

Summary
Presentiamo una metodologia per stabilire i requisiti di impollinazione dell'albicocca(Prunus armeniaca L.) che combinano la determinazione dell'auto-in) della microscopia a fluorescenza con l'identificazione del genotipo S mediante l'analisi PCR.
Abstract
L'autoincompatibilità in Rosaceae è determinata da un Gametophytic Self-Incompatibility System (GSI) che è controllato principalmente dal locus multiallelico S. In albicocca, la determinazione delle relazioni di auto-e inter-compatibilità è sempre più importante, poiché il rilascio di un numero importante di nuove cultivar ha portato all'aumento delle cultivar con requisiti di impollinazione sconosciuti. In questo caso, descriviamo una metodologia che combina la determinazione dell'auto-compatibilità mediante impollinazioni manuali e microscopia con l'identificazione del genotipo S mediantel'analisi PCR. Per la determinazione dell'auto-in)compatibilità, i fiori in fase di palloncino da ogni cultivar sono stati raccolti sul campo, impollinati a mano in laboratorio, fissati e macchiati con blu anilina per l'osservazione del comportamento del tubo di polline sotto la microscopia a fluorescenza. Per l'instaurazione di relazioni di incompatibilità tra le cultivar, il DNA di ogni cultivar è stato estratto da foglie giovani e gli alleli Ssono stati identificati dalla PCR. Questo approccio consente di stabilire gruppi di incompatibilità e di chiarire le relazioni di incompatibilità tra le cultivar, che fornisce una preziosa informazione per scegliere i minacciappropriati adatti nella progettazione di nuovi frutteti e per selezionare i genitori appropriati nei programmi di allevamento.
Introduction
L'autoincompatibilità è una strategia di fioritura delle piante per prevenire l'autoimpollinazione e promuovere l'outcrossing1. In Rosaceae, questo meccanismo è determinato da un Gametophytic Self-Incompatibility System (GSI) che è controllato principalmente dal locus multiallelico S2. Nello stile, il gene RNase codifica il determinante tylar S-s,un RNase3, mentre una proteina F-box, che determina il determinante S-pollen,è codificata dal gene SFB 4. L'interazione di autoincompatibilità avviene attraverso l'inibizione della crescita del tubo di polline lungo lo stile impedendo la fecondazione dell'ovulo5,6.
In albicocca, un rinnovamento varietale ha avuto luogo in tutto il mondo negli ultimi due decenni7,8. Questa introduzione di un numero importante di nuove cultivar, provenienti da diversi programmi di allevamento pubblico e privato, ha portato all'aumento delle cultivar di albicocche con requisiti di impollinazione sconosciuti8.
Diverse metodologie sono state utilizzate per determinare i requisiti di impollinazione nell'albicocca. Nel campo, l'auto-(in)compatibilità può essere stabilita da impollinazioni controllate in alberi in gabbia o in fiori evirati e successivamente registrando la percentuale di set di frutta9,10,11,12. Inoltre, in laboratorio sono state effettuate impollinazioni controllate dalla cultura semi-in vivo dei fiori e dall'analisi del comportamento del tubo di polline sotto microscopia a fluorescenza8,13,14,15,16,17. Recentemente, le tecniche molecolari, come l'analisi pcR e il sequenziamento, hanno permesso la caratterizzazione delle relazioni di incompatibilità basate sullo studio dei geni RNase e SFB 18,19. In albicocca, 33 S-alleli sono stati segnalati (S1 a S20, S22 a S30, S52, S53, Sv, Sx), tra cui un allele correlato con autocompatibilità (S)12,18,20,21,22,23,24. Finora, 26 gruppi di incompatibilità sono stati accoltellati in questa specie secondo la S-genotipo8,9,17,25,26,27. S Le cultivar con gli stessi S-allelisono intercompatibili, mentre le cultivar con almeno un allele Adiverso e, di conseguenza, assegnate in diversi gruppi incompatibili, sono intercompatibili.
Per definire i requisiti di impollinazione delle cultivar di albicocca, descriviamo una metodologia che combina la determinazione dell'auto-in)compatibilità mediante microscopia a fluorescenza con l'identificazione del genotipo S-mediante l'analisi PCR nelle cultivar di albicocca. Questo approccio consente di stabilire gruppi di incompatibilità e chiarire le relazioni di incompatibilità tra le cultivar.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. Determinazione dell'auto-in()compatibilità
- Assaggia i fiori nel campo. È necessario raccogliere i fiori in fase di palloncino (Figura 1A), corrispondente alla fase 58 sulla scala BBCH per albicocca28, per evitare l'impollinazione precedente indesiderata.
-
Auto e cross-impollinazioni in laboratorio
- Rimuovere le antre dei fiori in fase di palloncino e metterli su un pezzo di carta per asciugare a temperatura di laboratorio.
- Dopo 24 h, setacciare i grani di polline utilizzando una maglia fine (0,26 mm) (Figura 1B).
- Esalizzare un gruppo di 30 fiori nello stesso stadio palloncino per ogni autoimpollinazione e impollinazione incrociata e posizionare i pisciature sulla schiuma fioraio in acqua a temperatura di laboratorio (Figura 1C).
- Impollinare a mano i pistili con l'aiuto di un pennello con polline da fiori della stessa cultivar 24 h dopo emasculazione. Inoltre, impollinare un altro set di pistili di ogni cultivar con polline da fiori di un impollinante compatibile come controllo (Figura 1D).
- Dopo 72 h, fissare i pistili in una soluzione fissativa di etanolo/acido acetico (3:1) per almeno 24 h a 4 gradicentigradi. Quindi scartare il fissativo e aggiungere 75% etanolo assicurando che i campioni sono completamente sommersi nella soluzione. I campioni possono essere conservati in questa soluzione a 4 gradi centigradi fino all'usodi 8,17,30,31,32.
-
Valutare la vitalità del polline attraverso la germinazione del polline in vitro
- Per preparare il mezzo di germinazione, peso 25 g di saccarosio, 0,075 g di acido borico (H3BO3) e 0,075 g di nitrato di calcio (Ca(NO3)2)33.
- Aggiungere i componenti del mezzo in 250 mL di acqua distillata e sciogliere completamente.
- Solidificare il mezzo aggiungendo 2 g di agarose e mescolare vorticoso.
- Controllare il pH del mezzo utilizzando un misuratore di pH e regolare il valore su 7.0 con la soluzione NaOH o HCl.
- Autoclave la miscela per sterilizzare il mezzo.
- Dopo l'autoclaving, raffreddare il mezzo e distribuirlo nei piatti Petri in una sterile cappa di flusso laminare.
- Spargere i grani di polline delle stesse cultivar utilizzate per le impollinazioni controllate nel mezzo di germinazione del polline solidificato e osservarli al microscopio dopo 24 h6.
