,1% (v/v) aniline blauwe vlek: meng 0,1 mL aniline blauw in 100 mL van 0,1 N kalium fosfaat tribasic (K3PO4). Breng een druppel anilineblauw aan over de preparaten om calloseafzettingen te bevlekken tijdens de groei van de pollenbuis.
- Observeer de stuifmeelbuizen langs de stijl door een microscoop met UV-epifluorescentie met behulp van 340-380 bandpass en 425 longpass filters.
2. DNA-extractie
- Monster 2-3 bladeren in het veld in het voorjaar. Het wordt aanbevolen om de bladeren in jonge stadia te bemonsteren, omdat dna verkregen van hogere kwaliteit en lagere niveaus van fenolische verbindingen is in vergelijking met oude bladeren.
- Haal Genomic DNA naar aanleiding van de stappen beschreven in een commercieel beschikbare kit (zie Tabel van materialen).
- Analyseer de kwantiteit en kwaliteit van de DNA-concentraties met uv-visspectrfotometer (260 nm).
3. SS-allel identificatie
-
Opzetten van de PCR-reacties
- Bereid een verdunning van 50 ng/μL in gedestilleerd water van elk DNA-extractiemonster.
- Ontdooi de PCR-reagentia langzaam en houd ze op ijs. Laat het DNA polymerase in de vriezer tot het nodig is.
- Bereid de versterkingsreacties voor met behulp van de verschillende combinaties van primers. Maak de PCR-reactiemix door de componenten in tabel 1te combineren. Vortex de PCR reactie meng goed en verdeel het volume aangegeven voor de verschillende combinaties van primers naar elke put van de PCR plaat. Voeg vervolgens 1 μL van de DNA-verdunning toe in elke put.
- Plaats de PCR-plaat in de thermocycler en voer het bijbehorende PCR-programma uit dat in tabel 1wordt weergegeven.
- Analyseer de versterkte fragmenten. Er zijn voornamelijk twee verschillende manieren om de PCR versterkte fragmenten te analyseren: capillaire elektroforese (CE) met fluorescerende primers of als visualiseren amplicons van agarose gel elektroforese met niet-geëtiketteerde primers.
-
Capillaire elektroforese
- Om de laadbuffer voor te bereiden, mengt u 35 μL gedeïioneerde formamide met 0,45 μL gelabelde maatstandaard. Vortex het reagens goed te mengen, en vervolgens 35,5 μL in de put van de lezer plaat.
- Voeg 1 μL van het PCR-product toe aan de put. Voeg bovendien een druppel minerale olie toe om verdamping van water te voorkomen.
- Bereid de scheidingsplaat voor en voeg de scheidingsbuffer toe.
- Gebruik de commerciële software die bij de genanalyzer is meegeleverd (zie Tabel van materialen). Maak een nieuwe monsterplaat en sla de monsternamen voor alle putten op de plaat.
- Selecteer de analysemethode. In dit geval de monsters ontrijpen bij 90 °C voor 120 s, injecteren bij 2,0 kV voor 30 s en scheiden bij 6,0 kV gedurende 35 minuten.
- Plaats de twee platen in de genanalyzer. Vul de capillaire array met gedestilleerd water.
- Laad de gepatenteerde lineaire polyacrylamide (LPA) gel. Klik tot slot op Uitvoeren.
-
Gel Elektroforese
- Bereid een 1% agarose gel toe toevoegen 1,5 g van moleculaire biologie graad agarose in 150 mL van 1x TAE (Tris-acetaat-EDTA) elektroforese lopende buffer (40 mM Tris, 20 mM azijnzuur, en 1 mM EDTA bij pH 8.0). Los de agarose op door magnetronverwarming gedurende 2-3 min.
- Om het DNA te visualiseren, voeg je 4 μL van een nucleïnezuurvlek toe (zie Tabel van Materialen) en meng voorzichtig.
- Voeg een gelkam, met voldoende putten voor ladders, bedieningselementen en monsters, toe in een gellade. Giet vervolgens langzaam de mix in het midden van de gellade en vermijd bubbels.
- Laat de gel 30-45 minuten afkoelen bij kamertemperatuur tot de gel volledig gestold is. Introduceer de gel in de elektroforese kamer, verwijder de gelkam en vul de kamer met voldoende 1x TAE buffer om de gel te bedekken.
LET OP: Controleer de plaatsing van de gel. De putten moeten dicht bij de negatieve pool worden geplaatst, omdat negatief geladen DNA naar de kathode migreert. - Voeg 5 μL laadbuffer (0,1% (v/v) bromophenolblauw) toe aan de PCR-producten en meng goed.
- Om de grootte van de banden te schatten, laadt u 5 μL dna-moleculaire gewichtsladder (zie Tabel van Materialen).
- Laad de monsters in de extra putten van de gel.
- Zodra alle monsters en de DNA moleculaire gewicht ladder zijn geladen, voer de gel op 90 V voor 1-1,5 uur, totdat de blauwe kleurstof lijn is ongeveer op 75% van de lengte van de gel.
- Visualiseer de banden in een transilluminator voor nucleïnezuren.
-
Capillaire elektroforese
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Bestuivingsstudies bij abrikoos vereisen het gebruik van bloemen in het late ballonstadium een dag voor de anthese(figuur 1A). Deze fase wordt beschouwd als de meest gunstige voor zowel stuifmeel en stamper collectie, omdat florale structuren zijn bijna volwassen, maar anther dehiscence is nog niet opgetreden. Dit voorkomt de interferentie van ongewenste pollen, niet alleen van stuifmeel van dezelfde bloem, maar ook van andere bloemen, omdat de gesloten bloemblaadjes de komst van insecten die extern stuifmeel dragen belemmeren. De stuifmeelkorrels worden gemakkelijk gezeefd door een fijn gaas(figuur 1B)van gedehisceerde anthers die eerder 24 uur op een stuk papier bij kamertemperatuur of met lichte extra warmte werden geplaatst. Ook worden stampers verkregen uit bloemen in het ballonstadium na het verwijderen van bloemblaadjes, kelkbladen en meeldraden met behulp van pincetten of vingernagels (figuur 1C). Stampers kunnen zelf- en kruisbestuiving worden met een fijne borstel(figuur 1D).
