

Summary
Nous présentons une méthodologie pour établir les exigences de pollinisation des cultivarsd’abricot (Prunus armeniaca L.) combinant la détermination de l’auto-compatibilité par microscopie de fluorescence avec l’identification du S-génotype par analyse pcr.
Abstract
L’autocompatibilité dans rosaceae est déterminée par un système d’auto-incompatibilité gametophytique (GSI) qui est principalement contrôlé par le locus multiallelique S. Dans l’abricot, la détermination des relations d’auto-compatibilité et d’intercompatibilité est de plus en plus importante, puisque la libération d’un nombre important de nouveaux cultivars a entraîné l’augmentation des cultivars ayant des exigences de pollinisation inconnues. Ici, nous décrivons une méthodologie qui combine la détermination de l’auto-compatibilité par pollinisation manuelle et microscopie avec l’identification du génotype Spar l’analyse pcr. Pour la détermination de l’auto-compatibilité, les fleurs à l’étape du ballon de chaque cultivar ont été recueillies dans le champ, pollinisées à la main en laboratoire, fixes et tachées de bleu aniline pour l’observation du comportement du tube pollin sous la microscopie de fluorescence. Pour l’établissement de relations d’incompatibilité entre les cultivars, Sl’ADN de chaque cultivar a été extrait de jeunes feuilles et S-allèles ont été identifiés par PCR. Cette approche permet d’établir des groupes d’incompatibilité et d’élucider les relations d’incompatibilité entre les cultivars, ce qui fournit une information précieuse pour choisir les pollinisateurs appropriés dans la conception de nouveaux vergers et pour sélectionner les parents appropriés dans les programmes d’élevage.
Introduction
L’autocompatibilité est une stratégie des plantes à fleurs pour prévenir l’autopollinisation et promouvoir la croisement1. Dans rosaceae, ce mécanisme est déterminé par un système d’auto-incompatibilité gametophytique (GSI) qui est principalement contrôlé par le locus multiallelique S2. Dans le style, le gène RNase code le déterminant sylar S-s,un RNase3, tandis qu’une protéine F-box, qui détermine le déterminant du pollen S,est codifiée par le gène4de la SFB . L’interaction d’auto-incompatibilité a lieu par l’inhibition de la croissance du tube pollinique le long du style empêchant la fertilisation de l’ovule5,6.
En abricot, un renouvellement variétal a eu lieu dans le monde entier au cours des deux dernières décennies7,8. Cette introduction d’un nombre important de nouveaux cultivars, provenant de différents programmes d’élevage publics et privés, a entraîné l’augmentation des cultivars d’abricots ayant des besoins inconnus de pollinisation8.
Différentes méthodologies ont été utilisées pour déterminer les exigences de pollinisation dans l’abricot. Sur le terrain, la compatibilité de soi peut être établie par des pollinisations contrôlées dans des arbres en cage ou dans des fleurs émasculées et en enregistrant par la suite le pourcentage de fruits ensemble9,10,11,12. En outre, des pollinisations contrôlées ont été effectuées en laboratoire par culture semi-in vivo des fleurs et l’analyse du comportement du tube pollinique sous microscopie de fluorescence8,13,14,15,16,17. Récemment, les techniques moléculaires, telles que l’analyse et le séquençage du PCR, ont permis la caractérisation des relations d’incompatibilité basées sur l’étude des gènes RNase et SFB 18,19. En abricot, trente-trois S-allèles ont été signalés (S1 à S20, S22 à S30, S52, S53, Sv, Sx), y compris un allèle lié à l’auto-compatibilité (Sc)12,18,20,21,22,23,24. Jusqu’à présent, 26 groupes d’incompatibilité ont été stablis dans cette espèce selon le S-génotype8,9,17,25,26,27. Les cultivars avec les mêmes allèles Ssont inter-incompatibles, tandis que les cultivars avec au moins un S-allèledifférent et, par conséquent, répartis dans différents groupes incompatibles, sont intercompatibles.
Pour définir les exigences de pollinisation des cultivars d’abricots, nous décrivons une méthodologie qui combine la détermination de l’auto-compatibilité par microscopie de fluorescence avec l’identification du génotype Spar analyse pcr dans les cultivars d’abricot. Cette approche permet d’établir des groupes d’incompatibilité et d’élucider les relations d’incompatibilité entre les cultivars.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. Détermination de la compatibilité autonome
- Goûtez les fleurs dans le champ. Il est nécessaire de recueillir les fleurs au stade du ballon ( Figure 1A), correspondant àl’étape58 sur l’échelle BBCH pour l’abricot28, pour éviter la pollinisation préalable non désirée.
-
Autopollinisations en laboratoire
- Retirez les anths des fleurs au stade du ballon et placez-les sur un morceau de papier pour les sécher à température de laboratoire.
- Après 24 h, tamiser les grains de pollen à l’aide d’un maillage fin (0,26 mm) (figure 1B).
- Émasculer un groupe de 30 fleurs au même stade de ballon pour chaque auto-pollinisation et pollinisation croisée et placer les pistils sur la mousse de fleuriste dans l’eau à température de laboratoire (Figure 1C).
- Polliniser les pistils à la main à l’aide d’un pinceau avec du pollen provenant de fleurs du même cultivar 24 h après l’émasculation. En outre, polliniser un autre ensemble de pistils de chaque cultivar avec le pollen des fleurs d’un pollinisateur compatible comme contrôle (Figure 1D).
- Après 72 h, fixer les pistils dans une solution fixative d’éthanol/acide acétique (3:1) pendant au moins 24 h à 4 °C29. Ensuite, jetez le fixatif et ajoutez 75% d’éthanol en veillant à ce que les échantillons soient complètement immergés dans la solution. Les échantillons peuvent être conservés dans cette solution à 4 °C jusqu’à l’utilisation8,17,30,31,32.
-
Évaluation de la viabilité du pollen par germination in vitro de pollen
- Pour préparer le milieu de germination, poids 25 g de saccharose, 0,075 g d’acide borique (H3BO3) et 0,075 g de nitrate de calcium (Ca(NO3)2)33.
- Ajouter les composants du milieu dans 250 mL d’eau distillée et dissoudre complètement.
- Solidifier le milieu en ajoutant 2 g d’agarose et mélanger en tourbillonnant.
- Vérifiez le pH du milieu à l’aide d’un pH mètre et réglez la valeur à 7,0 avec la solution NaOH ou HCl.
- Autoclaver le mélange pour stériliser le milieu.
- Après l’autoclavage, refroidir le milieu et le distribuer dans les plats Petri dans un capot stérile de flux laminaire.
- Dispersez les grains de pollen des mêmes cultivars utilisés pour les pollinisations contrôlées dans le milieu de germination du pollen solidifié et observez-les au microscope après 24 h6.