NOTA: per sterilizzare il cofano a flusso laminare, pulire la superficie con il 70% di etanolo e accendere la lampada UV durante 10 min. - Conservare i piatti Petri in frigorifero a 4 gradi centigradi fino all'uso.
-
Osservazioni microscopiche
- Lavare i pistilli tre volte per 1 h con acqua distillata e lasciarli nel 5% di solfolito di sodio a 4 gradi centigradi. Dopo 24 h, autoclave a 1 kg /cm2 durante 10 min in solphite di sodio per ammorbidire i tessuti34.
- Posizionare i pistili autoclave su un vetrino di vetro e, con l'aiuto di un bisturi, rimuovere i tricomi intorno all'ovaio per ottenere una migliore visualizzazione dei tubi di polline. Quindi, schiacciare i pistili con un vetro di copertura.
- Preparare lo 0,1% (v/v) una macchia blu anilina: mescolare 0,1 mL di blu anilina in 100 mL di 0,1 N potassi fosfato tribasic (K3PO4). Applicare una goccia di blu anilina sopra i preparati per macchiare le deposizioni di callosi durante la crescita del tubo di polline.
- Osservare i tubi di polline lungo lo stile da un microscopio con epifluorescenza UV utilizzando 340-380 passabanda e 425 filtri longpass.
2. Estrazione del DNA
- Campione 2-3 foglie nel campo in primavera. Si raccomanda di campionare le foglie in giovani stadi poiché il DNA ottenuto è di qualità superiore e livelli più bassi di composti fenolici rispetto alle foglie vecchie.
- Estrarre il DNA genomico seguendo i passaggi descritti in un kit disponibile in commercio (vedere Tabella dei materiali).
- Analizzare la quantità e la qualità delle concentrazioni di DNA utilizzando lo spettrofotometro UV-vis (260 nm).
3. IdentificazioneS-allele
-
Impostazione delle reazioni PCR
- Preparare una diluizione di 50 ng/L nell'acqua distillata di ogni campione di estrazione del DNA.
- Scongelare lentamente i reagenti PCR e tenerli sul ghiaccio. Lasciare la polimerasi del DNA nel congelatore fino a quando necessario.
- Preparare le reazioni di amplificazione utilizzando le diverse combinazioni di primer. Creare il mix di reazione PCR combinando i componenti riportatinella Tabella 1 . Vortex la reazione PCR mescolare bene e distribuire il volume indicato per le diverse combinazioni di primer ad ogni pozzo della piastra PCR. Quindi, aggiungere 1 l della diluizione del DNA in ogni pozzo.
- Posizionare la piastra PCR nel termociclore ed eseguire il programma PCR corrispondente illustrato nella Tabella 1.
- Analizzare i frammenti amplificati. Ci sono principalmente due modi diversi per analizzare i frammenti amplificati PCR: elettroforesi capillare (CE) con primer fluorescenti etichettati o come esorti di elettroforesi gel agarose con primer non etichettati.
-
Elettroforesi capillare
- Per preparare il buffer di carico, mescolare 35 L di formamide deionizzato con 0,45 l di standard di dimensionamento etichettato. Vorticare il reagente per mescolare bene, e poi erogare 35,5 l nel pozzo della piastra del lettore.
- Aggiungere 1 ll l del prodotto PCR nel pozzo. Inoltre, aggiungere una goccia di olio minerale per evitare l'evaporazione dell'acqua.
- Preparare la piastra di separazione aggiungendo il buffer di separazione.
- Utilizzare il software commerciale incluso nell'analizzatore genetico (vedere Tabella dei materiali). Creare una nuova piastra di esempio e salvare i nomi di esempio per tutti i pozzetto sulla piastra.
- Selezionare il metodo di analisi. In questo caso, denaturare i campioni a 90 gradi centigradi per 120 s, iniettare a 2,0 kV per 30 s e separarli a 6,0 kV per 35 min.
- Inserire le due piastre nell'analizzatore genetico. Riempire la matrice capillare con acqua distillata.
- Caricare il gel brevettato in poliacrilammide lineare (LPA). Infine, fare clic su Esegui.
-
Gel Elettroforesi
- Preparare un 1% gel di agarose aggiungendo 1,5 g di agarose di biologia molecolare in 150 mL di 1x TAE (Tris-acetate-EDTA) buffer di elettroforesi (40 mM, 20 mM di acido acetico e 1 mM EDTA a pH 8.0). Sciogliere l'agarose mediante riscaldamento a microonde per 2-3 min.
- Per visualizzare il DNA, aggiungere 4 -L di una macchia acida nucleica (vedi Tabella dei materiali) e mescolare delicatamente.
- Aggiungere un pettine di gel, con pozzi sufficienti per scale, controlli e campioni, in un vassoio di gel. Quindi, versare lentamente il mix nel mezzo del vassoio di gel ed evitare le bolle.
- Lasciare raffreddare il gel per 30-45 min a temperatura ambiente fino a quando il gel si è completamente solidificato. Introdurre il gel nella camera di elettroforesi, rimuovere il pettine di gel e riempire la camera con abbastanza 1x tampone TAE per coprire il gel.
NOTA: Controllare il posizionamento del gel. I pozze devono essere posizionati vicino al polo negativo poiché il DNA caricato negativamente migra verso il catodo. - Aggiungere 5 l di buffer di carico (0,1% (v/v) di bromofenolo blu) ai prodotti PCR e mescolare bene.
- Per stimare le dimensioni delle bande, caricare 5 - L di DNA scala peso molecolare (vedere Tabella dei materiali).
- Caricare i campioni nei pozze aggiuntivi del gel.
- Una volta caricati tutti i campioni e la scala di peso molecolare del DNA, eseguire il gel a 90 V per 1-1,5 h, fino a quando la linea di tintura blu è approssimativamente al 75% della lunghezza del gel.
- Visualizza le bande in un transilluminatore per gli acidi nucleici.
-
Elettroforesi capillare
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Gli studi di impollinazione in albicocca richiedono l'uso di fiori nella fase tarda del palloncino un giorno prima dell'analisi (Figura 1A). Questa fase è considerata la più favorevole sia per la raccolta del polline che per la raccolta del pistilo, poiché le strutture floreali sono quasi mature, ma l'inestesia non si è ancora verificata. Questo impedisce l'interferenza di polline indesiderato, non solo di polline dallo stesso fiore, ma anche da altri fiori, poiché i petali chiusi impediscono l'arrivo di insetti che trasportano polline esterno. I grani di polline vengono facilmente setacciati attraverso una maglia fine (Figura 1B) da antere dehiscie precedentemente poste su un pezzo di carta per 24 h a temperatura ambiente o con un leggero calore supplementare. Allo stesso modo, i pistili sono ottenuti dai fiori in fase di palloncino dopo la rimozione di petali, sepali e stami con l'aiuto di pinzette o unghie (Figura 1C). I pistili possono essere auto-e cross-impollinato con un pennello fine (Figura 1D).
