

Summary
蛍光顕微鏡による自己適合性とPCR分析によるS-遺伝子型の同定を組み合わせたアプリコット(プルヌスアルメニアカL.)品種の受粉要件を確立する方法論を提示する。
Abstract
酒さにおける自己非適合性は、主に多アレリック軌跡Sによって制御されるGametophytic自己非適合システム(GSI)によって決定される。アプリコットでは、重要な数の新しい品種が放出され、未知の受粉要件を持つ品種が増加しているため、自己および相互互換性関係の決定がますます重要になっています。ここでは、自己(in)の適合性を、手受粉法と顕微鏡による、PCR解析によるS-遺伝子型の同定と組み合わせた方法論を述べた。自己(in)適合性決定のために、各品種からのバルーンステージで花を現場に集め、実験室で手受粉し、固定し、蛍光顕微鏡下で花粉管の挙動を観察するためにアニリンブルーで染色した。品種間の非相溶性関係の確立のために、各品種からDNAを若葉から抽出し、S-アレスをPCRによって同定した。このアプローチは、非互換性グループを確立し、新しい果樹園の設計に適切な受粉者を選択し、繁殖プログラムで適切な親を選択するための貴重な情報を提供する品種間の非互換性関係を解明することができます。
Introduction
自己非適合性は、自己受粉を防ぎ、1を越えることを促進するために植物を開花させる戦略です。酒さでは、この機構は、主に多アレリック軌跡S22によって制御されるGametophytic自己非適合システム(GSI)によって決定される。この様式では、RNase遺伝子はS−sタイラー決定基であるRNase3をコードし3、一方、S−花粉決定基を決定するFボックスタンパク質は、SFB遺伝子4によって体系化される。自己非適合性相互作用は、排卵体5,6,6の受精を防止するスタイルに沿った花粉管の成長の阻害を通じて行われる。
アプリコットでは、品種の更新は、過去20年間で世界中で行われました7,,8.この重要な数の新しい品種の導入は、異なる公的および民間の繁殖プログラムから、未知の受粉要件を有するアプリコット品種の増加をもたらした8。
アプリコットの受粉要件を決定するために、さまざまな方法論が使用されてきました。この分野では、ケージの木や魅惑的な花の中で制御された花粉化によって自己(in)互換性が確立され、その後、果実セット99、10、11、1210,11,12の割合を記録することができる。また、半生体内の花の培養や蛍,光顕微鏡,8、13、14、15、16、1713,14,の下で花粉管挙動の分析によって実験室で制御された受粉が行われている。151617近年、PCR解析やシーケンシングなどの分子技術により、RNase遺伝子とSFB遺伝子18,19,の研究に基づく非相溶関係の特徴付けが可能となった。アプリコットでは、33個のS対立アレス(S1〜S S20、S S22〜S S30、S,52、S S53、S Sv、S Sx)が報告されている(S Sc)12、18、20、21、22、23、24。22,23,2412,18,2021,cこれまで、26の非適合群は、S-遺伝子,型8、9、17、25、26、279,17に従ってこの種で刺されています。,26,27825同じS-allelesSを持つ品種は相互に互換性がありませんが、少なくとも1つの異なるS-対立アレスを持つ品種は、異なる非互換性グループに割り当てられ、相互互換性があります。
アプリコット品種の受粉要件を定義するために、蛍光顕微鏡による自己適合性の決定とアプリコット品種におけるPCR分析によるS-遺伝子型の同定を組み合わせた方法論を記述する。このアプローチは、非互換性グループを確立し、品種間の非互換性関係を解明することができます。
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. 自己(in)互換性判定
- 畑の花を試す。気球段階で花を集める必要があります (図1A),アプリコット28のためのBBCHスケールのステージ58に対応し、不要な以前の受粉を避けるために.