REMARQUE : Pour stériliser le capot d’écoulement laminaire, nettoyez la surface avec 70 % d’éthanol et allumez la lampe UV pendant 10 min. - Conserver les plats Petri au réfrigérateur à 4 °C jusqu’à l’utilisation.
-
Observations de microscopie
- Laver les pistils trois fois pendant 1 h avec de l’eau distillée et les laisser dans 5% de sulfite de sodium à 4 °C. Après 24 h, autoclavez-les à 1 kg/cm2 pendant 10 min de sulfite de sodium pour adoucir les tissus34.
- Placez les pistils autoclaved sur une lame de verre et, à l’aide d’un scalpel, retirez les trichomes autour de l’ovaire pour obtenir une meilleure visualisation des tubes de pollen. Ensuite, écraser les pistils avec un verre à couverture.
- Préparer 0,1% (v/v) tache bleue aniline: mélanger 0,1 mL de bleu aniline en 100 mL de 0,1 N phosphate de potassium tribasique (K3PO4). Appliquer une goutte de bleu aniline sur les préparations pour tacher les dépôts d’callose pendant la croissance du tube pollinique.
- Observez les tubes de pollen le long du style au microscope avec l’épifluorescence UV à l’aide de 340-380 filtres à bande et 425 filtres longpass.
2. Extraction d’ADN
- Échantillonner 2-3 feuilles dans le champ au printemps. Il est recommandé d’échantillonner les feuilles à de jeunes stades puisque l’ADN obtenu est de meilleure qualité et des niveaux inférieurs de composés phénoliques par rapport aux vieilles feuilles.
- Extraire l’ADN génomique suivant les étapes décrites dans un kit disponible dans le commerce (voir tableau des matériaux).
- Analyser la quantité et la qualité des concentrations d’ADN à l’aide du spectrophotomètre UV-vis (260 nm).
3. Identification s-allèle
-
Configuration des réactions PCR
- Préparer une dilution de 50 ng/μL dans l’eau distillée de chaque échantillon d’extraction d’ADN.
- Décongeler les réactifs PCR lentement et les garder sur la glace. Laisser la polymérase d’ADN dans le congélateur jusqu’à ce que nécessaire.
- Préparer les réactions d’amplification à l’aide des différentes combinaisons d’amorces. Créez le mélange de réactions PCR en combinant les composants du tableau 1. Vortex la réaction PCR mélanger bien et de distribuer le volume indiqué pour les différentes combinaisons d’amorces à chaque puits de la plaque PCR. Ensuite, ajouter 1 μL de la dilution de l’ADN dans chaque puits.
- Placez la plaque PCR dans le thermocycleur et exécutez le programme PCR correspondant indiqué dans le tableau 1.
- Analyser les fragments amplifiés. Il existe principalement deux façons différentes d’analyser les fragments amplifiés pcr : l’électrophorèse capillaire (CE) avec des amorces fluorescentes ou comme amplisons les amypplicons de l’électrophorèse de gel agarose avec des amorces non étiquetées.
-
Électrophorèse capillaire
- Pour préparer le tampon de chargement, mélanger 35 μL de formamide déionisé avec 0,45 μL de norme de dimensionnement étiquetée. Vortex le réactif pour bien mélanger, puis distribuer 35,5 μL dans le puits de la plaque du lecteur.
- Ajouter 1 μL du produit PCR dans le puits. En outre, ajouter une goutte d’huile minérale pour prévenir l’évaporation de l’eau.
- Préparer la plaque de séparation en ajoutant un tampon de séparation.
- Utilisez le logiciel commercial inclus avec l’analyseur de gènes (voir Tableau des matériaux). Créez une nouvelle plaque d’échantillonnage et enregistrez les noms d’échantillons pour tous les puits de la plaque.
- Sélectionnez la méthode d’analyse. Dans ce cas, dénaturer les échantillons à 90 °C pendant 120 s, injecter à 2,0 kV pour 30 s et les séparer à 6,0 kV pendant 35 min.
- Insérez les deux plaques dans l’analyseur de gènes. Remplissez le tableau capillaire d’eau distillée.
- Chargez le gel breveté de polyacrylamide linéaire (LPA). Enfin, cliquez sur Exécuter.
-
Gel Électrophorèse
- Préparer un gel agarose de 1 % en ajoutant 1,5 g d’agarose de qualité biologie moléculaire dans 150 mL d’électrophorèse TAE (Tris-acétate-EDTA) tampon en cours d’exécution (40 mM Tris, acide acétique de 20 mM et 1 mM EDTA à pH 8,0). Dissoudre l’agarose par chauffage au micro-ondes pendant 2-3 min.
- Pour visualiser l’ADN, ajouter 4 μL d’une tache d’acide nucléique (voir tableau des matériaux) et mélanger doucement.
- Ajouter un peigne à gel, avec suffisamment de puits pour les échelles, les contrôles et les échantillons, dans un plateau de gel. Ensuite, verser lentement le mélange dans le milieu du plateau de gel et éviter les bulles.
- Laisser refroidir le gel pendant 30-45 min à température ambiante jusqu’à ce que le gel se solidifie complètement. Introduire le gel dans la chambre électrophorèse, enlever le peigne à gel et remplir la chambre avec suffisamment de tampon TAE 1x pour couvrir le gel.
REMARQUE : Vérifiez le placement du gel. Les puits doivent être placés près du poteau négatif puisque l’ADN chargé négativement migre vers la cathode. - Ajouter 5 μL de tampon de chargement (0,1 % (v/v) bleu bromophénol) aux produits PCR et bien mélanger.
- Pour estimer la taille des bandes, chargez 5 μL de l’échelle de poids moléculaire de l’ADN(voir tableau des matériaux).
- Chargez les échantillons dans les puits supplémentaires du gel.
- Une fois que tous les échantillons et l’échelle de poids moléculaire de l’ADN sont chargés, exécutez le gel à 90 V pendant 1-1,5 h, jusqu’à ce que la ligne de colorant bleu soit approximativement à 75% de la longueur du gel.
- Visualisez les bandes dans un transilluminateur pour les acides nucléiques.
-
Électrophorèse capillaire
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Les études de pollinisation de l’abricot nécessitent l’utilisation de fleurs au stade avancé du ballon un jour avant l’anthèse (Figure 1A). Cette étape est considérée comme la plus favorable à la fois pour la collection de pollen et de pistil, puisque les structures florales sont presque matures, mais la déhiscence anther n’a pas encore eu lieu. Cela empêche l’interférence du pollen indésirable, non seulement du pollen de la même fleur, mais aussi d’autres fleurs, puisque les pétales fermés empêchent l’arrivée d’insectes transportant du pollen externe. Les grains de pollen sont facilement tamisés à travers un maillage fin (Figure 1B) à partir d’anthères déhisqués précédemment placés sur un morceau de papier pendant 24 h à température ambiante ou avec une légère chaleur supplémentaire. De même, les pistils sont obtenus à partir de fleurs au stade du ballon après l’enlèvement des pétales, des sépales et des étamines à l’aide de pinces ou d’ongles (Figure 1C). Les pistils peuvent être auto-pollinisés et croisés à l’aide d’une brosse fine (figure 1D).