I fiori ermatodici dell'albicocca hanno cinque sepali rossi scuri, cinque petali bianchi (Figura 1A), un unico pistillo (Figura 2A) e 25-30 stami. Il pistilè ha tre strutture principali: stigma, stile e ovaia. L'ovaio ha due ovuli, e la fecondazione di almeno uno di loro è necessaria per l'impostazione della frutta. Durante l'impollinazione, gli insetti, principalmente le api, trasferiscono i grani di polline allo stigma (Figura 1A), dove germinano (Figura 2B) entro 24 h dopo l'impollinazione. Un tubo di polline è prodotto da ogni chicco di polline germinante, che cresce attraverso le strutture del pistilo per raggiungere l'ovaio dopo 3-4 giorni e fertilizzare uno dei due ovuli dopo circa 7 giorni. Nelle cultivar auto-incompatibili in cui l'allele S del grano di polline è lo stesso di uno dei due pistili, tubo di polline smette di crescere allo stile superiore, impedendo la fecondazione (Figura 2C). Tuttavia, i tubi di polline di una cultivar compatibile, con un diverso allele S, possono crescere attraverso lo stile (Figura 2D), raggiungere l'ovaio (Figura 2E) e fertilizzare uno dei due ovuli.
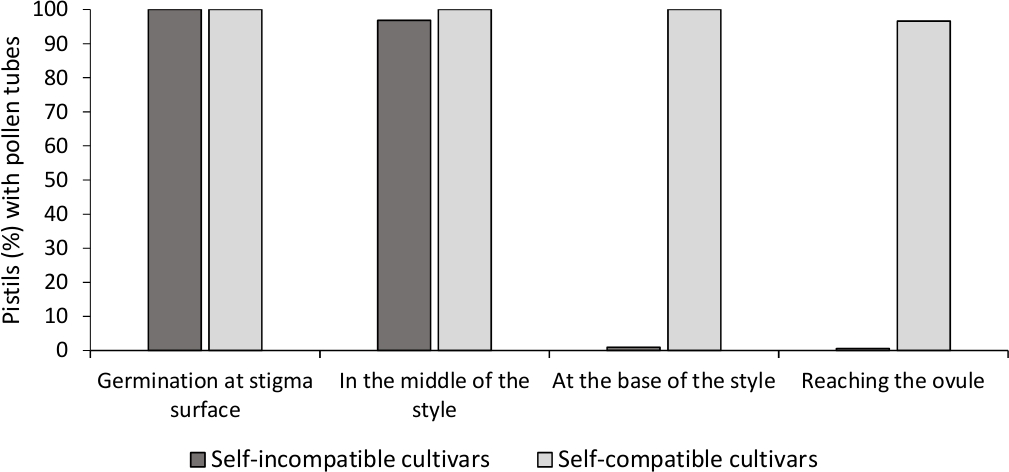
L'analisi della germinazione del polline in vitro ha mostrato una buona vitalità del polline in tutte le cultivar analizzate qui, poiché la maggior parte dei tubi di polline erano più lunghi della lunghezza del chicco di polline dopo 24 h nel mezzo di coltura. I grani di polline germinato sono stati osservati sulla superficie dello stigma (Figura 2B) in pistili da tutte le impollinazioni, indicando un'adeguata impollinazione (Figura 3).
Per determinare l'auto-in)compatibilità per ogni cultivar, il comportamento del tubo di polline in auto e cross-pollinazioni fatto in condizioni controllate dal laboratorio è stato osservato sotto microscopia a fluorescenza. La crescita del tubo di polline è stata registrata lungo lo stile in tutti i pistili esaminati. Le cultivar sono state considerate auto-incompatibili quando la crescita del tubo di polline è stata arrestata lungo lo stile nella maggior parte dei pistils auto-impollinati (Figura 2C, Figura 3) e autocompatibile quando almeno un tubo di polline ha raggiunto la base dello stile nella maggior parte dei pistili esaminati ( Figura2E, Figura 3).
Lo studio dell'analisi S-locus da PCR ha permesso di caratterizzare il genotipo S diogni cultivar. In primo luogo, gli alleli Ssono stati identificati dall'amplificazione del primo intron S-RNase utilizzando i primer SRc-F/SRc-R (tabella 2). La dimensione dei frammenti amplificati è stata analizzata dall'elettroforesi capillare (Figura 4A) ed è stata utilizzata per classificare i genotipi analizzati nel gruppo di incompatibilità corrispondente (I.G.) (Tabella 3).
Alcune coppie di alleli, come S1 e S7 o S6 e S9, hanno mostrato dimensioni di frammenti simili per il primo intron. Pertanto, la differenziazione di questi alleli è stata effettuata amplificando una regione del secondo intron del RNase con i primer Pru-C2/PruC4R, SHLM1/SHLM2 e SHLM3/SHLM4(tabella 2). La combinazione di primer PruC2/PruC4R è stata utilizzata per distinguere tra S6 e S9. Per S6è stato amplificato un frammento di 1300 bp, mentre è stato osservato un frammento di circa 700 bp per l'allele S9 ( Figura4B, tabella 3). I primer specifici SHLM1/SHLM2 e SHLM3/SHLM4 amplificavano un frammento di circa 650 bp nell'allele S1 e di 413 bp nell'allele S7 ( Figura4C, Tabella 3).
I primer AprFBC8-(F/R) che amplificano le regioni variabili V2 e HVb del gene SFB sono stati utilizzati per distinguere gli alleli Sc e S8 poiché entrambi gli alleli mostrano la stessa sequenza di RNase. L'allele S8 ha mostrato un frammento di PCR di circa 150 bp, mentre un frammento di 500 bp corrispondeva all'allele Sc ( Figura4D, Tabella 3). Una volta che il -genotipo Sè stato determinato per tutte le cultivar, cultivar auto-incompatibili sono state assegnate ai corrispondenti gruppi di incompatibilità in base ai loro S-alleli( Tabella3).