-
実験室での自己受粉とクロス受粉
- バルーンステージで花のアンサーを取り除き、実験室の温度で乾燥させるために紙の上に置きます。
- 24時間後、細かいメッシュ(0.26mm)を用いて花粉粒をふるいにかける(図1B)。
- 自己受粉とクロス受粉のそれぞれについて同じバルーンステージで30本の花のグループをエマキュレートし、実験室の温度で水の中で花屋の泡にピスチルを置く(図1C)。
- 手は、同じ品種24時間後の花から花粉で絵筆の助けを借りて、ピステイルを受粉させる。さらに、各品種の別のピスチルのセットを、コントロールとして互換性のある受粉者の花から花粉で受粉する(図1D)。
- 72時間後、4°C29で少なくとも24時間エタノール/酢酸(3:1)の固定溶液中のピスチルを固定する。その後、固定液を捨て、75%のエタノールを加えて、サンプルが溶液に完全に沈み込まないようにします。サンプル,は、8、17、30、31、32,17,30,31を使用するまで4°Cでこの溶液中で保存することができます。32
-
体外花粉の発芽による花粉の生存率の評価
- 発芽培地を調製するために、スクロースの重量25g、ホウ酸の0.075g(H33BO3)および0.075gの硝酸カルシウム(Ca(NO33)2)33。233
- 250mLの蒸留水に培地の成分を加え、完全に溶解します。
- アガロースの2gを加えた培地を固め、渦巻いて混ぜます。
- pHメーターを使用して培地のpHを確認し、NaOHまたはHCl溶液で値を7.0に調整します。
- 混合物をオートクレーブして培地を殺菌する。
- オートクレーブ処理後、培地を冷やして、滅菌層流れフードでペトリ皿に配ります。
- 固化花粉発芽培地で制御された花粉に使用される同じ品種の花粉粒を散乱し、24時間6後に顕微鏡下でそれらを観察する。
注:ラミナルフローフードを殺菌するには、70%エタノールで表面を清掃し、10分の間にUVランプをオンにします。 - ペトリ皿は使用するまで4°Cの冷蔵庫に保管してください。
-
顕微鏡観察
- 蒸留水で1時間3回ニスティルを洗浄し、4°Cで5%硫酸ナトリウムに入れておきます。24時間後、10分の間に1kg/cm22でオートクレーブし、組織34を軟化させる。
- ガラススライドの上にオートクレーブのピスティルを置き、メスの助けを借りて、花粉管のより良い視覚化を得るために卵巣の周りの毛状突起を取り除きます。次に、カバーガラスでカステイルを押しつぶします。
- 0.1%(v/v)アニリンブルー染色を準備する:0.1 Nリン酸カリウム3塩基の100 mLにアニリンブルーの0.1mLを混ぜる(K3PO4)。花粉管の成長中にカロース堆積物を染色する製剤の上にアニリンブルーの滴を適用します。
- 340-380バンドパスと425のロングパスフィルターを使用して、UV発蛍光を用いた顕微鏡でスタイルに沿って花粉管を観察します。
2. DNA抽出
- 春のフィールドでサンプル2-3の葉。得られたDNAは古い葉と比較してより高品質で低いレベルのフェノール化合物であるため、若い段階で葉をサンプリングすることをお勧めします。
- 市販キットに記載されている手順に従ってゲノムDNAを抽出する(材料表を参照)。
- UV-vis分光光度計(260nm)を用いてDNA濃度の量と質を解析します。