Les fleurs hermaphrodéditiques d’abricot ont cinq sépales rouge foncé, cinq pétales blancs (figure 1A), un pistil unique (Figure 2A) et 25-30 étamines. Le pistil a trois structures principales : la stigmatisation, le style et l’ovaire. L’ovaire a deux ovules, et la fertilisation d’au moins un d’entre eux est nécessaire pour l’établissement des fruits. Pendant la pollinisation, les insectes, principalement les abeilles, transfèrent les grains de pollen à la stigmatisation (figure 1A), où ils germent (figure 2B) dans les 24 heures suivant la pollinisation. Un tube de pollen est produit à partir de chaque grain de pollen germinant, qui se développe à travers les structures pistil pour atteindre l’ovaire après 3-4 jours et fertiliser l’un des deux ovules après environ 7 jours. Dans les cultivars auto-incompatibles dans lesquels l’allèle S du grain de pollen est le même que l’un des deux pistils, tube de pollen cesse de croître au niveau supérieur, empêchant la fécondation (Figure 2C). Cependant, les tubes de pollen d’un cultivar compatible, avec un allèle S différent, peuvent se développer à travers le style (Figure 2D), atteindre l’ovaire (Figure 2E) et fertiliser l’un des deux ovules.
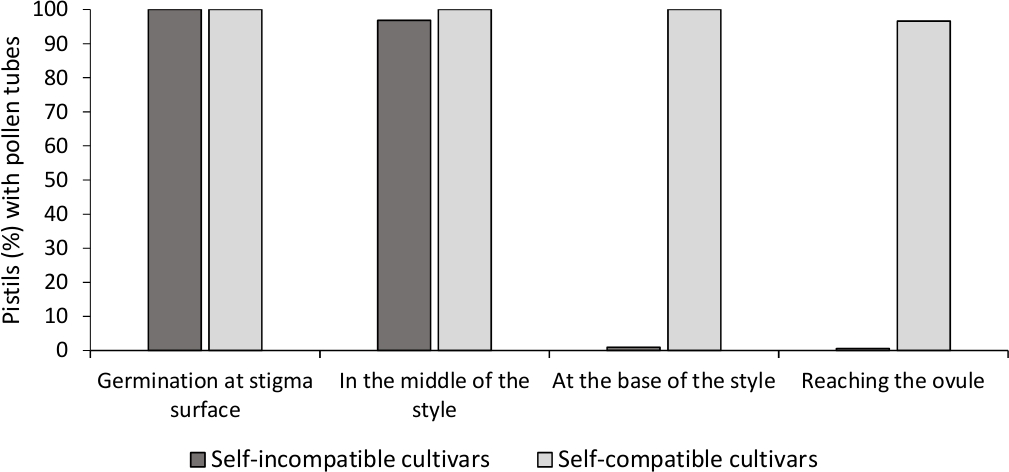
L’analyse de la germination in vitro du pollen a montré une bonne viabilité du pollen dans tous les cultivars analysés ici, puisque la plupart des tubes de pollen étaient plus longs que la longueur du grain de pollen après 24 h dans le milieu de culture. Des grains de pollen germés ont été observés à la surface de la stigmatisation (figure 2B) dans les pistils de toutes les pollinisations, ce qui indique une pollinisation adéquate (figure 3).
Pour déterminer l’auto-compatibilité pour chaque cultivar, le comportement du tube pollinique dans les pollinisations auto-et croisées effectuées dans des conditions contrôlées en laboratoire a été observé sous microscopie de fluorescence. La croissance de tube de pollen a été enregistrée le long du modèle dans tous les pistils examinés. Les cultivars étaient considérés comme auto-incompatibles lorsque la croissance du tube pollinique a été arrêtée le long du style dans la plupart des pistils auto-pollinisés (figure 2C, figure 3) et auto-compatibles lorsqu’au moins un tube de pollen a atteint la base du style dans la plupart des pistils examinés (figure 2E, figure 3).
L’étude du S-locus par analyse pcr a permis de caractériser le S-génotype de chaque cultivar. Tout d’abord, les Alles Sont été identifiés par l’amplification du premier intron S-RNase à l’aide des amorces SRc-F/SRc-R (Tableau 2). La taille des fragments amplifiés a été analysée par électrophorèse capillaire (figure 4A) et a été utilisée pour classer les génotypes analysés dans leur groupe d’incompatibilité correspondant (I.G.) (Tableau 3).
Certaines paires d’allèles, comme S1 et S7 ou S6 et S9, ont montré des tailles de fragments similaires pour le premier intron. Ainsi, la différenciation de ces allèles a été faite en amplifiant une région du deuxième intron du RNase avec les amorces Pru-C2/Prruc4R, SHLM1/SHLM2 et SHLM3/SHLM4 (Tableau 2). La combinaison d’amorce PruC2/Prruc4R a été utilisée pour distinguer S6 et S9. Pour S6, un fragment de 1300 pb a été amplifié alors qu’un fragment d’environ 700 pb a été observé pour l’allèle S9 ( figure4B, tableau 3). Les amorces spécifiques SHLM1/SHLM2 et SHLM3/SHLM4 ont amplifié un fragment d’environ 650 pb dans l’allèle S1 et de 413 pb dans l’allèle S7 (Figure 4C, Tableau 3).
Les amorces AprFBC8-(F/R) qui amplifient les régions variables V2 et HVb du gène SFB ont été utilisées pour distinguer les allèles Sc et S8 puisque les deux allèles présentent une séquence RNase identique. L’allèle S8 présentait un fragment de PCR d’environ 150 pb alors qu’un fragment de 500 pb correspondait à l’allèle Sc (Figure 4D, Tableau 3). Une fois que le génotype Sa été déterminé pour tous les cultivars, les cultivars auto-incompatibles ont été affectés à leurs groupes d’incompatibilité correspondants en fonction de leurs allèles S( Tableau3).