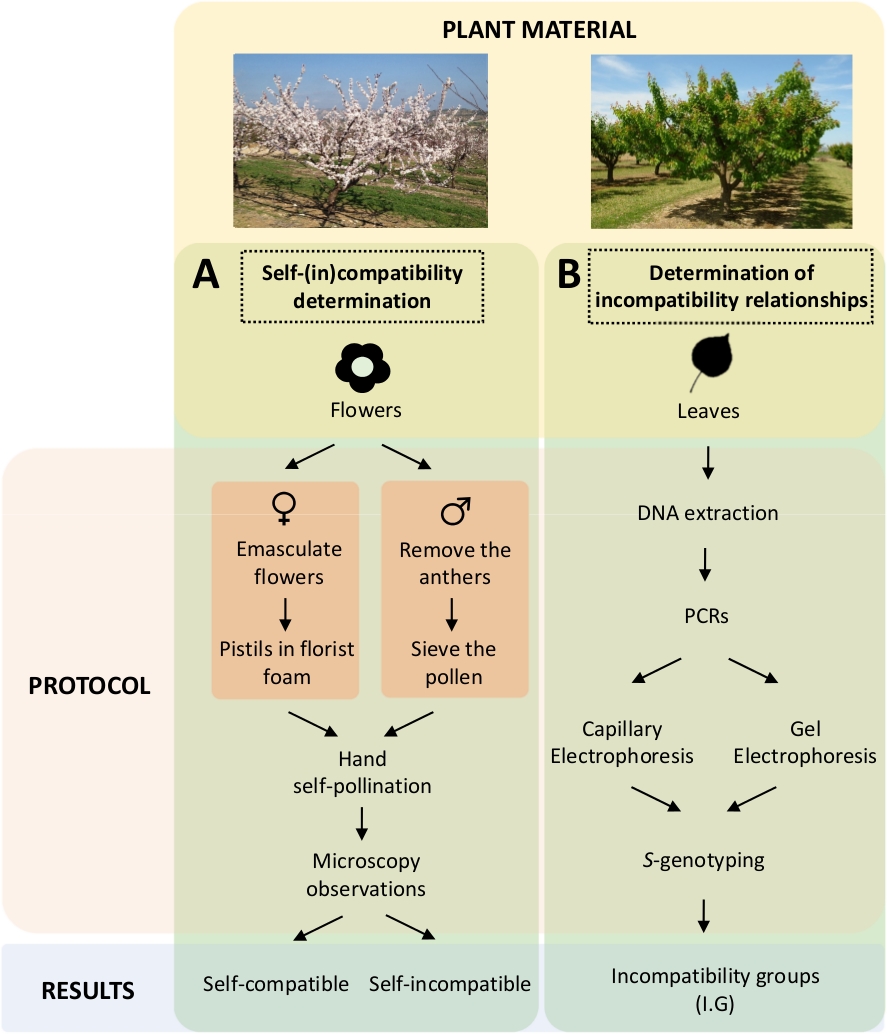
Questo approccio richiede di determinare l'auto(in)compatibilità di ogni cultivar mediante auto-impollinazioni controllate in laboratorio (Figura 5A) in concomitanza con la caratterizzazione del genotipo Sdall'analisigenetica( Figura 5B). Di conseguenza, è possibile determinare i requisiti di impollinazione di ogni cultivar e le relazioni di incompatibilità tra le cultivar di albicocca.

come illustrato nella Figura 1. Sperimentale istituito per la determinazione dell'auto-in(in)compatibilità in albicocca.
(A) Fiori in fase a palloncino (frecce nere) nel campo. (B) Setaccio di grani di polline utilizzando una maglia fine. (C) Pistils posto sulla schiuma fiorista in acqua. (D) impollinazione a mano dei pistili con l'aiuto di un pennello. Fare clic qui per visualizzare una versione più grande di questa figura.

come illustrato nella Figura 2. Rappresentazione diagrammatica delle relazioni di incompatibilità gametophytic nei fiori di albicocca.
(A) In Gametophytic Self-Incompatibility (GSI), i grani di polline compatibili e incompatibili germinano sullo stigma. Il granello di polline trasporta uno dei due alleli Adel genotipo originale, in questo caso S1 o S2. Se la S-allele del grano di polline corrisponde a uno dei due -alleli Sdel pistil, in questo caso S1S3, la crescita del tubo di polline è inibita nella parte superiore di un terzo dello stile. (B) Germinazione dei grani di polline sulla superficie dello stigma. (C) Tubo di polline arrestato nello stile indicando un comportamento incompatibile. (D) Tubi di polline che crescono lungo lo stile. (E) Tubi di polline alla base dello stile che indicano un comportamento compatibile. Barre della scala, 100 m. Fare clic qui per visualizzare una versione più grande di questa figura.

come illustrato nella figura 3. Risultati rappresentativi della germinazione del polline e della crescita dei tubi di polline attraverso lo stile di cultivar auto-compatibili e auto-incompatibili dopo le autoiscrizioni.
Percentuale di pistili con grani di polline che germinano sulla superficie dello stigma, con tubi di polline a metà dello stile, alla base dello stile, e raggiungendo l'ovulo. Fare clic qui per visualizzare una versione più grande di questa figura.

come illustrato nella Figura 4. Amplificazione del frammento pcR utilizzando cinque combinazioni di coppie di primer per l'identificazione degli alleli S.
(A) Uscita analizzatore genetico per i primer SRc-(F/R) che mostra la dimensione dei due frammenti amplificati della regione RNase per la prima introne corrispondente agli alleli S. (B) amplificazione PCR utilizzando le primer PruC2/PruC4R per l'identificazione degli alleli S6 e S9. (C) prodotti PCR ottenuti utilizzando le primer specifiche SHLM1 e SHLM2 per la differenziazione dell'allele S1 e SHLM3 e SHLM4 per distinguere l'allele S7. (D) amplificazione PCR con le prime rinportaA-(F/R) per l'identificazione degli alleli Sc e S8. MI: 1 kb DNA Ladder. MII: 100 bp DNA Ladder. Fare clic qui per visualizzare una versione più grande di questa figura.

come illustrato nella Figura 5. Schema del progetto sperimentale per chiarire le relazioni di compatibilità auto-in nelle cultivar di albicocca.
(A) Flusso di lavoro di determinazione dell'auto-in)compatibilità mediante impollinazioni controllate in laboratorio. (B) Flusso di lavoro dell'identificazione delle S-allelemediante approcci molecolari. Fare clic qui per visualizzare una versione più grande di questa figura.