De hermafrodische bloemen van abrikoos hebben vijf donkerrode kelkbladen, vijf witte bloemblaadjes (Figuur 1A), een enkele stamper (Figuur 2A) en 25-30 meeldraden. De stamper heeft drie hoofdstructuren: stigma, stijl en eierstok. De eierstok heeft twee ovules, en de bevruchting van ten minste een van hen is nodig voor fruit instelling. Tijdens de bestuiving brengen insecten, voornamelijk bijen, stuifmeelkorrels over naar het stigma (figuur 1A), waar ze binnen24uur na bestuiving ontkiemen ( figuur 2B). Een pollenbuis wordt geproduceerd uit elke ontkiemende stuifmeelkorrel, die door de stamperstructuren groeit om de eierstok na 3-4 dagen te bereiken en één van de twee ovules na ongeveer 7 dagen te bevruchten. In zelfonverenige cultivars waarbij het S-allel van de pollenkorrel hetzelfde is als een van de twee stampers, stopt de pollenbuis met groeien in de bovenste stijl, waardoor bevruchting wordt voorkomen(figuur 2C). Echter, de stuifmeel buizen van een compatibele cultivar, met een ander S allel, kan groeien door de stijl (Figuur 2D),bereiken de eierstok(Figuur 2E) en bevruchten een van de twee eicellen.
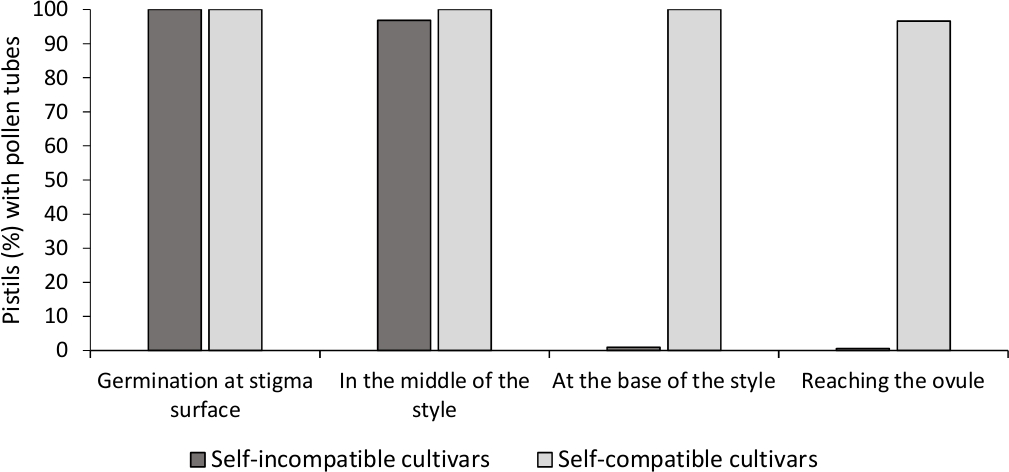
De analyse van in vitro stuifmeel kieming toonde een goede pollen levensvatbaarheid in alle cultivars hier geanalyseerd, omdat de meeste pollen buizen waren langer dan de lengte van het stuifmeel graan na 24 uur in de cultuur medium. Geramineerde stuifmeelkorrels werden waargenomen op het stigmaoppervlak (figuur 2B) in stampers van alle bestuivingen, wat duidt op een adequate bestuiving (figuur 3).
Om de zelf-(in)compatibiliteit voor elke cultivar te bepalen, werd pollenbuisgedrag in zelf- en kruisbestuivingen gedaan in laboratoriumgecontroleerde omstandigheden waargenomen onder fluorescentiemicroscopie. Pollen buis groei werd geregistreerd langs de stijl in alle onderzochte stampers. Cultivars werden beschouwd als zelf-onverenigbaar toen pollen buis groei werd gearresteerd langs de stijl in de meeste zelfbestoven stampers (Figuur 2C, Figuur 3) en zelfcompatibel wanneer ten minste een pollen buis bereikte de basis van de stijl in de meeste van de onderzochte stampers (Figuur 2E, Figuur 3).
De studie van de S-locus door PCR analyse toegestaan karakteriseren van de S-genotypevan elke cultivar. Ten eerste Swerden de S-allelen geïdentificeerd door de versterking van de eerste S-RNase intron met behulp van de primers SRc-F/SRc-R (Tabel 2). De grootte van de versterkte fragmenten werd geanalyseerd door capillaire elektroforese(figuur 4A) en werd gebruikt om de genotypen geanalyseerd in hun overeenkomstige incompatibiliteitsgroep (I.G.) te classificeren (Tabel 3).
Sommige paren van allelen, zoals S1 en S7 of S6 en S9, toonden vergelijkbare fragmentgroottes voor de eerste intron. Zo werd de differentiatie van deze allelen gedaan door het versterken van een regio van de tweede intron van de RNase met de primers Pru-C2/PruC4R, SHLM1/SHLM2 en SHLM3/SHLM4 (Tabel 2). De PruC2/PruC4R primer combinatie werd gebruikt om onderscheid te maken tussen S6 en S9. Voor S6werd een fragment van 1300 bp versterkt, terwijl voor het S9-allel een fragment van ongeveer 700 basispunten werd waargenomen ( figuur4B, tabel 3). De specifieke primers SHLM1/SHLM2 en SHLM3/SHLM4 versterkten een fragment van ongeveer 650 bp in het S1-allel en 413 bp in het S7-allel (figuur 4C, tabel 3).