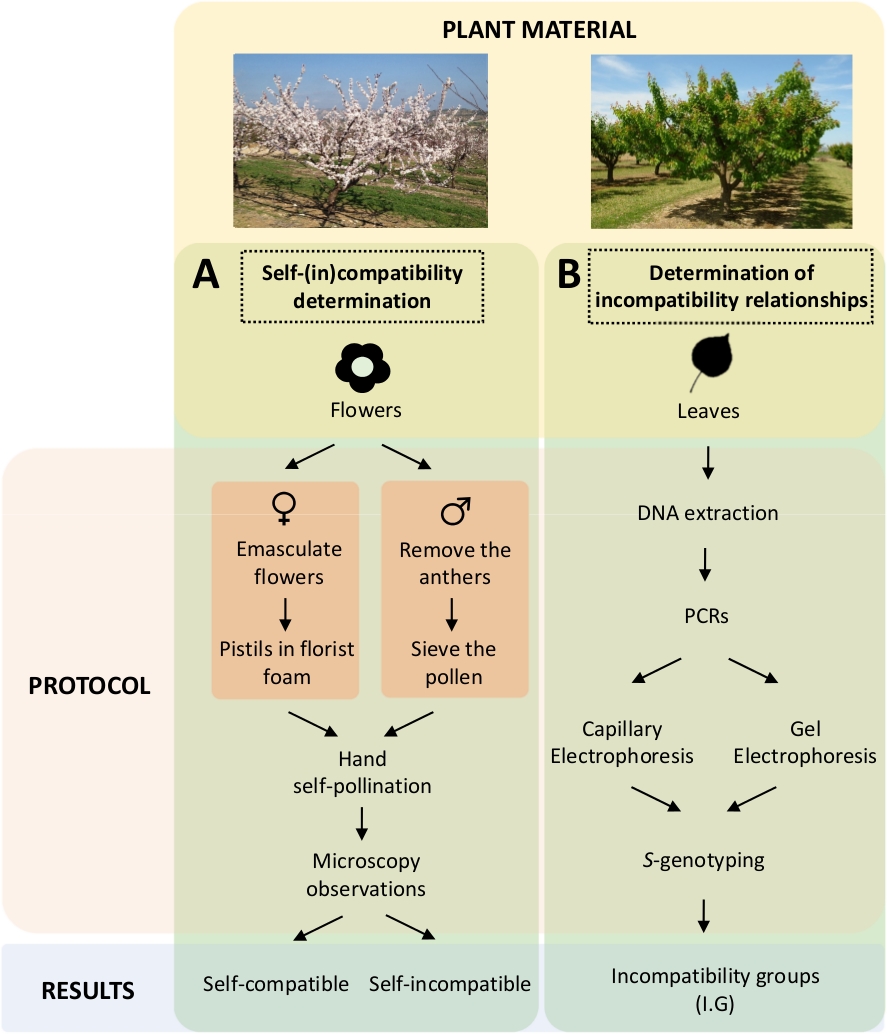
Cette approche exige de déterminer l’auto-compatibilité de chaque cultivar par auto-pollinisation contrôlée et croisée en laboratoire (figure 5A) en même temps que la caractérisation du S-génotypepar analyse génétique ( figure5B). En conséquence, les exigences de pollinisation de chaque cultivar et les relations d’incompatibilité entre les cultivars d’abricot peuvent être déterminées.

Figure 1. Mise en place expérimentale pour la détermination de l’auto-compatibilité dans l’abricot.
(A) Fleurs au stade de ballon (flèches noires) dans le champ. (B) Tamis des grains de pollen à l’aide d’un maillage fin. (C) Pistils placés sur la mousse de fleuriste dans l’eau. (D) Pollinisation manuelle des pistils à l’aide d’un pinceau. Veuillez cliquer ici pour voir une version plus grande de ce chiffre.

Figure 2. Représentation schématique des relations d’incompatibilité gametophytique dans les fleurs d’abricot.
(A) Dans l’auto-incompatibilité gametophytique (GSI), les grains de pollen compatibles et incompatibles germent sur la stigmatisation. Le grain de pollen porte l’un des deux S-allèles du génotype d’origine, en l’occurrence S1 ou S2. S Si le S-allèledu grain de pollen correspond à l’un des deux SS-allèles du pistil, dans ce cas S1S3, la croissance du tube de pollen est inhibée dans le tiers supérieur du style. S (B) Germination de grains de pollen à la surface de la stigmatisation. (C) Tube de pollen arrêté dans le style indiquant un comportement incompatible. (D) Tubes de pollen qui poussent le long du style. (E) Tubes de pollen à la base du style indiquant un comportement compatible. Barres d’échelle, 100 μm. Veuillez cliquer ici pour voir une version plus grande de ce chiffre.

Figure 3. Résultats représentatifs de la germination du pollen et de la croissance du tube pollinique à travers le style des cultivars auto-compatibles et auto-incompatibles après l’auto-pollinisation.
Pourcentage de pistils avec des grains de pollen qui germent à la surface de la stigmatisation, avec des tubes de pollen à mi-chemin du style, à la base du style, et atteignant l’ovule. Veuillez cliquer ici pour voir une version plus grande de ce chiffre.

Figure 4. Amplification des fragments PCR à l’aide de cinq combinaisons de paires d’amorces pour l’identification des allèles S.
(A) Sortie d’analyseur de gènes pour les amorces SRc-(F/R) indiquant la taille des deux fragments amplifiés de la première région intron de RNase correspondant aux allèles S.. (B) amplification pcr utilisant les amorces PrulC2/PrulC4R pour l’identification des allèles S6 et S9. (C) produits PCR obtenus à l’aide des amorces spécifiques SHLM1 et SHLM2 pour la différenciation de l’allèle S1 et SHLM3 et SHLM4 pour distinguer l’allèle S7. (D) Amplification pcr avec les amorces AprFBC8-(F/R) pour l’identification des allèles Sc et S8. MI: Échelle d’ADN de 1 kb. MII: Échelle d’ADN de 100 pb. Veuillez cliquer ici pour voir une version plus grande de ce chiffre.

Figure 5. Schéma de la conception expérimentale pour élucider les relations de compatibilité auto- et inter-(in) dans les cultivars d’abricot.