| Miscela Master PCR | Condizioni termociclista | ||||||
| Componenti | Concentrazione finale | 15 - Reazione L | Passaggio del ciclo | Temperatura | Ore | Cicli | |
| 10x NH4 Buffer di reazione | 10x (10x) | 1,5 ll | Denaturazione iniziale | 94 gradi centigradi | 3 min | 1 | |
| Soluzione MgCl2 da 50 mM | 25 mM | 1,2 lL | Denaturazione | 94 gradi centigradi | 1 min | 35 | |
| 100 mM dNTP | 2,5 mM | 0,6 l | Ricottura | 55 gradi centigradi | 1 min | ||
| Primer SRc-F | 10 M | 0,6 l | Estensione | 72 gradi centigradi | 3 min | ||
| Primer SRc-R | 10 M | 0,6 l | Estensione finale | 72 gradi centigradi | 5 min | 1 | |
| 500 U Taq DNA Polymerasi | 0,5 U | 0,2 l l | 4 gradi centigradi | Tenere | |||
| H2O | 8.3 LL | ||||||
| Componenti | Concentrazione finale | Reazione a 25 l | Passaggio del ciclo | Temperatura | Ore | Cicli | |
| Buffer PCR 10x | 10x (10x) | 2,5 l l | Denaturazione iniziale | 94 gradi centigradi | 2 min | 1 | |
| 5x soluzione Q | 5x (in stato di | 5 ll | Denaturazione | 94 gradi centigradi | 10 s | 10 | |
| 100 mM dNTP | 2,5 mM | 0,5 l l | Ricottura | 55 gradi centigradi | 2 min | ||
| Primer PruC2 | 10 M | 0,2 l l | Estensione | 68 gradi centigradi | 2 min | ||
| Primer C4R | 10 M | 0,2 l l | Denaturazione | 94 gradi centigradi | 10 s | 25 | |
| 250 U Taq DNA Polymerasi | 10 U | 0,13 l | Ricottura | 58 gradi centigradi | 2 min | ||
| H2O | 15,5 ll | Estensione | 68 gradi centigradi | 2 min | |||
| Estensione finale | 72 gradi centigradi | 5 min | 1 | ||||
| 4 gradi centigradi | Tenere | ||||||
| - con 10 s aggiunto ogni ciclo al passaggio di estensione 68 % C. | |||||||
| Componenti | Concentrazione finale | Reazione a 25 l | Passaggio del ciclo | Temperatura | Ore | Cicli | |
| Buffer PCR 10x | 10x (10x) | 2,5 l l | Denaturazione iniziale | 94 gradi centigradi | 2 min | 1 | |
| 5x soluzione Q | 5x (in stato di | 5 ll | Denaturazione | 94 gradi centigradi | 30 s | 35 | |
| 100 mM dNTP | 2,5 mM | 0,5 l l | Ricottura | 62 gradi centigradi | 1,5 min | ||
| Primer SHLM1 | 10 M | 0,2 l l | Estensione | 72 gradi centigradi | 2 min | ||
| Primer SHLM2 | 10 M | 0,2 l l | Estensione finale | 72 gradi centigradi | 5 min | 1 | |
| 250 U Taq DNA Polymerasi | 10 U | 0,13 l | 4 gradi centigradi | Tenere | |||
| H2O | 15,5 ll | ||||||
| Componenti | Concentrazione finale | Reazione a 20 l | Passaggio del ciclo | Temperatura | Ore | Cicli | |
| 5x Buffer PCR | 5x (in stato di | 4 ll | Denaturazione iniziale | 98 gradi centigradi | 30 s | 1 | |
| dNTP | 2,5 mM | 1,6 l | Denaturazione | 98 gradi centigradi | 10 s | 35 | |
| Primer SHLM3 | 10 M | 1 ll | Ricottura | 51 gradi centigradi | 30 s | ||
| Primer SHLM4 | 10 M | 1 ll | Estensione | 72 gradi centigradi | 1 min | ||
| 100 U DNA Polymerasi | 5 U | 0,2 l l | Estensione finale | 72 gradi centigradi | 5 min | 1 | |
| H2O | 12,4 l l | 4 gradi centigradi | Tenere | ||||
| Componenti | Concentrazione finale | Reazione a 25 l | Passaggio del ciclo | Temperatura | Ore | Cicli | |
| Buffer PCR 10x | 10x (10x) | 2,5 l l | Denaturazione iniziale | 94 gradi centigradi | 2 min | 1 | |
| 100 mM dNTP | 2,5 mM | 2 ll | Denaturazione | 94 gradi centigradi | 30 s | 35 | |
| Primer FBC8-F | 10 M | 1 ll | Ricottura | 55 gradi centigradi | 1,5 min | ||
| Primer FBC8-R | 10 M | 1 ll | Estensione | 72 gradi centigradi | 2 min | ||
| 250 U Taq DNA Polymerasi | 10 U | 0,125 l | Estensione finale | 72 gradi centigradi | 5 min | 1 | |
| H2O | 17,4 L | 4 gradi centigradi | Tenere | ||||
Tabella 1. Condizioni di reazione e ciclismo per diverse combinazioni di primer utilizzate in questo protocollo.
| Primer | Sequenza | Riferimento |
| SRc-F | 5'-CTCGCTTTCTTTGTTCTCTGC-3' | 18 |
| SRc-R | 5'-GGCCATTGTTGCACCTG-3' | 18 |
| Pru-C2 | 5'-CTTTGGCCAAGTAATTATTCAAACC-3' | 35 |
| Pru-C4R | 5'-GGATGTGGTACGATTGAAGCG-3' | 35 |
| SHLM1-F | 5'-GGTGGAGGTGATAGGTAGCC-3' | 17 |
| SHLM2-R | 5'-GGCTGCATAAGGAGGCTGTAG3' | 17 |
| SHLM3-F | 5'-TATATCTTACTCTCTTTGGC-3' | 17 |
| SHLM4-R | 5'-CACTATGATAATGTGTATG-3' | 17 |
| AprFBC8-F | 5'-CATGGAAAAAGCTGACTTATGG-3' | 26 |
| AprFBC8-R | 5'-GCCTCTAATGTCATCTCTCTCTTAG-3' | 26 |
Tabella 2. Primers utilizzati in questo protocollo, sequenza e riferimento per la caratterizzazione S-genotipo in Prunus armeniaca.
| Cultivar | SRc-(F/R) (bp) | PruC2/PruC4R (bp) | SHLM1/SHLM2 (bp) | SHLM3/SHLM4 (bp) | AprFBC8-(F/R) (bp) | S-Genotipo | Gruppo di incompatibilità (I.G) |
| Meraviglia Cot8 | 420, 420 | 749, 1386 | S6S9 | Viii | |||
| Cot magico8 | 334, 420 | 749 | S2S9 | Xx | |||
| Colpo d'oro8 | 334, 420 | 749 | S2S9 | - | |||
| T06917 | 334, 408 | 650 | S1S2 | Ho | |||
| T12017 | 334, 408 | 650 | S1S2 | - | |||
| C-6 | 334, 408 | 413 | S2S7 | Iv | |||
| Cooper Cot8 | 274, 408 | 650 | S1S3 | XVIII | |||
| Albicocia | 358, 358 | 500 | Scsc | - | |||
| Bergecot8 | 334, 358 | 500 | S2Sc | - | |||
| Blush Primavera8 | 274, 358 | 150 | S3S8 | XXI |
Tabella 3. S-genotipizzazione di cultivar di albicocca con cinque coppie di primer utilizzate in questo protocollo e assegnazione di gruppo di incompatibilità. Le diverse dimensioni dei prodotti di Sreazione della catena polimerasi di S-alleles amplificati utilizzando SRc-(F/R), PruC2/PruC4R, SHLM1/ SHLM2, SHLM3/SHLM4 e AprFBC8-(F/R) primer sono mostrati nella tabella.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Tradizionalmente, la maggior parte delle cultivar europee di albicocche commerciali erano auto-compatibili36. Tuttavia, l'uso di cultivar auto-incompatibili nordamericane come genitori nei programmi di allevamento negli ultimi decenni ha portato al rilascio di un numero crescente di nuove cultivar auto-incompatibili con requisiti di impollinazione sconosciuti7,8,37. Pertanto, la determinazione delle relazioni di auto-e inter-compatibilità nelle cultivar di albicocca è sempre più importante. Questo è accentuato in quelle zone in cui il raffreddamento invernale sta diminuendo, dal momento che le variazioni elevate di anno in anno nel tempo della fioritura impediscono la coincidenza nella fioritura delle cultivar e dei loro pollinizzatori in molti casi, soprattutto nelle cultivar con elevati requisiti di raffreddamento38. La metodologia qui descritta, che combina l'impollinazione manuale, la microscopia e le analisi genetiche, è stata molto utile per determinare l'auto(in)compatibilità di ogni cultivar e per stabilire le sue potenziali cultivar impollinatori.