De primers AprFBC8-(F/R) die de V2- en HVb-variabele gebieden van het SFB-gen versterken, werden gebruikt om Sc- en S8-allelen te onderscheiden, aangezien beide allelen identieke RNase-sequentie vertonen. Het S8-allel vertoonde een PCR-fragment van ongeveer 150 bp, terwijl een fragment van 500 bp overeenkwam met het Sc-allel ( figuur4D, tabel 3). Zodra Sde S-genotype voor alle cultivars werd bepaald, werden zelfoncompatibele cultivars toegewezen aan hun overeenkomstige onverenigbaarheidsgroepen op basis van hun S-alleles( Tabel3).
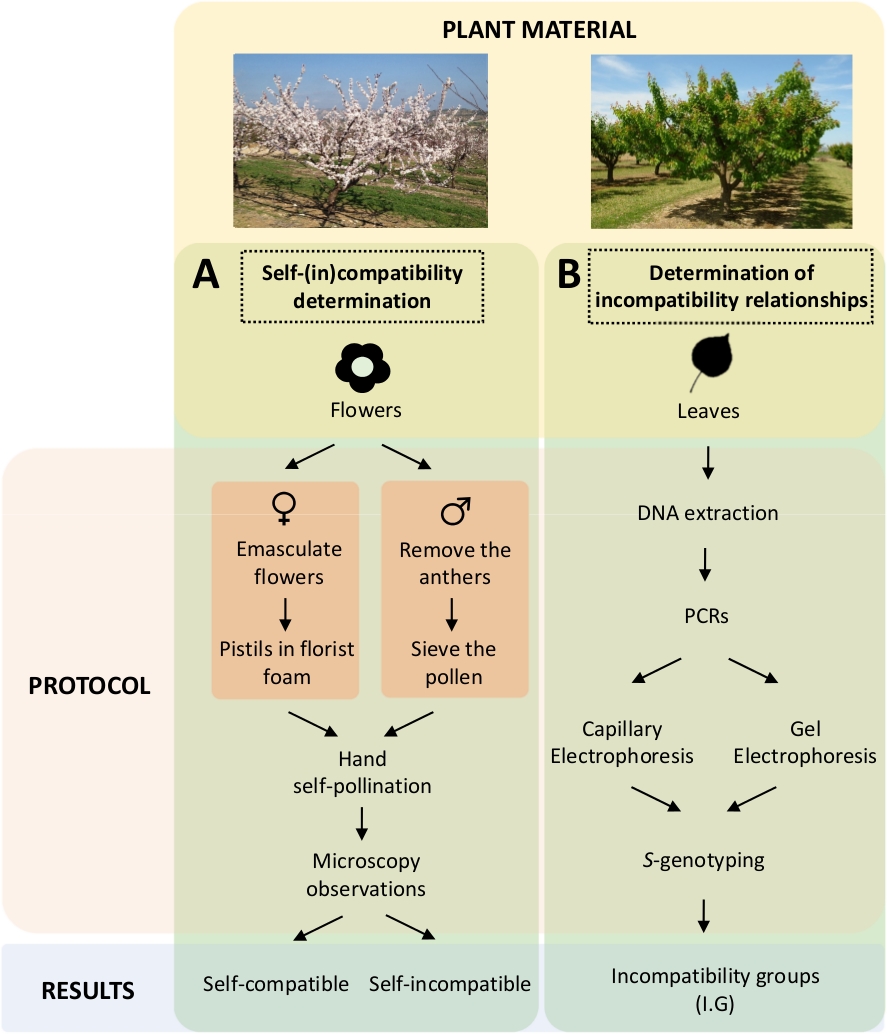
Deze benadering vereist het bepalen van de zelf-(in)compatibiliteit van elke cultivar door gecontroleerde zelfbestuivingen in het laboratorium (figuur 5A) gelijktijdig met de karakterisering van het S-genotypedoor genetische analyse ( figuur5B). Als gevolg hiervan kunnen de bestuivingsvereisten van elke cultivar en de onverenigbaarheidsrelaties tussen abrikozencultiveurs worden bepaald.

Figuur 1. Experimentele opzet voor de bepaling van zelf-(in)compatibiliteit in abrikoos.
(A) Bloemen op ballon stadium (zwarte pijlen) in het veld. (B) Zeef van stuifmeelkorrels met een fijn gaas. (C) Stampers geplaatst op bloemist schuim in water. (D) Handbestuiving van de stampers met behulp van een penseel. Klik hier om een grotere versie van dit cijfer te bekijken.

Figuur 2. Diagrammatische weergave van gametophytische onverenigbaarheidsrelaties in abrikozenbloemen.
(A) In Gametophytic Self-Incompatibility (GSI), zowel compatibele als onverenigbare stuifmeelkorrels ontkiemen op het stigma. Het stuifmeel korrel draagt een van de twee S-allelenvan de oorspronkelijke genotype, in dit geval ofwel S1 of S2. Als het S-allelvan de pollenkorrel overeenkomt met een van de twee S-allelenvan de stamper, in dit geval S1S3,wordt de groei van de pollenbuis geremd in de bovenste een derde van de stijl. (B) Ontkieming van stuifmeelkorrels op het stigmaoppervlak. (C) Pollen buis gearresteerd in de stijl die wijst op een onverenigbaar gedrag. (D) Stuifmeel buizen groeien langs de stijl. (E) Pollen buizen aan de basis van de stijl die een compatibel gedrag aangeeft. Schaalbalken, 100 μm. Klik hier om een grotere versie van dit cijfer te bekijken.

Figuur 3. Representatieve resultaten van pollenkieming en pollenbuisgroei door de stijl voor zelfcompatibele en zelfonverenigbare cultivars na zelfbestuivingen.
Percentage van stampers met stuifmeelkorrels ontkiemen op het stigma oppervlak, met pollen buizen in de helft van de stijl, aan de basis van de stijl, en het bereiken van de ovule. Klik hier om een grotere versie van dit cijfer te bekijken.