(A)Flux de travail de détermination de l’auto-compatibilité par pollinisation contrôlée en laboratoire. (B) Flux de travail de l’identification de l’allèle Spar des approches moléculaires. Veuillez cliquer ici pour voir une version plus grande de ce chiffre.
| PCR Master Mix | Conditions thermocycler | ||||||
| Composants | Concentration finale | 15 μL de réaction | Étape de cycle | Température | Heure | Cycles | |
| Tampon de réaction 10x NH4 | 10x | 1,5 μL | Dénaturation initiale | 94 °C | 3 min | 1 | |
| 50 mM MgCl2 Solution | 25 mM | 1,2 μL | Dénaturation | 94 °C | 1 min | 35 | |
| 100 mM dNTP | 2,5 mM | 0,6 μL | Recuit | 55 °C | 1 min | ||
| Amorce SRc-F | 10 μM | 0,6 μL | Extension | 72 °C | 3 min | ||
| Amorce SRc-R | 10 μM | 0,6 μL | Prolongation finale | 72 °C | 5 min | 1 | |
| 500 U Taq ADN Polymérase | 0,5 U | 0,2 μL | 4 °C | Tenir | |||
| H2O | 8,3 μL | ||||||
| Composants | Concentration finale | 25 μL de réaction | Étape de cycle | Température | Heure | Cycles | |
| Mémoire tampon PCR 10x | 10x | 2,5 μL | Dénaturation initiale | 94 °C | 2 min | 1 | |
| Solution Q 5x | 5x | 5 μL | Dénaturation | 94 °C | 10 s | 10 | |
| 100 mM dNTP | 2,5 mM | 0,5 μL | Recuit | 55 °C | 2 min | ||
| Amorce Pruc2 | 10 μM | 0,2 μL | Extension | 68 °C | 2 min | ||
| Amorce C4R | 10 μM | 0,2 μL | Dénaturation | 94 °C | 10 s | 25 | |
| 250 U Taq ADN Polymérase | 10 U | 0,13 μL | Recuit | 58 °C | 2 min | ||
| H2O | 15,5 μL | Extension* | 68 °C | 2 min | |||
| Prolongation finale | 72 °C | 5 min | 1 | ||||
| 4 °C | Tenir | ||||||
| * avec 10 s ajouté chaque cycle à l’étape d’extension de 68 %C. | |||||||
| Composants | Concentration finale | 25 μL de réaction | Étape de cycle | Température | Heure | Cycles | |
| Mémoire tampon PCR 10x | 10x | 2,5 μL | Dénaturation initiale | 94 °C | 2 min | 1 | |
| Solution Q 5x | 5x | 5 μL | Dénaturation | 94 °C | 30 s | 35 | |
| 100 mM dNTP | 2,5 mM | 0,5 μL | Recuit | 62 °C | 1,5 min | ||
| Amorce SHLM1 | 10 μM | 0,2 μL | Extension | 72 °C | 2 min | ||
| Amorce SHLM2 | 10 μM | 0,2 μL | Prolongation finale | 72 °C | 5 min | 1 | |
| 250 U Taq ADN Polymérase | 10 U | 0,13 μL | 4 °C | Tenir | |||
| H2O | 15,5 μL | ||||||
| Composants | Concentration finale | 20 μL de réaction | Étape de cycle | Température | Heure | Cycles | |
| Mémoire tampon PCR 5x | 5x | 4 μL | Dénaturation initiale | 98 °C | 30 s | 1 | |
| dNTP | 2,5 mM | 1,6 μL | Dénaturation | 98 °C | 10 s | 35 | |
| Amorce SHLM3 | 10 μM | 1 μL | Recuit | 51 °C | 30 s | ||
| Amorce SHLM4 | 10 μM | 1 μL | Extension | 72 °C | 1 min | ||
| 100 U ADN Polymérase | 5 U | 0,2 μL | Prolongation finale | 72 °C | 5 min | 1 | |
| H2O | 12,4 μL | 4 °C | Tenir | ||||
| Composants | Concentration finale | 25 μL de réaction | Étape de cycle | Température | Heure | Cycles | |
| Mémoire tampon PCR 10x | 10x | 2,5 μL | Dénaturation initiale | 94 °C | 2 min | 1 | |
| 100 mM dNTP | 2,5 mM | 2 μL | Dénaturation | 94 °C | 30 s | 35 | |
| Amorce FBC8-F | 10 μM | 1 μL | Recuit | 55 °C | 1,5 min | ||
| Amorce FBC8-R | 10 μM | 1 μL | Extension | 72 °C | 2 min | ||
| 250 U Taq ADN Polymérase | 10 U | 0,125 μL | Prolongation finale | 72 °C | 5 min | 1 | |
| H2O | 17,4 μL | 4 °C | Tenir | ||||
Tableau 1. Conditions de réaction et de vélo pour différentes combinaisons d’amorces utilisées dans ce protocole.
| Amorces | Séquence | Référence |
| SRc-F | 5'-CTCGCTTTCCTTTTTTTTGC-3' | 18 |
| SRc-R | 5'-GGCCATTGTGCACCCCTTG-3' | 18 |
| Pru-C2 | 5'-CTTTGGCCAAGTAATTTCAAACC-3' | 35 |
| Pru-C4R | 5'-GGATGTGTGAGATTGAAGG-3' | 35 |
| SHLM1-F | 5'-GGTGGAGGTGATAAGGTAGCC-3' | 17 |
| SHLM2-R | 5'-GGCTGCATAAGGAAGCTTTAGG-3' | 17 |
| SHLM3-F | 5'-TATATCTACTCTTTGGGC-3' | 17 |
| SHLM4-R | 5'-CCATATGATAATGTGTGTRAG-3' | 17 |
| AprFBC8-F | 5'-CAGGAAAAGCTGACTTATTAT-3' | 26 |
| AprFBC8-R | 5'-GCCTCTAATGTCATCTACTCTACTTAG-3' | 26 |
Tableau 2. Amorces utilisées dans ce protocole, séquence et référence pour la caractérisation du génotype Sdans Prunus armeniaca.
| Cultivar | SRc-(F/R) (pb) | Prruc2/Pruc4R (bp) | SHLM1/SHLM2 (pb) | SHLM3/SHLM4 (pb) | AprFBC8-(F/R) (pb) | S-Génotype | Groupe d’incompatibilité (I.G) |
| Wonder Cot8 | 420, 420 | 749, 1386 | S6S9 | Viii | |||
| Magic Cot8 | 334, 420 | 749 | S2S9 | Xx | |||
| Goldstrike8 | 334, 420 | 749 | S2S9 | - | |||
| T06917 | 334, 408 | 650 | S1S2 | Ⅰ | |||
| T12017 | 334, 408 | 650 | S1S2 | - | |||
| C-6 | 334, 408 | 413 | S2S7 | Iv | |||
| Cooper Cot8 | 274, 408 | 650 | S1S3 | Xviii | |||
| Apriqueen | 358, 358 | 500 | ScSc | - | |||
| Bergecot8 | 334, 358 | 500 | S2Sc | - | |||
| Blush printanier8 | 274, 358 | 150 | S3S8 | Xxi |
Tableau 3. S-génotypage des cultivars d’abricot avec cinq paires d’amorces utilisées dans ce protocole et affectation de groupe d’incompatibilité. Les différentes tailles de produits Sde réaction en chaîne de polymérase de S-allèles amplifiés à l’aide d’amorces SRc-(F/R), Prul2/Pruc4R, SHLM1/ SHLM2, SHLM3/SHLM4 et AprFBC8-(F/R) sont indiquées dans le tableau.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Traditionnellement, la plupart des cultivars européens d’abricots commerciaux étaient auto-compatibles36. Néanmoins, l’utilisation de cultivars auto-incompatibles nord-américains comme parents dans les programmes d’élevage au cours des dernières décennies a entraîné la libération d’un nombre croissant de nouveaux cultivars auto-incompatibles ayant des exigences de pollinisation inconnues7,8,37. Ainsi, la détermination des relations d’auto et d’intercompatibilité dans les cultivars d’abricot est de plus en plus importante. Cela est accentué dans les zones où le refroidissement hivernal diminue, puisque les variations élevées d’une année à l’autre dans le temps de floraison empêchent la coïncidence dans la floraison des cultivars et de leurs pollinisateurs dans de nombreux cas, en particulier dans les cultivars ayant des exigences de refroidissement élevées38. La méthodologie décrite dans le présent, combinant pollinisation manuelle, microscopie et analyses génétiques, a été très utile pour déterminer la compatibilité de chaque cultivar et établir ses cultivars pollinisateurs potentiels.