3. S-アレーレ識別
-
PCR 反応の設定
- 各DNA抽出サンプルの蒸留水に50 ng/μLの希釈液を調製します。
- PCR試薬をゆっくりと解凍し、氷の上に保管してください。必要になるまで、DNAポリメラーゼを冷凍庫に入れておきます。
- プライマーの異なる組み合わせを使用して増幅反応を準備します。表 1のコンポーネントを組み合わせて PCR 反応ミックスを作成します。PCR反応をよく混合し、プライマーの異なる組み合わせに対して示された体積をPCRプレートの各ウェルに分配します。次に、各ウェルに1μLのDNA希釈液を加える。
- PCR プレートをサーモサイクラーに入れ、表 1に示す対応する PCR プログラムを実行します。
- 増幅された断片を分析します。PCR増幅断片を分析する方法は主に2種類あります:蛍光標識プライマーを用いたキャピラリー電気泳動(CE)または標識されていないプライマーを有するアガロースゲル電気泳動のアンプリコンを可視化する方法があります。
-
毛細管電気泳動
- ローディングバッファを準備するには、35 μLの脱イオン化ホルムアミドを0.45 μLのラベル付きサイジング規格と混合します。試薬をよく混ぜ合わせ、35.5 μLをリーダープレートのウェルに分配する。
- 1 μLの PCR産物をウェルに加えます。また、水の蒸発を防ぐために、ミネラルオイルの滴を追加します。
- 分離用のプレートを用意し、分離バッファーを追加します。
- 遺伝子分析装置に付属する商用ソフトウェアを使用します (材料表を参照)。新しいサンプルプレートを作成し、プレート上のすべてのウェルのサンプル名を保存します。
- 分析方法を選択します。この場合、サンプルを120sの90°Cで変性し、2.0kVで30sで注入し、6.0kVで35分間分離する。
- 2つのプレートを遺伝子分析装置に挿入します。キャピラリーアレイに蒸留水を充填します。
- 特許取得済みのリニアポリアクリルアミド(LPA)ゲルをロードします。最後に、[ファイルを実行] をクリックします。
-
ゲル電気泳動
- 1x TAE(トリス-アセテート-EDTA)の150 mLに1.5gの分子生物学グレードアガロースを加える1%アガロースゲルを調製します(40 mM Tris、20 mM酢酸、およびpH 8.0で1mM EDTA)。2~3分間マイクロ波加熱でアガロースを溶かします。
- DNAを可視化するには、核酸染色体を4μL加え(材料表を参照)、軽く混ぜます。
- はしご、コントロール、サンプルに十分なウェルを備えたゲルコームをゲルトレイに追加します。その後、ゆっくりとゲルトレイの中央にミックスを注ぎ、泡を避けます。
- ゲルが完全に固まるまで、ゲルを室温で30〜45分間冷却します。電気泳動チャンバにゲルを導入し、ゲルコームを取り外し、ゲルを覆うのに十分な1x TAEバッファーでチャンバーを満たします。
注:ゲルの配置を確認してください。負に荷電したDNAが陰極に向かって移動するので、井戸は負極の近くに置かれるべきです。 - PCR製品に5μLのローディングバッファー(0.1%(v/v)ブロモフェノールブルー)を加え、よく混ぜます。
- バンドのサイズを推定するには、DNA分子量ラダーの5 μLをロードします(材料表を参照)。