Figuur 4. PCR fragment versterking met behulp van vijf Sprimer pair combinaties voor de identificatie van S-alleles.
(A) Gen analyzer output voor de SRc-(F /R) primers met de grootte van de twee versterkte fragmenten van de RNase eerste intron regio die overeenkomt met de S-alleles. S (B) PCR-versterking met behulp van de primers PruC2/PruC4R voor de identificatie van de S6 en S9 allelen. (C) PCR-producten verkregen met behulp van de specifieke primers SHLM1 en SHLM2 voor de differentiatie van het S1-allel en SHLM3 en SHLM4 om het S7-allel te onderscheiden. (D) PCR-versterking met de AprFBC8-(F/R) primers voor het identificeren van Sc en S8 allelen. MI: 1 kb DNA Ladder. MII: 100 bp DNA Ladder. Klik hier om een grotere versie van dit cijfer te bekijken.

Figuur 5. Schema van het experimentele ontwerp om de zelf- en inter-(in) compatibiliteitsrelaties in abrikozencultivivars op te helderen.
(A) Workflow van zelf-(in)compatibiliteitsbepaling door gecontroleerde bestuivingen in het laboratorium. (B) Workflow van de S-allel identificatie door moleculaire benaderingen. Klik hier om een grotere versie van dit cijfer te bekijken.
| PCR-mastermix | Thermocycler-omstandigheden | ||||||
| Onderdelen | Definitieve concentratie | 15 μL reactie | Cyclusstap | Temperatuur | Tijd | Cycli | |
| 10x NH4 Reactiebuffer | 10x | 1,5 μL | Initiële denaturatie | 94 °C | 3 min | 1 | |
| 50 mM MgCl2-oplossing | 25 mM | 1,2 μL | Denatureren | 94 °C | 1 min | 35 | |
| 100 mM dNTP | 2,5 mM | 0,6 μL | Annealing | 55 °C | 1 min | ||
| Primer SRc-F | 10 μM | 0,6 μL | Extensie | 72 °C | 3 min | ||
| Primer SRc-R | 10 μM | 0,6 μL | Laatste uitbreiding | 72 °C | 5 min | 1 | |
| 500 U Taq DNA Polymerase | 0,5 U | 0,2 μL | 4 °C | Houden | |||
| H2O | 8,3 μL | ||||||
| Onderdelen | Definitieve concentratie | 25 μL reactie | Cyclusstap | Temperatuur | Tijd | Cycli | |
| 10x PCR-buffer | 10x | 2,5 μL | Initiële denaturatie | 94 °C | 2 min | 1 | |
| 5x Q-oplossing | 5x | 5 μL | Denatureren | 94 °C | 10 s | 10 | |
| 100 mM dNTP | 2,5 mM | 0,5 μL | Annealing | 55 °C | 2 min | ||
| Primer PruC2 | 10 μM | 0,2 μL | Extensie | 68 °C | 2 min | ||
| Primer C4R | 10 μM | 0,2 μL | Denatureren | 94 °C | 10 s | 25 | |
| 250 U Taq DNA Polymerase | 10 U | 0,13 μL | Annealing | 58 °C | 2 min | ||
| H2O | 15,5 μL | Extensie* | 68 °C | 2 min | |||
| Laatste uitbreiding | 72 °C | 5 min | 1 | ||||
| 4 °C | Houden | ||||||
| * met 10 s toegevoegd elke cyclus aan de 68 %C extensie stap. | |||||||
| Onderdelen | Definitieve concentratie | 25 μL reactie | Cyclusstap | Temperatuur | Tijd | Cycli | |
| 10x PCR-buffer | 10x | 2,5 μL | Initiële denaturatie | 94 °C | 2 min | 1 | |
| 5x Q-oplossing | 5x | 5 μL | Denatureren | 94 °C | 30 s | 35 | |
| 100 mM dNTP | 2,5 mM | 0,5 μL | Annealing | 62 °C | 1,5 min | ||
| Primer SHLM1 | 10 μM | 0,2 μL | Extensie | 72 °C | 2 min | ||
| Primer SHLM2 | 10 μM | 0,2 μL | Laatste uitbreiding | 72 °C | 5 min | 1 | |
| 250 U Taq DNA Polymerase | 10 U | 0,13 μL | 4 °C | Houden | |||
| H2O | 15,5 μL | ||||||
| Onderdelen | Definitieve concentratie | 20 μL reactie | Cyclusstap | Temperatuur | Tijd | Cycli | |
| 5x PCR-buffer | 5x | 4 μL | Initiële denaturatie | 98 °C | 30 s | 1 | |
| dNTP | 2,5 mM | 1,6 μL | Denatureren | 98 °C | 10 s | 35 | |
| Primer SHLM3 | 10 μM | 1 μL | Annealing | 51 °C | 30 s | ||
| Primer SHLM4 | 10 μM | 1 μL | Extensie | 72 °C | 1 min | ||
| 100 U DNA Polymerase | 5 U | 0,2 μL | Laatste uitbreiding | 72 °C | 5 min | 1 | |
| H2O | 12,4 μL | 4 °C | Houden | ||||
| Onderdelen | Definitieve concentratie | 25 μL reactie | Cyclusstap | Temperatuur | Tijd | Cycli | |
| 10x PCR-buffer | 10x | 2,5 μL | Initiële denaturatie | 94 °C | 2 min | 1 | |
| 100 mM dNTP | 2,5 mM | 2 μL | Denatureren | 94 °C | 30 s | 35 | |
| Primer FBC8-F | 10 μM | 1 μL | Annealing | 55 °C | 1,5 min | ||
| Primer FBC8-R | 10 μM | 1 μL | Extensie | 72 °C | 2 min | ||
| 250 U Taq DNA Polymerase | 10 U | 0,125 μL | Laatste uitbreiding | 72 °C | 5 min | 1 | |
| H2O | 17,4 μL | 4 °C | Houden | ||||
Tabel 1. Reactie- en fietsomstandigheden voor verschillende primercombinaties die in dit protocol worden gebruikt.