I requisiti di impollinazione possono essere determinati attraverso esperimenti di controllo sul campo nelle condizioni del frutteto11,39. Tuttavia, l'esposizione a fattori esterni, tra cui condizioni meteorologiche avverse, può causare il fallimento dell'impollinazione10, il che può comportare diagnosi errate di autoincompatibilità. La metodologia descritta di seguito consente di valutare l'auto-compatibilità in modo più accurato mediante osservazioni microscopiche di fiori impollinati a mano in condizioni controllate da laboratorio, evitando l'influenza ambientale. Inoltre, questo approccio consente di analizzare un maggior numero di cultivar all'anno, poiché è richiesto solo un piccolo numero di fiori al posto di diversi alberi adulti per ogni cultivar che sono necessari negli esperimenti sul campo40.
Le relazioni di incompatibilità possono essere stabilite combinando impollinazioni a mano e microscopia14. Tuttavia, le impollinazioni possono essere eseguite solo per un breve periodo durante la stagione della fioritura in primavera, e gli alberi adulti vicino al laboratorio sono necessari, poiché la durata della vita dei fiori raccolti è molto breve. Così, il numero di relazioni di incompatibilità che possono essere analizzate da impollinazioni a mano controllate in ogni stagione è molto basso. La caratterizzazione dei geni codificati dalla S-locusha permesso lo sviluppo di metodi basati su PCR per la genotipizzazione di S-allele18,41. Questo approccio Saccelera l'identificazione Di-allele in quanto non richiede fiori e gli esperimenti possono essere effettuati con qualsiasi tessuto vegetativo42. Questo si estende il periodo durante il quale il materiale vegetale, di solito foglie giovani, può essere raccolto43. Inoltre, le foglie possono essere lofilizzate o congelate, in modo che l'analisi possa essere fatta in qualsiasi momento dell'anno, a differenza delle impollinazioni che possono essere fatte solo su fiori freschi durante la stagione della fioritura44. Un ulteriore vantaggio è che le foglie possono essere raccolte da giovani alberi anche prima di entrare in età di fioritura, facilitando la raccolta di campioni e l'ottenimento precoce dei risultati45.
L'analisi genetica consente una migliore differenziazione degli alleli di autoincompatibilità in quanto fornisce risultati precisi delle dimensioni dei frammenti amplificati21,46. Ad oggi, trentatré alleli Asono stati identificati nell'albicocca12,18,20,2121,22,23,23,24, che ha permesso di stabilire 36 gruppi di incompatibilità basati su S-genotipo8,9,17,25,26,27. D'altra parte, uno svantaggio di questa metodologia è che diversi alleli nella stessa dimensione dell'intervallo o mutazioni possono essere erroneamente identificati come lo stesso allele. Pertanto, gli alleli Sc e S8 sono identici per la sequenza RNase, ma un inserimento di 358 bp si trova nel gene SFB di Sc19. Allo stesso modo, la prima regione intron degli alleli S1 e S7 sono identici e sono indistinguibili utilizzando i primer SRc-F/SRc-R. Inoltre, sono state trovate diverse omologie, come S6 e S528 o S20 e S55, e S7, S13 (EF062341) e S4617, perché alcuni di questi alleli sono stati parzialmente sequenziati o da guasti durante l'amplificazione PCR e, di conseguenza, sono necessari ulteriori lavori per distinguerli correttamente.
L'analisi PCR e il sequenziamento S-RNase sono adeguati per stabilire relazioni di incompatibilità attraverso l'identificazione degli Alleli Se l'assegnazione delle cultivar nel corrispondente Incompatibility Group8,17,26,27. Tuttavia, questa metodologia ha la limitazione di impedire la determinazione dell'auto-compatibilità per particolari cultivar di albicocche. L'autocompatibilità (SC) è stata associata a particolari alleli Sin altre specie di Prunus 47, come mandorla (Sf)48,49 o ciliegia dolce (S4')50,51. Tuttavia, in albicocca, l'allele Sc, che è stato associato a SC21, può essere erroneamente identificato come S8, un allele auto-incompatibile19,22, e possibili mutazioni non legate al locus S, come il M-locus12,52, conferendo SC sono stati identificati. Recentemente, Mil M-locus è stato genotipato utilizzando marcatori SSR12. Pertanto, l'identificazione genetica di SC per i genotipi albicocche ha bisogno di ulteriori ricerche e, al fine di evitare errori dovuti a fattori non legati al locus S, in questo lavoro la caratterizzazione dell'auto-in)compatibilità è stata determinata anche fenotipizzando il comportamento dei tubi di polline attraverso il pistillo dei fiori autoimpollinati.
La metodologia qui descritta combinando la determinazione dell'auto-in) compatibilità mediante impollinazioni manuali in condizioni di laboratorio con la successiva osservazione del comportamento dei tubi di polline nel pistillo delle auto-pollini a controllo nell'ambito della microscopia a fluorescenza e l'identificazione del genotipo Smediante l'analisi PCR stabilendo i requisiti di impollinazione delle cultivar di albicocca. Questo fornisce una preziosa informazione per i coltivatori e gli allevatori, in quanto permette di stabilire le relazioni di incompatibilità tra le cultivar di scegliere impollinatori adatti nella progettazione di nuovi frutteti e di selezionare genitori appropriati per progettare nuove croci nei programmi di allevamento di albicocche.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Gli autori non hanno nulla da rivelare.