- サンプルをゲルの追加ウェルにロードします。
- すべてのサンプルとDNA分子量ラダーをロードしたら、青い染料線がゲルの約75%の長さになるまで、ゲルを1〜1.5時間90Vで実行します。
- 核酸のトランスイルミニューエーターでバンドを視覚化します。
-
毛細管電気泳動
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
アプリコットの受粉研究は、麻酔の前日に風船の後期段階で花を使用する必要があります(図1A)。花の構造はほぼ成熟しているので、この段階は花粉とピスティルコレクションの両方にとって最も有利であると考えられていますが、アンサーの非難はまだ起こっていません。これは、閉じた花びらは外部花粉を運ぶ昆虫の到着を妨げるので、同じ花から花粉だけでなく、他の花から、望ましくない花粉の干渉を防ぎます。花粉粒は、室温またはわずかな余分な熱で24時間紙の上に置かれた脱ヒスズ化したアンサーから、細かいメッシュ(図1B)を通して容易にふるいにかけられている。同様に、ピスティルは、ピンセットまたは爪の助けを借りて花びら、セパルおよびstamensを除去した後、バルーン段階で花から得られる(図1C)。ピスティルは、細かいブラシで自己受粉および交差受粉することができる(図1D)。
アプリコットの雌雄同染色花には、5つの濃い赤いセパル、5つの白い花びら(図1A)、単一のピスティル(図2A)、25-30の雄大な雄花があります。ピスティルは、3つの主要な構造を有する:汚名、スタイルおよび卵巣。卵巣には2つの排卵があり、そのうちの少なくとも1つの受精は果実の設定に必要である。受粉時には、主にミツバチの昆虫が花粉粒を汚名(図1A)に移し、そこで受粉後24時間以内に発芽する(図2B)。花粉管は、各発芽花粉粒から生成され、3〜4日後に卵巣に到達するために、ピスティル構造を介して成長し、約7日後に2つの排卵のうちの1つを受精する。花粉粒のSアレールが2つのピスチルの1つと同じである自己非互換の品種では、花粉管は上層型で成長しなくなり、受精を防ぐ(図2C)。しかし、異なるSアレルを有する適合性品種からの花粉管は、スタイル(図2D)を通して成長し、卵巣(図2E)に達し、2つの卵卵のうちの1つを受精させることができる。
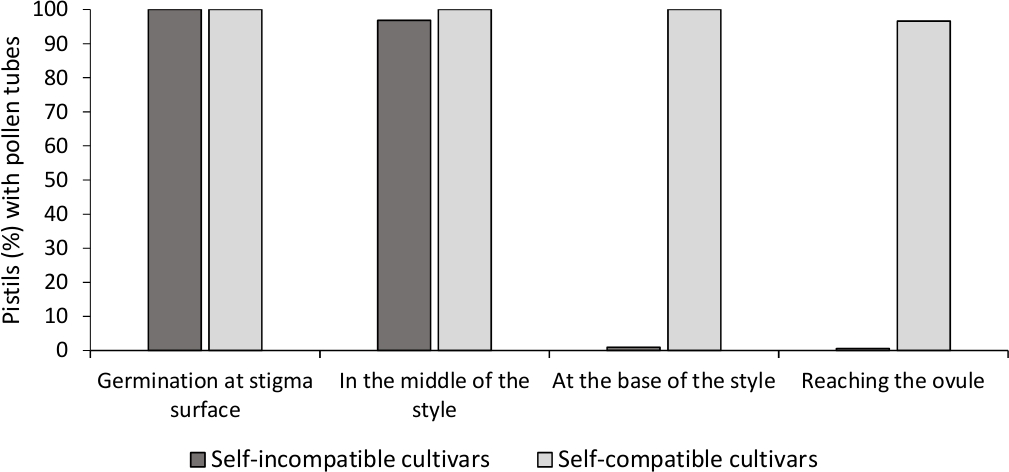
ほとんどの花粉管は、培養培地中で24時間後の花粉粒の長さよりも長かったため、in vitro花粉発芽の分析は、ここで分析されたすべての品種において良好な花粉の生存率を示した。発芽花粉粒は、すべての受粉からピスティルで、スティグマ表面(図2B)で観察され、十分な受粉を示す(図3)。
各品種の自己(in)適合性を決定するために、実験室制御条件で行われた自己及びクロス受粉における花粉管挙動が蛍光顕微鏡下で観察された。花粉管の成長は、検査されたすべてのピスチルでスタイルに沿って記録された。花粉管の成長がほとんどの自己受粉型のピスチル(図2C、図3)のスタイルに沿って逮捕されたとき、品種は自己互換性がないと考えられていた(図2E、図3)。