| Inleidingen | Volgorde | Verwijzing |
| SRc-F | 5'-CTCGCTTTTTTTGTTCTTGC-3' | 18 |
| SRc-R | 5'-GGCCATTGTTGCACCCCTTG-3' | 18 |
| Pru-C2 | 5'-CTTTGGCCAAGTAATTATTCAAACC-3' | 35 |
| Pru-C4R | 5'-GGATGTGGTACGATTGAAGCG-3' | 35 |
| SHLM1-F | 5'-GGTGGGTGATAAGGTAGCC-3' | 17 |
| SHLM2-R SHLM2-R | 5'-GGCTGCATAAGGAAGCTGTAGG-3' | 17 |
| SHLM3-F | 5'-TATATCTTACTCTTTGGC-3' | 17 |
| SHLM4-R SHLM4-R | 5'-CACTATAATGTGTATG-3' | 17 |
| AprFBC8-F | 5'-CATGGAAAAAGCTGACTTATGG-3' | 26 |
| AprFBC8-R | 5'-GCCTCTAATGTCTACTCTTAG-3' | 26 |
Tabel 2. Primers gebruikt in dit protocol, volgorde en referentie voor S-genotype karakterisering in Prunus armeniëca.
| Cultivar | SRc-(F/R) (bp) | PruC2/PruC4R (bp) | SHLM1/SHLM2 (bp) | SHLM3/SHLM4 (bp) | AprFBC8-(F/R) (bp) | S-Genotype S-Genotype | Incompatibiliteitsgroep (I.G) |
| Wonder Cot8 | 420, 420 | 749, 1386 | S6S9 | Viii | |||
| Magische wieg8 | 334, 420 | 749 | S2S9 | Xx | |||
| Goldstrike8 | 334, 420 | 749 | S2S9 | - | |||
| T06917 | 334, 408 | 650 | S1S2 | I | |||
| T12017 | 334, 408 | 650 | S1S2 | - | |||
| C-6 | 334, 408 | 413 | S2S7 | Iv | |||
| Cooper Cot8 | 274, 408 | 650 | S1S3 | Xviii | |||
| Apriqueen | 358, 358 | 500 | ScSc | - | |||
| Bergecot8 | 334, 358 | 500 | S2Sc | - | |||
| Lente Blush8 | 274, 358 | 150 | S3S8 | Xxi |
Tabel 3. S-genotyping van abrikozencultivatoren met vijf primerparen die in dit protocol en onverenigbaarheidsgroeptoewijzing worden gebruikt. S De verschillende polymerase kettingreactie Sproductmaten van S-alleles versterkt met behulp van SRc-(F/R), PruC2/PruC4R, SHLM1/ SHLM2, SHLM3/SHLM4 en AprFBC8-(F/R) primers worden weergegeven in de tabel.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Traditioneel, de meeste commerciële abrikozen Europese cultivars waren zelf-compatibel36. Niettemin heeft het gebruik van Noord-Amerikaanse zelfonverenige cultivars als ouders in fokprogramma's in de afgelopen decennia geresulteerd in de release van een toenemend aantal nieuwe zelfonverenige cultivars met onbekende bestuivingsvereisten7,8,37. Zo wordt de bepaling van zelf- en inter-(in)compatibiliteitsrelaties in abrikozencultiveurs steeds belangrijker. Dit wordt geaccentueerd in die gebieden waar de winter koelen afneemt, omdat hoge jaar tot jaar variaties in de tijd van de bloei zijn het voorkomen van het toeval in de bloei van cultivars en hun pollenisators in veel gevallen, vooral in cultivars met hoge koelen eisen38. De hierin beschreven methode, waarbij handbestuiving, microscopie en genetische analyses worden gecombineerd, is zeer nuttig geweest om de zelf(in)compatibiliteit van elke cultivar te bepalen en de potentiële bestuivingscultivatorcultivatoren te bepalen.
Bestuivingsvereisten kunnen worden bepaald door middel van veldcontrole-experimenten in boomgaardomstandigheden11,39. Echter, de expositie naar externe factoren, waaronder meteorologische ongunstige omstandigheden kan leiden tot bestuiving falen10, wat kan resulteren in onjuiste diagnoses van zelf-onverenigbaarheid. De hierin beschreven methode maakt het mogelijk om zelf-(in)compatibiliteit nauwkeuriger te evalueren door microscopie-waarnemingen van handbestovende bloemen in laboratoriumgecontroleerde omstandigheden, waardoor milieu-invloed wordt vermeden. Bovendien maakt deze aanpak het analyseren van een hoger aantal cultivars per jaar mogelijk, omdat slechts een klein aantal bloemen nodig is in plaats van meerdere volwassen bomen voor elke cultivar die nodig zijn inveldexperimenten 40.
Incompatibiliteitsrelaties kunnen worden vastgesteld door handbestuivingen en microscopie14. Bestuivingen kunnen echter slechts voor een korte periode worden uitgevoerd tijdens het bloeiseizoen in het voorjaar, en volwassen bomen in de buurt van het laboratorium zijn nodig, omdat de levensduur van de verzamelde bloemen zeer kort is. Zo is het aantal incompatibiliteitsrelaties dat kan worden geanalyseerd door gecontroleerde handbestuivingen in elk seizoen zeer laag. De karakterisering van de genen gecodeerd door de S-locusheeft de ontwikkeling van PCR-gebaseerde methoden voor S-allelgenotyping18,41mogelijk gemaakt . Deze aanpak Sversnelt S-allel identificatie omdat het geen bloemen nodig heeft, en de experimenten kunnen worden uitgevoerd met een vegetatief weefsel42. Dit verlengt de periode waarin plantaardig materiaal, meestal jonge bladeren, kan worden verzameld43. Bovendien kunnen de bladeren worden gelyofof bevroren, zodat de analyse op elk moment van het jaar kan worden uitgevoerd, in tegenstelling tot bestuivingen die alleen op verse bloemen kunnen worden gedaan tijdens het bloeiseizoen44. Een bijkomend voordeel is dat bladeren kunnen worden verzameld van jonge bomen, zelfs voordat het invoeren van de bloei leeftijd, het vergemakkelijken van de verzameling van monsters en het vroeg verkrijgen van de resultaten45.