Les exigences de pollinisation peuvent être déterminées par des expériences de contrôle sur le terrain dans des conditions de verger11,39. Toutefois, l’exposition à des facteurs externes, y compris les conditions météorologiques défavorables peuvent causer l’échec de pollinisation10, qui peut entraîner des diagnostics erronés d’auto-incompatibilité. La méthodologie décrite dans le présent document permet d’évaluer plus précisément la compatibilité de soi par des observations de microscopie de fleurs pollinisées à la main dans des conditions contrôlées en laboratoire, évitant ainsi l’influence environnementale. En outre, cette approche permet d’analyser un plus grand nombre de cultivars par an, puisque seul un petit nombre de fleurs est nécessaire au lieu de plusieurs arbres adultes pour chaque cultivar qui sont nécessaires dans les expériences sur le terrain40.
Des relations d’incompatibilité peuvent être établies combinant pollinisations manuelles et microscopie14. Cependant, les pollinisations ne peuvent être effectuées que pendant une courte période pendant la saison de floraison au printemps, et des arbres adultes près du laboratoire sont nécessaires, puisque la durée de vie des fleurs recueillies est très courte. Ainsi, le nombre de relations d’incompatibilité qui peuvent être analysées par des pollinisations manuelles contrôlées à chaque saison est très faible. La caractérisation des gènes codés par le S-locus a permis le développement de méthodes basées sur pcr pour le génotypage S-allele18,41. S Cette approche accélère l’identification s-allèlecar elle ne nécessite pas de fleurs, et les expériences peuvent être effectuées avec n’importe quel tissu végétatif42. Cela prolonge la période pendant laquelle le matériel végétal, généralement de jeunes feuilles, peut être recueilli43. En outre, les feuilles peuvent être lyophilisées ou congelées, de sorte que l’analyse peut être faite à tout moment de l’année, contrairement aux pollinisations qui ne peuvent être faites sur les fleurs fraîches pendant la saison de floraison44. Un avantage supplémentaire est que les feuilles peuvent être prélevées sur les jeunes arbres avant même d’entrer en âge de fleurir, ce qui facilite la collecte des échantillons et l’obtention précoce des résultats45.
L’analyse génétique permet une meilleure différenciation des allèles d’auto-incompatibilité puisqu’elle fournit des résultats précis des tailles de fragments amplifiés21,46. À ce jour, trente-trois S-allèles ont été identifiés dans l’abricot12,18,20,21,22,23,24, qui a permis d’établir 36 groupes d’incompatibilité basés sur S-génotype8,9,17,25,26,27. S D’autre part, un inconvénient de cette méthodologie est que différents allèles dans la même taille de gamme ou des mutations peuvent être identifiés à tort comme le même allèle. Ainsi, les allèles Sc et S8 sont identiques pour la séquence RNase mais une insertion de 358 pb se trouve dans le gène SFB de Sc19. De même, la première région intron des allèles S1 et S7 sont identiques et sont indiscernables à l’aide des amorces SRc-F/SRc-R. En outre, plusieurs homologies, telles que S6 et S528 ou S20 et S55, et S7, S13 (EF062341) et S4617, ont été trouvés parce que certains de ces allèles ont été partiellement séquencés ou par des défaillances lors de l’amplification pcr et, par conséquent, d’autres travaux sont nécessaires pour les distinguer correctement.
L’analyse pcr et le séquençage S-RNase sont suffisants pour établir des relations d’incompatibilité par l’identification des allèles Set l’allocation des cultivars dans leur groupe d’incompatibilité correspondant8,17,26,27. Toutefois, cette méthode limite la prévention de la détermination de l’auto-compatibilité pour des cultivars d’abricots particuliers. L’autocompatibilité (SC) a été associée à des allèles S-allèles particuliers chez d’autres espèces de Prunus 47, comme amande (Sf)48,49 ou cerise douce (S4')50,51. S Cependant, dans l’abricot, l’allèle Sc, qui a été associé à SC21, peut être identifié à tort comme S8, un allèle auto-incompatible19,22, et les mutations possibles non liées au Locus, comme le M-locus12,52, conférant SC ont été identifiés. Récemment, Mle M-locus a été génotype à l’aide de marqueurs SSR12. Par conséquent, l’identification génétique de SC pour les génotypes d’abricot a besoin de recherches plus poussées et, afin d’éviter les erreurs dues à des facteurs non liés au Locus S, dans ce travail, la caractérisation de l’auto-compatibilité a également été déterminée par le phénotypage du comportement des tubes de pollen à travers le pistil des fleurs auto-pollinisées.
La méthodologie décrite ci-après combinant la détermination de l’auto-compatibilité par pollinisation manuelle dans des conditions de laboratoire avec l’observation ultérieure du comportement des tubes polliniques dans le pistil des autopollinisations contrôlées sous la microscopie de fluorescence et l’identification du s-génotype par analyse PCR permet d’établir les exigences de pollinisation des cultivars d’abricot. S Cela fournit une information précieuse pour les producteurs et les éleveurs, car il permet d’établir les relations d’incompatibilité entre les cultivars de choisir des pollinisateurs appropriés dans la conception de nouveaux vergers ainsi que de sélectionner les parents appropriés pour concevoir de nouvelles croix dans les programmes d’élevage d’abricots.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Les auteurs n’ont rien à divulguer.