PCR分析によるS-遺伝子座の検討により、各品種のS-遺伝子型を特徴付けすることができた。まず、S-対立体は、プライマーSRc-F/SRc-Rを用いた最初のS-RNaseイントロンの増幅によって同定された(表2)。増幅断片の大きさを毛細管電気泳動で解析し、対応Aする非適合群で解析した遺伝子型を分類するために用いた(I.G.)(表 3)
S1およびSS7またはSS6SおよびS99のような対立のペアは、第1イントロンに対して同様の断片サイズを示した。したがって、これらの対立両体の分化は、プライマーPru-C2/プルC4R、SHLM1/SHLM2およびSHLM3/SHLM4とRNaseの第2イントロンの領域を増幅することによって行われた(表2)。PruC2/PruC4R プライマーの組み合わせは、S6とS9を区別するために使用されました。S66の場合、1300bpのフラグメントを増幅したのに対し、S99アレルに対して約700bpの断片が観察された(図4B、表3)。特定のプライマーSHLM1/SHLM2およびSHLM3/SHLM4は、S1アレルで約650bp、S7アレルで413bpの断片をS7増幅した(S図4C、表3)。
SFB遺伝子のV2およびHVb可変領域を増幅するプライマーAprFBC8-(F/R)は、両方の対立遺伝子が同一のRNase配列を示すため、SccおよびS88対立遺伝子を区別するために使用された。S88アレーレは約150bpのPCRフラグメントを示し、500bpフラグメントはSccアレーレに対応した(図4D、表3)。S-genotypeが全ての品種について決定されると、自己非互換の品種は、S-対立遺伝子に基づいて対応する非互換性グループに割り当Sてられた(表3)。 S
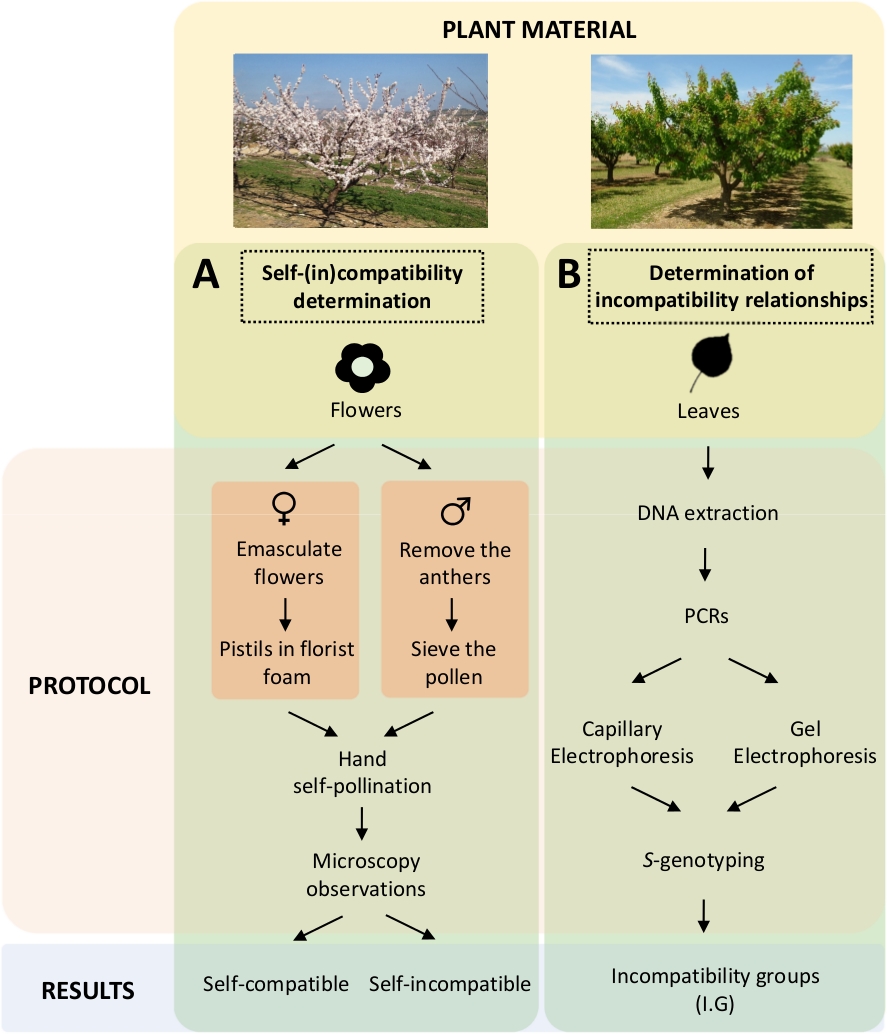
このアプローチでは、遺伝子解析によるS-遺伝子型の特性と併記して、研究室Aにおける制御された自己及びクロス受粉による各品種の自己(in)適合性を決定する必要がある(図5B)。その結果、各品種の受粉要件とアプリコット品種間の非適合性関係を決定することができる。

図 1.アプリコットにおける自己(in)互換性の決定のための実験的なセットアップ。
(A) フィールド内のバルーンステージ(黒矢印)の花。(B)細かいメッシュを使用した花粉粒のふるい。(C)水の花屋の泡の上に置かれるピスティル。(D) 絵筆を使って、毛の手受粉を行う。この図の大きなバージョンを表示するには、ここをクリックしてください。

図 2.アプリコット花におけるゲームの不適合関係の図示的表現。
(A) Gametophytic自己非適合性(GSI)では、互換性のある花粉粒と互換性のない花粉粒の両方がスティグマに発芽する。花粉粒は、元の遺伝子2型の2つのS-対立遺伝子のうちのS1つを運びます。 S花粉粒のS-対立体が、2つのS-対立胞の1つに一致する場合、この場合S11S33は、花粉管の成長がスタイルの上の1/3で阻害される。(B) 汚名表面上の花粉粒の発芽。(C) 花粉管は、相性の無い行動を示すスタイルで逮捕された。(D) 花粉管は、スタイルに沿って成長しています。(E) スタイルの基部にある花粉管は、互換性のある動作を示します。スケールバー、100 μm。この図の大きなバージョンを表示するには、ここをクリックしてください。

図 3.自己受粉後の自己互換性と自己互換性のない品種のスタイルを通じて花粉発芽と花粉管成長の代表的な結果.