De genetische analyse maakt een betere differentiatie van zelf-onverenigbaarheid allelen mogelijk, omdat het nauwkeurige resultaten van versterkte fragmentgroottes21,46biedt . Tot op heden zijn drieëndertig S-allelengeïdentificeerd in abrikozen12,18,,20,21,,22,23,24, waardoor 36 onverenigbaarheidsgroepen op basis van S-genotype8,9,,17,25,26,27zijn opgericht . Aan de andere kant is een nadeel van deze methode dat verschillende allelen in dezelfde grootte of mutaties ten onrechte kunnen worden geïdentificeerd als hetzelfde allel. Zo zijn Sc en S8 allelen identiek voor de RNase sequentie, maar een invoegsel met 358 basis is te vinden in het SFB-gen van Sc19. Ook de eerste intron regio van de allelen S1 en S7 zijn identiek en zijn niet te onderscheiden met behulp van de primers SRc-F/SRc-R. Daarnaast zijn verschillende homologieën, zoals S6 en S528 of S20 en S55, en S7, S13 (EF062341) en S4617, gevonden omdat sommige van deze allelen gedeeltelijk zijn gesequenced of door storingen tijdens PCR-versterking en dus verder werk nodig is om ze correct te onderscheiden.
PCR-analyse en S-RNase-sequencing zijn toereikend om onverenigbaarheidsrelaties tot stand te brengen door de identificatie van S-allelenen de toewijzing van cultivars in hun overeenkomstige Onverenigbaarheidsgroep8,17,26,27. Deze methode heeft echter de beperking om de bepaling van de zelf-(in)compatibiliteit voor bepaalde abrikozencultivivars te voorkomen. Zelfcompatibiliteit (SC) is in verband gebracht met bepaalde S-allelenin andere Prunus-soorten 47, zoals amandel (Sf)48,49 of zoete kers (S4')50,51. In abrikoos kan het Sc-allel, dat is gekoppeld aan SC21,echter ten onrechte worden geïdentificeerd als S8, een zelfincompatibel allel19,22en mogelijke mutaties die niet verband houden met de S-locus, aangezien de M-locus12,52, verlenend SC zijn geïdentificeerd. Sc Onlangs is Mde M-locus gegenyped met behulp van SSR markers12. Daarom moet de genetische identificatie van SC voor abrikozengenypes verder onderzoek doen en om fouten als gevolg van factoren die niet verband houden met de S locus te voorkomen, is in dit werk de karakterisering van zelf-(in)compatibiliteit ook bepaald door het gedrag van de pollenbuizen te phenotyping door het stamper van zelfbestuivende bloemen.
De hier beschreven methode die de bepaling van zelf-(in)compatibiliteit door handbestuivingen in laboratoriumomstandigheden combineert met de daaropvolgende observatie van het gedrag van stuifmeelbuizen Sin de stamper van gecontroleerde zelfbestuivingen onder de fluorescentiemicroscopie en de identificatie van het S-genotype door PCR-analyse maakt het mogelijk om de bestuivingsvereisten van abrikozencultivivars vast te stellen. Dit biedt een waardevolle informatie voor telers en kwekers, omdat het mogelijk maakt de incompatibiliteitsrelaties tussen cultivars vast te stellen om geschikte bestuivers te kiezen bij het ontwerp van nieuwe boomgaarden en om geschikte ouders te selecteren om nieuwe kruisen te ontwerpen in abrikozenfokkerijprogramma's.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
De auteurs hebben niets te onthullen.
Acknowledgments
Dit onderzoek werd gefinancierd door Ministerio de Ciencia, Innovación y Universidades-European Regional Development Fund, European Union (AGL2016-77267-R en AGL2015-74071-JIN); Instituto Nacional de Investigación y Tecnología Agraria y Alimentaria (RFP2015-00015-00, RTA2017-00003-00); Gobierno de Aragón-Europees Sociaal Fonds, Europese Unie (Grupo Consolidado A12_17R), Fundación Biodiversidad en Agroseguro S.A.
Materials
| Name | Company | Catalog Number | Comments |
| Agarose D1 Low EEO | Conda | 8010.22 | |
| BIOTAQ DNA Polymerase kit | Bioline | BIO-21060 | |
| Bright field microscope | Leica Microsystems | DM2500 | |
| CEQ System Software | Beckman Coulter | ||
| DNeasy Plant Mini Kit | QIAGEN | 69106 | |
| dNTP Set, 4 x 25 µmol | Bioline | BIO-39025 | |
| GenomeLab DNA Size Standard Kit - 400 | Beckman Coulter | 608098 | |
| GenomeLab GeXP Genetic Analysis System | Beckman Coulter | ||
| GenomeLab Separation Buffer | Beckman Coulter | 608012 | |
| GenomeLab Separation Gel LPA-1 | Beckman Coulter | 391438 | |
| HyperLadder 100bp | Bioline | BIO-33029 | |
| HyperLadder 1kb | Bioline | BIO-33025 | |
| Image Analysis System | Leica Microsystems | ||
| Molecular Imager VersaDoc MP 4000 system | Bio-Rad | 170-8640 | |
| NanoDrop One Spectrophotometer | Thermo Fisher Scientific | 13-400-518 | |
| pH-Meter BASIC 20 | Crison | ||
| Phusion High-Fidelity PCR Kit | Thermo Fisher Scientific | F553S | |
| Power Pack P 25 T | Biometra | ||
| Primer Forward | Isogen Life Science | ||
| Primer Reverse | Isogen Life Science | ||
| Quantity One Software | Bio-Rad | ||
| Stereoscopic microscope | Leica Microsystems | MZ-16 | |
| Sub-Cell GT | Bio-Rad | ||
| SYBR Safe DNA Gel Stain | Thermo Fisher Scientific | S33102 | |
| T100 Thermal Cycler | Bio-Rad | 1861096 | |
| Taq DNA Polymerase | QIAGEN | 201203 | |
| Vertical Stand Autoclave | JP Selecta |
References
- Silva, N. F., Goring, D. R.