Acknowledgments
Cette recherche a été financée par Ministerio de Ciencia, Innovacion y Universidades-European Regional Development Fund, Union européenne (AGL2016-77267-R, et AGL2015-74071-JIN); Instituto Nacional de Investigación y Tecnología Agraria y Alimentaria (DFP2015-00015-00, RTA2017-00003-00); Gobierno de Aragón-European Social Fund, Union européenne (Grupo Consolidado A12_17R), Fundación Biodiversidad et Agroseguro S.A.
Materials
| Name | Company | Catalog Number | Comments |
| Agarose D1 Low EEO | Conda | 8010.22 | |
| BIOTAQ DNA Polymerase kit | Bioline | BIO-21060 | |
| Bright field microscope | Leica Microsystems | DM2500 | |
| CEQ System Software | Beckman Coulter | ||
| DNeasy Plant Mini Kit | QIAGEN | 69106 | |
| dNTP Set, 4 x 25 µmol | Bioline | BIO-39025 | |
| GenomeLab DNA Size Standard Kit - 400 | Beckman Coulter | 608098 | |
| GenomeLab GeXP Genetic Analysis System | Beckman Coulter | ||
| GenomeLab Separation Buffer | Beckman Coulter | 608012 | |
| GenomeLab Separation Gel LPA-1 | Beckman Coulter | 391438 | |
| HyperLadder 100bp | Bioline | BIO-33029 | |
| HyperLadder 1kb | Bioline | BIO-33025 | |
| Image Analysis System | Leica Microsystems | ||
| Molecular Imager VersaDoc MP 4000 system | Bio-Rad | 170-8640 | |
| NanoDrop One Spectrophotometer | Thermo Fisher Scientific | 13-400-518 | |
| pH-Meter BASIC 20 | Crison | ||
| Phusion High-Fidelity PCR Kit | Thermo Fisher Scientific | F553S | |
| Power Pack P 25 T | Biometra | ||
| Primer Forward | Isogen Life Science | ||
| Primer Reverse | Isogen Life Science | ||
| Quantity One Software | Bio-Rad | ||
| Stereoscopic microscope | Leica Microsystems | MZ-16 | |
| Sub-Cell GT | Bio-Rad | ||
| SYBR Safe DNA Gel Stain | Thermo Fisher Scientific | S33102 | |
| T100 Thermal Cycler | Bio-Rad | 1861096 | |
| Taq DNA Polymerase | QIAGEN | 201203 | |
| Vertical Stand Autoclave | JP Selecta |
References
- Silva, N. F., Goring, D. R. Mechanisms of self-incompatibility in flowering plants. Cellular and Molecular Life Sciences. 58, 1988-2007 (2001).
- Charlesworth, D., Vekemans, X., Castric, V., Glémin, S. Plant self-incompatibility systems: A molecular evolutionary perspective. New phytologist. 168, 61-69 (2005).
- Tao, R., et al. Identification of stylar RNases associated with gametophytic self-incompatibility in almond (Prunus dulcis). Plant and Cell Physiology. 38, 304-311 (1997).
- Ushijima, K., et al. Structural and transcriptional analysis of the self-incompatibility locus of almond: Identification of a pollen-expressed F-box gene with haplotype-specific polymorphism. The Plant cell. 15, 771-781 (2003).
- Bedinger, P. A., Broz, A. K., Tovar-Mendez, A., McClure, B. Pollen-Pistil Interactions and Their Role in Mate Selection. Plant Physiology. 173, 79-90 (2017).
- Guerra, M. E., Rodrigo, J. Japanese plum pollination: A review. Scientia Horticulturae. 197, 674-686 (2015).
- Zhebentyayeva, T., Ledbetter, C., Burgos, L., Llacer, G. Apricot. Fruit Breeding. Badenes, M. L., Byrne, D. , Springer. Boston, MA. 415-458 (2012).
- Herrera, S., Lora, J., Hormaza, J. I., Herrero, M., Rodrigo, J. Optimizing Production in the New Generation of Apricot Cultivars: Self-incompatibility, S-RNase Allele Identification, and Incompatibility Group Assignment. Frontiers in Plant Science. 9, 527 (2018).
- Egea, J., Burgos, L. Detecting Cross-incompatibility of Three North American Apricot Cultivars and Establishing the First Incompatibility Group in Apricot. Journal of the American Society for Horticultural Science. 121, 1002-1005 (1996).
- Rodrigo, J., Herrero, M. Effects of pre-blossom temperatures on flower development and fruit set in apricot. Scientia Horticulturae. 92, 125-135 (2002).
- Julian, C., Herrero, M., Rodrigo, J. Flower bud differentiation and development in fruiting and non-fruiting shoots in relation to fruit set in apricot (Prunus armeniaca). Trees. 24, 833-841 (2010).
- Muñoz-Sanz, J. V., Zuriaga, E., López, I., Badenes, M. L., Romero, C. Self-(in)compatibility in apricot germplasm is controlled by two major loci, S and M. BMC Plant Biology. 17, 82 (2017).
- Burgos, L., Berenguer, T., Egea, J. Self- and Cross-compatibility among Apricot Cultivars. HortScience. 28, 148-150 (1993).
- Rodrigo, J., Herrero, M. Evaluation of pollination as the cause of erratic fruit set in apricot "Moniqui". Journal of Horticultural Science. 71, 801-805 (1996).
- Milatović, D., Nikolić, D., Krška, B. Testing of self-(in)compatibility in apricot cultivars from European breeding programmes. Horticultural Science. 40 (2), 65-71 (2013).
- Milatović, D., Nikolić, D., Fotirić-Aksić, M., Radović, A. Testing of self-(in)compatibility in apricot cultivars using fluorescence microscopy. Acta Scientiarum Polonorum, Hortorum Cultus. 12 (6), 103-113 (2013).
- Herrera, S., Rodrigo, J., Hormaza, J. I., Lora, J. Identification of Self-Incompatibility Alleles by Specific PCR Analysis and S-RNase Sequencing in Apricot. Int J Mol Sci. 19, 3612 (2018).
- Romero, C., et al. Analysis of the S-locus structure in Prunus armeniaca L. Identification of S-haplotype specific S-RNase and F-box genes. Plant Molecular Biology. 56, 145-157 (2004).
- Halász, J., Pedryc, A., Hegedus, A. Origin and dissemination of the pollen-part mutated SC haplotype which confers self-compatibility in apricot (Prunus armeniaca). New Phytologist. 176, 792-803 (2007).
- Halász, J., Hegedus, A., Hermán, R., Stefanovits-Bányai, É, Pedryc, A. New self-incompatibility alleles in apricot (Prunus armeniaca L.) revealed by stylar ribonuclease assay and S-PCR analysis. Euphytica. 145, 57-66 (2005).
- Vilanova, S., Romero, C., Llacer, G., Badenes, M. L., Burgos, L. Identification of Self-(in)compatibility Alleles in Apricot by PCR and Sequence Analysis. Journal of the American Society for Horticultural Science. 130, 893-898 (2005).