花粉粒が汚名表面で発芽し、花粉管がスタイルの途中、スタイルの基部にあり、排卵に達する花粉粒の割合。この図の大きなバージョンを表示するには、ここをクリックしてください。

図 4.S-アレルの同定のための5つのプライマー対の組み合わせを用いたPCR断片増幅。
(A)SRc-(F/R)プライマーの遺伝子分析出力は、RNaseの第1イントロン領域の2つの増幅断片のサイズを示すS-アレルに対応する。SB(B)S6および6S9対立素の同定のためのプライマーPruC2/PruC4RをS用いたPCR増幅。(C)S1アレスおよびSHLM3およびSHLM4の分化のために特異的なプライマーSHLM1およびSHLM2を用いて得られたPCR産物はS1、S77アレーレを区別する。C(D) ScおよびS8対立を識別するためのAprFBC8-(F/R)プライマーを用いたPCR増幅。MI: 1 kb DNA ラダー。MII:100 bp DNAラダーこの図の大きなバージョンを表示するには、ここをクリックしてください。

図 5.アプリコット品種における自己適合性と相互互換性関係を解明するための実験計画のスキーム。
(A)研究室における制御された受粉による自己適合性判定のワークフロー(B) 分子アプローチによるS-アリール同定のワークフローこの図の大きなバージョンを表示するには、ここをクリックしてください。
| PCR マスター ミックス | サーモサイクラーの状態 | ||||||
| コンポーネント | 最終濃度 | 15 μL反応 | サイクルステップ | 温度 | 時間 | サイクル | |
| 10x NH4反応バッファー | 10倍 | 1.5 μL | 初期変性 | 94°C | 3分 | 1 | |
| 50 mM MgCl2溶液 | 25mM | 1.2 μL | 変化 | 94°C | 1分 | 35 | |
| 100 mM dNTP | 2.5 mM | 0.6 μL | 焼鈍 | 55°C | 1分 | ||
| プライマー SRc-F | 10 μM | 0.6 μL | 拡張子 | 72°C | 3分 | ||
| プライマー SRc-R | 10 μM | 0.6 μL | 最終拡張 | 72°C | 5分 | 1 | |
| 500 U Taq DNA ポリメラーゼ | 0.5 U | 0.2 μL | 4°C | 保持 | |||
| H2O | 8.3 μL | ||||||
| コンポーネント | 最終濃度 | 25 μL反応 | サイクルステップ | 温度 | 時間 | サイクル | |
| 10x PCR バッファー | 10倍 | 2.5 μL | 初期変性 | 94°C | 2分 | 1 | |
| 5倍のQソリューション | 5x | 5 μL | 変化 | 94°C | 10 s | 10 | |
| 100 mM dNTP | 2.5 mM | 0.5 μL | 焼鈍 | 55°C | 2分 | ||
| プライマープル2 | 10 μM | 0.2 μL | 拡張子 | 68°C | 2分 | ||
| プライマー C4R | 10 μM | 0.2 μL | 変化 | 94°C | 10 s | 25 | |
| 250 U Taq DNAポリメラーゼ | 10 U | 0.13 μL | 焼鈍 | 58°C | 2分 | ||
| H2O | 15.5 μL | エクステンション* | 68°C | 2分 | |||
| 最終拡張 | 72°C | 5分 | 1 | ||||
| 4°C | 保持 | ||||||
| * 10 s で、各サイクルを 68 %C 拡張ステップに追加しました。 | |||||||
| コンポーネント | 最終濃度 | 25 μL反応 | サイクルステップ | 温度 | 時間 | サイクル | |
| 10x PCR バッファー | 10倍 | 2.5 μL | 初期変性 | 94°C | 2分 | 1 | |
| 5倍のQソリューション | 5x | 5 μL | 変化 | 94°C | 30 s | 35 | |
| 100 mM dNTP | 2.5 mM | 0.5 μL | 焼鈍 | 62°C | 1.5分 | ||