- Charlesworth, D., Vekemans, X., Castric, V., Glémin, S. Plant self-incompatibility systems: A molecular evolutionary perspective. New phytologist. 168, 61-69 (2005).
- Tao, R., et al. Identification of stylar RNases associated with gametophytic self-incompatibility in almond (Prunus dulcis). Plant and Cell Physiology. 38, 304-311 (1997).
- Ushijima, K., et al. Structural and transcriptional analysis of the self-incompatibility locus of almond: Identification of a pollen-expressed F-box gene with haplotype-specific polymorphism. The Plant cell. 15, 771-781 (2003).
- Bedinger, P. A., Broz, A. K., Tovar-Mendez, A., McClure, B. Pollen-Pistil Interactions and Their Role in Mate Selection. Plant Physiology. 173, 79-90 (2017).
- Guerra, M. E., Rodrigo, J. Japanese plum pollination: A review. Scientia Horticulturae. 197, 674-686 (2015).
- Zhebentyayeva, T., Ledbetter, C., Burgos, L., Llacer, G. Apricot. Fruit Breeding. Badenes, M. L., Byrne, D. , Springer. Boston, MA. 415-458 (2012).
- Herrera, S., Lora, J., Hormaza, J. I., Herrero, M., Rodrigo, J. Optimizing Production in the New Generation of Apricot Cultivars: Self-incompatibility, S-RNase Allele Identification, and Incompatibility Group Assignment. Frontiers in Plant Science. 9, 527 (2018).
- Egea, J., Burgos, L. Detecting Cross-incompatibility of Three North American Apricot Cultivars and Establishing the First Incompatibility Group in Apricot. Journal of the American Society for Horticultural Science. 121, 1002-1005 (1996).
- Rodrigo, J., Herrero, M. Effects of pre-blossom temperatures on flower development and fruit set in apricot. Scientia Horticulturae. 92, 125-135 (2002).
- Julian, C., Herrero, M., Rodrigo, J. Flower bud differentiation and development in fruiting and non-fruiting shoots in relation to fruit set in apricot (Prunus armeniaca). Trees. 24, 833-841 (2010).
- Muñoz-Sanz, J. V., Zuriaga, E., López, I., Badenes, M. L., Romero, C. Self-(in)compatibility in apricot germplasm is controlled by two major loci, S and M. BMC Plant Biology. 17, 82 (2017).
- Burgos, L., Berenguer, T., Egea, J. Self- and Cross-compatibility among Apricot Cultivars. HortScience. 28, 148-150 (1993).
- Rodrigo, J., Herrero, M. Evaluation of pollination as the cause of erratic fruit set in apricot "Moniqui". Journal of Horticultural Science. 71, 801-805 (1996).
- Milatović, D., Nikolić, D., Krška, B. Testing of self-(in)compatibility in apricot cultivars from European breeding programmes. Horticultural Science. 40 (2), 65-71 (2013).
- Milatović, D., Nikolić, D., Fotirić-Aksić, M., Radović, A. Testing of self-(in)compatibility in apricot cultivars using fluorescence microscopy. Acta Scientiarum Polonorum, Hortorum Cultus. 12 (6), 103-113 (2013).
- Herrera, S., Rodrigo, J., Hormaza, J. I., Lora, J. Identification of Self-Incompatibility Alleles by Specific PCR Analysis and S-RNase Sequencing in Apricot. Int J Mol Sci. 19, 3612 (2018).
- Romero, C., et al. Analysis of the S-locus structure in Prunus armeniaca L. Identification of S-haplotype specific S-RNase and F-box genes. Plant Molecular Biology. 56, 145-157 (2004).
- Halász, J., Pedryc, A., Hegedus, A. Origin and dissemination of the pollen-part mutated SC haplotype which confers self-compatibility in apricot (Prunus armeniaca). New Phytologist. 176, 792-803 (2007).
- Halász, J., Hegedus, A., Hermán, R., Stefanovits-Bányai, É, Pedryc, A. New self-incompatibility alleles in apricot (Prunus armeniaca L.) revealed by stylar ribonuclease assay and S-PCR analysis. Euphytica. 145, 57-66 (2005).
- Vilanova, S., Romero, C., Llacer, G., Badenes, M. L., Burgos, L. Identification of Self-(in)compatibility Alleles in Apricot by PCR and Sequence Analysis. Journal of the American Society for Horticultural Science. 130, 893-898 (2005).
- Feng, J., et al. Detection and transcript expression of S-RNase gene associated with self-incompatibility in apricot (Prunus armeniaca L.). Molecular Biology Reports. 33, 215-221 (2006).
- Zhang, L., et al. Identification of self-incompatibility (S-) genotypes of Chinese apricot cultivars. Euphytica. 160, 241-248 (2008).
- Wu, J., et al. Identification of S-haplotype-specific S-RNase and SFB alleles in native Chinese apricot (Prunus armeniaca L). Journal of Horticultural Science and Biotechnology. 84, 645-652 (2009).
- Szabó, Z., Nyéki, J. Blossoming, fructification and combination of apricot varieties. Acta Horticulturae. 293, 295-302 (1991).