Acknowledgments
Questa ricerca è stata finanziata dal ministro de Ciencia, Innovaciàn y Universidades-European Regional Development Fund, European Union (AGL2016-77267-R e AGL2015-74071-JIN); Instituto Nacional de Investigaciàn y Tecnolog à Agraria y Alimentaria (RFP2015-00015-00, RTA2017-00003-00); Gobierno de Aragàn-European Social Fund, European Union (Grupo Consolidado A12_17R), Fundaciàn Biodiversidad e Agroseguro S.A.
Materials
| Name | Company | Catalog Number | Comments |
| Agarose D1 Low EEO | Conda | 8010.22 | |
| BIOTAQ DNA Polymerase kit | Bioline | BIO-21060 | |
| Bright field microscope | Leica Microsystems | DM2500 | |
| CEQ System Software | Beckman Coulter | ||
| DNeasy Plant Mini Kit | QIAGEN | 69106 | |
| dNTP Set, 4 x 25 µmol | Bioline | BIO-39025 | |
| GenomeLab DNA Size Standard Kit - 400 | Beckman Coulter | 608098 | |
| GenomeLab GeXP Genetic Analysis System | Beckman Coulter | ||
| GenomeLab Separation Buffer | Beckman Coulter | 608012 | |
| GenomeLab Separation Gel LPA-1 | Beckman Coulter | 391438 | |
| HyperLadder 100bp | Bioline | BIO-33029 | |
| HyperLadder 1kb | Bioline | BIO-33025 | |
| Image Analysis System | Leica Microsystems | ||
| Molecular Imager VersaDoc MP 4000 system | Bio-Rad | 170-8640 | |
| NanoDrop One Spectrophotometer | Thermo Fisher Scientific | 13-400-518 | |
| pH-Meter BASIC 20 | Crison | ||
| Phusion High-Fidelity PCR Kit | Thermo Fisher Scientific | F553S | |
| Power Pack P 25 T | Biometra | ||
| Primer Forward | Isogen Life Science | ||
| Primer Reverse | Isogen Life Science | ||
| Quantity One Software | Bio-Rad | ||
| Stereoscopic microscope | Leica Microsystems | MZ-16 | |
| Sub-Cell GT | Bio-Rad | ||
| SYBR Safe DNA Gel Stain | Thermo Fisher Scientific | S33102 | |
| T100 Thermal Cycler | Bio-Rad | 1861096 | |
| Taq DNA Polymerase | QIAGEN | 201203 | |
| Vertical Stand Autoclave | JP Selecta |
References
- Silva, N. F., Goring, D. R. Mechanisms of self-incompatibility in flowering plants. Cellular and Molecular Life Sciences. 58, 1988-2007 (2001).
- Charlesworth, D., Vekemans, X., Castric, V., Glémin, S. Plant self-incompatibility systems: A molecular evolutionary perspective. New phytologist. 168, 61-69 (2005).
- Tao, R., et al. Identification of stylar RNases associated with gametophytic self-incompatibility in almond (Prunus dulcis). Plant and Cell Physiology. 38, 304-311 (1997).
- Ushijima, K., et al. Structural and transcriptional analysis of the self-incompatibility locus of almond: Identification of a pollen-expressed F-box gene with haplotype-specific polymorphism. The Plant cell. 15, 771-781 (2003).
- Bedinger, P. A., Broz, A. K., Tovar-Mendez, A., McClure, B. Pollen-Pistil Interactions and Their Role in Mate Selection. Plant Physiology. 173, 79-90 (2017).
- Guerra, M. E., Rodrigo, J. Japanese plum pollination: A review. Scientia Horticulturae. 197, 674-686 (2015).
- Zhebentyayeva, T., Ledbetter, C., Burgos, L., Llacer, G. Apricot. Fruit Breeding. Badenes, M. L., Byrne, D. , Springer. Boston, MA. 415-458 (2012).
- Herrera, S., Lora, J., Hormaza, J. I., Herrero, M., Rodrigo, J. Optimizing Production in the New Generation of Apricot Cultivars: Self-incompatibility, S-RNase Allele Identification, and Incompatibility Group Assignment. Frontiers in Plant Science. 9, 527 (2018).
- Egea, J., Burgos, L. Detecting Cross-incompatibility of Three North American Apricot Cultivars and Establishing the First Incompatibility Group in Apricot. Journal of the American Society for Horticultural Science. 121, 1002-1005 (1996).
- Rodrigo, J., Herrero, M. Effects of pre-blossom temperatures on flower development and fruit set in apricot. Scientia Horticulturae. 92, 125-135 (2002).
- Julian, C., Herrero, M., Rodrigo, J. Flower bud differentiation and development in fruiting and non-fruiting shoots in relation to fruit set in apricot (Prunus armeniaca). Trees. 24, 833-841 (2010).
- Muñoz-Sanz, J. V., Zuriaga, E., López, I., Badenes, M. L., Romero, C. Self-(in)compatibility in apricot germplasm is controlled by two major loci, S and M. BMC Plant Biology. 17, 82 (2017).
- Burgos, L., Berenguer, T., Egea, J. Self- and Cross-compatibility among Apricot Cultivars. HortScience. 28, 148-150 (1993).
- Rodrigo, J., Herrero, M. Evaluation of pollination as the cause of erratic fruit set in apricot "Moniqui". Journal of Horticultural Science. 71, 801-805 (1996).
- Milatović, D., Nikolić, D., Krška, B. Testing of self-(in)compatibility in apricot cultivars from European breeding programmes. Horticultural Science. 40 (2), 65-71 (2013).
- Milatović, D., Nikolić, D., Fotirić-Aksić, M., Radović, A. Testing of self-(in)compatibility in apricot cultivars using fluorescence microscopy. Acta Scientiarum Polonorum, Hortorum Cultus. 12 (6), 103-113 (2013).
- Herrera, S., Rodrigo, J., Hormaza, J. I., Lora, J. Identification of Self-Incompatibility Alleles by Specific PCR Analysis and S-RNase Sequencing in Apricot. Int J Mol Sci. 19, 3612 (2018).
- Romero, C., et al. Analysis of the S-locus structure in Prunus armeniaca L. Identification of S-haplotype specific S-RNase and F-box genes. Plant Molecular Biology. 56, 145-157 (2004).
- Halász, J., Pedryc, A., Hegedus, A. Origin and dissemination of the pollen-part mutated SC haplotype which confers self-compatibility in apricot (Prunus armeniaca). New Phytologist. 176, 792-803 (2007).
- Halász, J., Hegedus, A., Hermán, R., Stefanovits-Bányai, É, Pedryc, A. New self-incompatibility alleles in apricot (Prunus armeniaca L.) revealed by stylar ribonuclease assay and S-PCR analysis. Euphytica. 145, 57-66 (2005).