| プライマー SHLM1 | 10 μM | 0.2 μL | 拡張子 | 72°C | 2分 | ||
| プライマー SHLM2 | 10 μM | 0.2 μL | 最終拡張 | 72°C | 5分 | 1 | |
| 250 U Taq DNAポリメラーゼ | 10 U | 0.13 μL | 4°C | 保持 | |||
| H2O | 15.5 μL | ||||||
| コンポーネント | 最終濃度 | 20 μL反応 | サイクルステップ | 温度 | 時間 | サイクル | |
| 5x PCR バッファー | 5x | 4 μL | 初期変性 | 98°C | 30 s | 1 | |
| dNTP | 2.5 mM | 1.6 μL | 変化 | 98°C | 10 s | 35 | |
| プライマー SHLM3 | 10 μM | 1 μL | 焼鈍 | 51°C | 30 s | ||
| プライマー SHLM4 | 10 μM | 1 μL | 拡張子 | 72°C | 1分 | ||
| 100 U DNA ポリメラーゼ | 5 U | 0.2 μL | 最終拡張 | 72°C | 5分 | 1 | |
| H2O | 12.4 μL | 4°C | 保持 | ||||
| コンポーネント | 最終濃度 | 25 μL反応 | サイクルステップ | 温度 | 時間 | サイクル | |
| 10x PCR バッファー | 10倍 | 2.5 μL | 初期変性 | 94°C | 2分 | 1 | |
| 100 mM dNTP | 2.5 mM | 2 μL | 変化 | 94°C | 30 s | 35 | |
| プライマー FBC8-F | 10 μM | 1 μL | 焼鈍 | 55°C | 1.5分 | ||
| プライマー FBC8-R | 10 μM | 1 μL | 拡張子 | 72°C | 2分 | ||
| 250 U Taq DNAポリメラーゼ | 10 U | 0.125 μL | 最終拡張 | 72°C | 5分 | 1 | |
| H2O | 17.4 μL | 4°C | 保持 | ||||
表 1.このプロトコルで使用される異なるプライマーの組み合わせのための反応およびサイクリング条件。
| プライマー | シーケンス | 参照 |
| SRc-F | 5'-CTCGCTTCCTTTGTTCTTGC-3' | 18 |
| SRc-R | 5'-GGCCATTGTTTTTTG-3' | 18 |
| プル-C2 | 5'-CTTTGGCCAGTTAtTCAAACC-3' | 35 |
| プル-C4R | 5'-GGATGTGGTACGGTGAAGCG-3' | 35 |
| SHLM1-F | 5'-GGTGGAGGTガタタグタグック-3' | 17 |
| シュマム2-R | 5'-GGCTGCAAGGAAGCTGG-3' | 17 |
| SHLM3-F | 5'-タタットクトクトクトクトクトッグ3' | 17 |
| シュマム4-R | 5'-カクタットガタートGTGTATG-3' | 17 |
| 4月8日-F | 5'-CATGGAAAAAAAGCTACTTATGG-3' | 26 |
| 4月8日-R | 5'-GCCTAATGTCATCTCTCTTAG-3' | 26 |
表 2.このプロトコルで使用されるプライマーは、PrunusアルメニアカにおけるS-遺伝子型特性の配列および参照です。
| 品種 | SRc-(F/R) (bp) | プルC2/プルC4R (bp) | SHLM1/SHLM2 (bp) | SHLM3/SHLM4 (bp) | AprFBC8-(F/R) (bp) | S-遺伝子型 | 非互換性グループ (I.G) |
| ワンダーコット8 | 420, 420 | 749, 1386 | S6S9 | Viii | |||
| マジックコット8 | 334, 420 | 749 | S2S9 | Xx | |||
| ゴールドストライク8 | 334, 420 | 749 | S2S9 | - | |||
| T06917 | 334, 408 | 650 | S1S2 | 私 | |||