- Halász, J., Pedryc, A., Ercisli, S., Yilmaz, K. U., Hegedűs, A. S-genotyping supports the genetic relationships between Turkish and Hungarian apricot germplasm. Journal of the American Society for Horticultural Science. 135, 410-417 (2010).
- Lachkar, A., et al. Identification of self-(in)compatibility S-alleles and new cross-incompatibility groups in Tunisian apricot (Prunus armeniaca L.) cultivars. The Journal of Horticultural Science and Biotechnology. 88, 497-501 (2013).
- Pérez-Pastor, A., Ruiz-Sánchez, M. C., Domingo, R., Torrecillas, A. Growth and phenological stages of Búlida apricot trees in South-East. Agronomie. 24, 93-100 (2004).
- Williams, J. H., Friedman, W. E., Arnold, M. L. Developmental selection within the angiosperm style: using gamete DNA to visualize interspecific pollen competition. Proceedings of the National Academy of Sciences of the United States of America. 96, 9201-9206 (1999).
- Julian, C., Herrero, M., Rodrigo, J. Anther meiosis time is related to winter cold temperatures in apricot (Prunus armeniaca L.). Environmental and Experimental Botany. 100, 20-25 (2014).
- Guerra, M. E., López-Corrales, M., Wünsch, A., Rodrigo, J. Lack of Fruit Set Caused by Ovule Degeneration in Japanese Plum. Journal of the American Society for Horticultural Science. 136 (6), 375-381 (2011).
- Guerra, M. E., Wünsch, A., López-Corrales, M., Rodrigo, J. Flower Emasculation as the Cause for Lack of Fruit Set in Japanese Plum Crosses. Journal of the American Society for Horticultural Science. 135 (6), 556-562 (2010).
- Hormaza, J. I., Pinney, K., Polito, V. S. Correlation in the tolerance to ozone between sporophytes and male gametophytes of several fruit and nut tree species (Rosaceae). Sexual Plant Reproduction. 9, 44-48 (1996).
- Alcaraz, M. L., Hormaza, J. I., Rodrigo, J. Pistil Starch Reserves at Anthesis Correlate with Final Flower Fate in Avocado (Persea americana). PLOS ONE. 8 (10), 78467 (2013).
- Tao, R., et al. Molecular typing of S-alleles through Identification, Characterization and cDNA cloning for S-RNases in Sweet Cherry. Journal of the American Society for Horticultural Science. 124, 224-233 (1999).
- Burgos, L., et al. The self-compatibility trait of the main apricot cultivars and new selections from breeding programmes. Journal of Horticultural Science. 72, 147-154 (1997).
- Hormaza, J. I., Yamane, H., Rodrigo, J. Apricot. Genome Mapping and Molecular Breeding in Plants, Volume 4 Fruits and Nuts. Kole, C. , Springer. New York, NY. 171-187 (2007).
- Benmoussa, H., Ghrab, M., Ben Mimoun, M., Luedeling, E. Chilling and heat requirements for local and foreign almond (Prunus dulcis Mill.) cultivars in a warm Mediterranean location based on 30 years of phenology records. Agricultural and Forest Meteorology. 239, 34-46 (2017).
- Rodrigo, J., Herrero, M., Hormaza, J. I. Pistil traits and flower fate in apricot (Prunus armeniaca). Annals of Applied Biology. 154, 365-375 (2009).
- Williams, R. R. Techniques used in fruit-set experiments. Towards Regulated Cropping. Williams, R. R., Wilson, D. , Grower Books. London. 57-61 (1970).
- Sutherland, B. G., Robbins, T. P., Tobutt, K. R. Primers amplifying a range of Prunus S-alleles. Plant Breeding. 123, 582-584 (2004).
- Murray, M. G., Thompson, W. F. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Research. 8, 4321-4325 (1980).
- Porebski, S., Bailey, L. G., Baum, B. R. Modification of a CTAB DNA Extraction Protocol for Plants Containing High Polysaccharide and Polyphenol Components. Plant Molecular Biology Reporter. 15 (1), 8-15 (1997).
- Rogers, S. O., Bendich, A. J. Extraction of DNA from milligram amounts of fresh, herbarium and mummified plant tissues. Plant Molecular Biology. 5 (2), 69-76 (1985).
- Hormaza, J. I. Molecular characterization and similarity relationships among apricot (Prunus armeniaca L.) genotypes using simple sequence repeats. Theoretical and Applied Genetics. 104, 321-328 (2002).
- Sonneveld, T., Tobutt, K. R., Robbins, T. P. Allele-specific PCR detection of sweet cherry self-incompatibility (S) alleles S1 to S16 using consensus and allele-specific primers. Theoretical and Applied Genetics. 107, 1059-1070 (2003).
- Hegedus, A., Lénárt, J., Halász, J. Sexual incompatibility in Rosaceae fruit tree species: molecular interactions and evolutionary dynamics. Biologia Plantarum. 56 (2), 201-209 (2012).
- Fernández i Martí, A., Gradziel, T. M., Socias i Company, R. Methylation of the Sf locus in almond is associated with S-RNase loss of function. Plant Molecular Biology. 86, 681-689 (2014).
- Company, R. S. i, Kodad, O., Martí, A. F. i, Alonso, J. M. Mutations conferring self-compatibility in Prunus species: From deletions and insertions to epigenetic alterations. Scientia Horticulturae. 192, 125-131 (2015).
- Boskovic, R., Tobutt, K. R. Correlation of stylar ribonuclease zymograms with incompatibility alleles in sweet cherry. Euphytica. 90, 245-250 (1996).
- Cachi, A. M., Wünsch, A. S-genotyping of sweet cherry varieties from Spain and S-locus diversity in Europe. Euphytica. 197 (2), 229-236 (2014).
- Zuriaga, E., et al. An S-locus Independent Pollen Factor Confers Self-Compatibility in "Katy" Apricot. PLoS ONE. 8 (1), 53947 (2013).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}