- Vilanova, S., Romero, C., Llacer, G., Badenes, M. L., Burgos, L. Identification of Self-(in)compatibility Alleles in Apricot by PCR and Sequence Analysis. Journal of the American Society for Horticultural Science. 130, 893-898 (2005).
- Feng, J., et al. Detection and transcript expression of S-RNase gene associated with self-incompatibility in apricot (Prunus armeniaca L.). Molecular Biology Reports. 33, 215-221 (2006).
- Zhang, L., et al. Identification of self-incompatibility (S-) genotypes of Chinese apricot cultivars. Euphytica. 160, 241-248 (2008).
- Wu, J., et al. Identification of S-haplotype-specific S-RNase and SFB alleles in native Chinese apricot (Prunus armeniaca L). Journal of Horticultural Science and Biotechnology. 84, 645-652 (2009).
- Szabó, Z., Nyéki, J. Blossoming, fructification and combination of apricot varieties. Acta Horticulturae. 293, 295-302 (1991).
- Halász, J., Pedryc, A., Ercisli, S., Yilmaz, K. U., Hegedűs, A. S-genotyping supports the genetic relationships between Turkish and Hungarian apricot germplasm. Journal of the American Society for Horticultural Science. 135, 410-417 (2010).
- Lachkar, A., et al. Identification of self-(in)compatibility S-alleles and new cross-incompatibility groups in Tunisian apricot (Prunus armeniaca L.) cultivars. The Journal of Horticultural Science and Biotechnology. 88, 497-501 (2013).
- Pérez-Pastor, A., Ruiz-Sánchez, M. C., Domingo, R., Torrecillas, A. Growth and phenological stages of Búlida apricot trees in South-East. Agronomie. 24, 93-100 (2004).
- Williams, J. H., Friedman, W. E., Arnold, M. L. Developmental selection within the angiosperm style: using gamete DNA to visualize interspecific pollen competition. Proceedings of the National Academy of Sciences of the United States of America. 96, 9201-9206 (1999).
- Julian, C., Herrero, M., Rodrigo, J. Anther meiosis time is related to winter cold temperatures in apricot (Prunus armeniaca L.). Environmental and Experimental Botany. 100, 20-25 (2014).
- Guerra, M. E., López-Corrales, M., Wünsch, A., Rodrigo, J. Lack of Fruit Set Caused by Ovule Degeneration in Japanese Plum. Journal of the American Society for Horticultural Science. 136 (6), 375-381 (2011).
- Guerra, M. E., Wünsch, A., López-Corrales, M., Rodrigo, J. Flower Emasculation as the Cause for Lack of Fruit Set in Japanese Plum Crosses. Journal of the American Society for Horticultural Science. 135 (6), 556-562 (2010).
- Hormaza, J. I., Pinney, K., Polito, V. S. Correlation in the tolerance to ozone between sporophytes and male gametophytes of several fruit and nut tree species (Rosaceae). Sexual Plant Reproduction. 9, 44-48 (1996).
- Alcaraz, M. L., Hormaza, J. I., Rodrigo, J. Pistil Starch Reserves at Anthesis Correlate with Final Flower Fate in Avocado (Persea americana). PLOS ONE. 8 (10), 78467 (2013).
- Tao, R., et al. Molecular typing of S-alleles through Identification, Characterization and cDNA cloning for S-RNases in Sweet Cherry. Journal of the American Society for Horticultural Science. 124, 224-233 (1999).
- Burgos, L., et al. The self-compatibility trait of the main apricot cultivars and new selections from breeding programmes. Journal of Horticultural Science. 72, 147-154 (1997).
- Hormaza, J. I., Yamane, H., Rodrigo, J. Apricot. Genome Mapping and Molecular Breeding in Plants, Volume 4 Fruits and Nuts. Kole, C. , Springer. New York, NY. 171-187 (2007).
- Benmoussa, H., Ghrab, M., Ben Mimoun, M., Luedeling, E. Chilling and heat requirements for local and foreign almond (Prunus dulcis Mill.) cultivars in a warm Mediterranean location based on 30 years of phenology records. Agricultural and Forest Meteorology. 239, 34-46 (2017).
- Rodrigo, J., Herrero, M., Hormaza, J. I. Pistil traits and flower fate in apricot (Prunus armeniaca). Annals of Applied Biology. 154, 365-375 (2009).
- Williams, R. R. Techniques used in fruit-set experiments. Towards Regulated Cropping. Williams, R. R., Wilson, D. , Grower Books. London. 57-61 (1970).
- Sutherland, B. G., Robbins, T. P., Tobutt, K. R. Primers amplifying a range of Prunus S-alleles. Plant Breeding. 123, 582-584 (2004).
- Murray, M. G., Thompson, W. F. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Research. 8, 4321-4325 (1980).
- Porebski, S., Bailey, L. G., Baum, B. R. Modification of a CTAB DNA Extraction Protocol for Plants Containing High Polysaccharide and Polyphenol Components. Plant Molecular Biology Reporter. 15 (1), 8-15 (1997).
- Rogers, S. O., Bendich, A. J. Extraction of DNA from milligram amounts of fresh, herbarium and mummified plant tissues. Plant Molecular Biology. 5 (2), 69-76 (1985).
- Hormaza, J. I. Molecular characterization and similarity relationships among apricot (Prunus armeniaca L.) genotypes using simple sequence repeats. Theoretical and Applied Genetics. 104, 321-328 (2002).
- Sonneveld, T., Tobutt, K. R., Robbins, T. P. Allele-specific PCR detection of sweet cherry self-incompatibility (S) alleles S1 to S16 using consensus and allele-specific primers. Theoretical and Applied Genetics. 107, 1059-1070 (2003).
- Hegedus, A., Lénárt, J., Halász, J. Sexual incompatibility in Rosaceae fruit tree species: molecular interactions and evolutionary dynamics. Biologia Plantarum. 56 (2), 201-209 (2012).
- Fernández i Martí, A., Gradziel, T. M., Socias i Company, R. Methylation of the Sf locus in almond is associated with S-RNase loss of function. Plant Molecular Biology. 86, 681-689 (2014).
- Company, R. S. i, Kodad, O., Martí, A. F. i, Alonso, J. M. Mutations conferring self-compatibility in Prunus species: From deletions and insertions to epigenetic alterations. Scientia Horticulturae. 192, 125-131 (2015).
- Boskovic, R., Tobutt, K. R. Correlation of stylar ribonuclease zymograms with incompatibility alleles in sweet cherry. Euphytica. 90, 245-250 (1996).
- Cachi, A. M., Wünsch, A. S-genotyping of sweet cherry varieties from Spain and S-locus diversity in Europe. Euphytica. 197 (2), 229-236 (2014).
- Zuriaga, E., et al. An S-locus Independent Pollen Factor Confers Self-Compatibility in "Katy" Apricot. PLoS ONE. 8 (1), 53947 (2013).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}