- Feng, J., et al. Detection and transcript expression of S-RNase gene associated with self-incompatibility in apricot (Prunus armeniaca L.). Molecular Biology Reports. 33, 215-221 (2006).
- Zhang, L., et al. Identification of self-incompatibility (S-) genotypes of Chinese apricot cultivars. Euphytica. 160, 241-248 (2008).
- Wu, J., et al. Identification of S-haplotype-specific S-RNase and SFB alleles in native Chinese apricot (Prunus armeniaca L). Journal of Horticultural Science and Biotechnology. 84, 645-652 (2009).
- Szabó, Z., Nyéki, J. Blossoming, fructification and combination of apricot varieties. Acta Horticulturae. 293, 295-302 (1991).
- Halász, J., Pedryc, A., Ercisli, S., Yilmaz, K. U., Hegedűs, A. S-genotyping supports the genetic relationships between Turkish and Hungarian apricot germplasm. Journal of the American Society for Horticultural Science. 135, 410-417 (2010).
- Lachkar, A., et al. Identification of self-(in)compatibility S-alleles and new cross-incompatibility groups in Tunisian apricot (Prunus armeniaca L.) cultivars. The Journal of Horticultural Science and Biotechnology. 88, 497-501 (2013).
- Pérez-Pastor, A., Ruiz-Sánchez, M. C., Domingo, R., Torrecillas, A. Growth and phenological stages of Búlida apricot trees in South-East. Agronomie. 24, 93-100 (2004).
- Williams, J. H., Friedman, W. E., Arnold, M. L. Developmental selection within the angiosperm style: using gamete DNA to visualize interspecific pollen competition. Proceedings of the National Academy of Sciences of the United States of America. 96, 9201-9206 (1999).
- Julian, C., Herrero, M., Rodrigo, J. Anther meiosis time is related to winter cold temperatures in apricot (Prunus armeniaca L.). Environmental and Experimental Botany. 100, 20-25 (2014).
- Guerra, M. E., López-Corrales, M., Wünsch, A., Rodrigo, J. Lack of Fruit Set Caused by Ovule Degeneration in Japanese Plum. Journal of the American Society for Horticultural Science. 136 (6), 375-381 (2011).
- Guerra, M. E., Wünsch, A., López-Corrales, M., Rodrigo, J. Flower Emasculation as the Cause for Lack of Fruit Set in Japanese Plum Crosses. Journal of the American Society for Horticultural Science. 135 (6), 556-562 (2010).
- Hormaza, J. I., Pinney, K., Polito, V. S. Correlation in the tolerance to ozone between sporophytes and male gametophytes of several fruit and nut tree species (Rosaceae). Sexual Plant Reproduction. 9, 44-48 (1996).
- Alcaraz, M. L., Hormaza, J. I., Rodrigo, J. Pistil Starch Reserves at Anthesis Correlate with Final Flower Fate in Avocado (Persea americana). PLOS ONE. 8 (10), 78467 (2013).
- Tao, R., et al. Molecular typing of S-alleles through Identification, Characterization and cDNA cloning for S-RNases in Sweet Cherry. Journal of the American Society for Horticultural Science. 124, 224-233 (1999).
- Burgos, L., et al. The self-compatibility trait of the main apricot cultivars and new selections from breeding programmes. Journal of Horticultural Science. 72, 147-154 (1997).
- Hormaza, J. I., Yamane, H., Rodrigo, J. Apricot. Genome Mapping and Molecular Breeding in Plants, Volume 4 Fruits and Nuts. Kole, C. , Springer. New York, NY. 171-187 (2007).
- Benmoussa, H., Ghrab, M., Ben Mimoun, M., Luedeling, E. Chilling and heat requirements for local and foreign almond (Prunus dulcis Mill.) cultivars in a warm Mediterranean location based on 30 years of phenology records. Agricultural and Forest Meteorology. 239, 34-46 (2017).
- Rodrigo, J., Herrero, M., Hormaza, J. I. Pistil traits and flower fate in apricot (Prunus armeniaca). Annals of Applied Biology. 154, 365-375 (2009).
- Williams, R. R. Techniques used in fruit-set experiments. Towards Regulated Cropping. Williams, R. R., Wilson, D. , Grower Books. London. 57-61 (1970).
- Sutherland, B. G., Robbins, T. P., Tobutt, K. R. Primers amplifying a range of Prunus S-alleles. Plant Breeding. 123, 582-584 (2004).
- Murray, M. G., Thompson, W. F. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Research. 8, 4321-4325 (1980).
- Porebski, S., Bailey, L. G., Baum, B. R. Modification of a CTAB DNA Extraction Protocol for Plants Containing High Polysaccharide and Polyphenol Components. Plant Molecular Biology Reporter. 15 (1), 8-15 (1997).
- Rogers, S. O., Bendich, A. J. Extraction of DNA from milligram amounts of fresh, herbarium and mummified plant tissues. Plant Molecular Biology. 5 (2), 69-76 (1985).
- Hormaza, J. I. Molecular characterization and similarity relationships among apricot (Prunus armeniaca L.) genotypes using simple sequence repeats. Theoretical and Applied Genetics. 104, 321-328 (2002).
- Sonneveld, T., Tobutt, K. R., Robbins, T. P. Allele-specific PCR detection of sweet cherry self-incompatibility (S) alleles S1 to S16 using consensus and allele-specific primers. Theoretical and Applied Genetics. 107, 1059-1070 (2003).
- Hegedus, A., Lénárt, J., Halász, J. Sexual incompatibility in Rosaceae fruit tree species: molecular interactions and evolutionary dynamics. Biologia Plantarum. 56 (2), 201-209 (2012).
- Fernández i Martí, A., Gradziel, T. M., Socias i Company, R. Methylation of the Sf locus in almond is associated with S-RNase loss of function. Plant Molecular Biology. 86, 681-689 (2014).
- Company, R. S. i, Kodad, O., Martí, A. F. i, Alonso, J. M. Mutations conferring self-compatibility in Prunus species: From deletions and insertions to epigenetic alterations. Scientia Horticulturae. 192, 125-131 (2015).
- Boskovic, R., Tobutt, K. R. Correlation of stylar ribonuclease zymograms with incompatibility alleles in sweet cherry. Euphytica. 90, 245-250 (1996).
- Cachi, A. M., Wünsch, A. S-genotyping of sweet cherry varieties from Spain and S-locus diversity in Europe. Euphytica. 197 (2), 229-236 (2014).
- Zuriaga, E., et al. An S-locus Independent Pollen Factor Confers Self-Compatibility in "Katy" Apricot. PLoS ONE. 8 (1), 53947 (2013).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}