| T12017 | 334, 408 | 650 | S1S2 | - | |||
| C-6 | 334, 408 | 413 | S2S7 | Iv | |||
| クーパーコット8 | 274, 408 | 650 | S1S3 | 188年 | |||
| アプリクイーン | 358, 358 | 500 | ScSc | - | |||
| ベルゲコット8 | 334, 358 | 500 | S2Sc | - | |||
| スプリングブラッシュ8 | 274, 358 | 150 | S3S8 | XXI |
表 3.このプロトコルと非適合性グループの割り当てで使用される5つのプライマーペアを持つアプリコット品種のS-ジェノタイピング。SRc-(F/R)、プルC2/プルC4R、SHLM1/SHLM2、SHLM3/SHLM4、およびAprFBC8-(F/R)プライマーを使用して増幅されたS-対立胞の異なるポリメラーゼ連鎖反応生成物サイズを表に示す。
Subscription Required. Please recommend JoVE to your librarian.
Discussion
伝統的に、ほとんどの市販アプリコットヨーロッパの品種は、自己互換性のある36でした。それにもかかわらず、過去数十年の繁殖プログラムにおける親としての北米の自己適合性品種の使用は、未知の受粉要件77、8、378,37を持つ新しい自己互換性のない品種の増加をもたらした。したがって、アプリコット品種における自己適合性と相互互換性関係の決定はますます重要になっています。これは、冬の冷えが減少している地域で強調され、開花時の高年から年の変動は、多くの場合、特に高い冷たい要件を持つ品種で、品種の開花の偶然を防いでいるので38。本明細書に記載されている方法論は、手受粉、顕微鏡および遺伝子解析を組み合わせることによって、各品種の自己(in)相溶性を決定し、その潜在的な受粉体品種を確立するのに非常に有用であった。
受粉要件は、果樹園条件11,39,39におけるフィールド制御実験を通じて決定することができる。しかし、気象不利な条件を含む外的要因への博覧会は、受粉障害10を引き起こし、自己非適合性の誤った診断を引き起こす可能性がある。本明細書に記載された方法論は、実験室で制御された条件での手受粉花の顕微鏡観察によって自己(in)相溶性をより正確に評価し、環境への影響を回避することを可能にする。さらに、このアプローチは、フィールド実験40で必要とされる品種ごとに数本の成虫の木の代わりに少数の花が必要であるので、年間に多くの品種を分析することを可能にする。
非互換性関係は、手受粉と顕微鏡14を組み合わせて確立することができる。しかし、花の寿命が非常に短いため、花の開花期には短時間しか行う必要がないため、花粉は非常に短い。したがって、各季節における制御された手受粉によって分析できる非互換性関係の数は非常に少ない。S-locusによってコードされる遺伝子の特性S評価により、S-アレーレ遺伝子型化18,41のPCRベースの方法の18開発が可能になった。このアプローチは花を必要としないのでS-アリール同定を加速し、実験は任意の栄養組織42で行うことができる。これは、植物材料、通常若い葉、43を収集することができる期間を延長します。さらに、葉は凍結乾燥または凍結することができるので、開花期44の間に新鮮な花にしか行えることができない花粉化とは異なり、分析は一年中いつでも行うことができる。追加の利点は、開花期に入る前に若い木から葉を収集することができるということです, サンプルの収集を容易にし、結果45の早期取得.
遺伝子解析は、増幅されたフラグメントサイズ21,46,46の精密な結果を提供するので自己非適合性対立遺伝子のより良い分化を可能にする。現在までに、33個の,S-対立,遺伝子が,22,、S-遺伝子型,,,,8、9、17、25、26、27に基づく2636の非互換性グループを確立することを可能にしたアプリコット25817912、18、20、21、22、23、24,27で同定されている。121820212324,一方、この方法論の欠点は、同じ範囲の大きさまたは突然変異の異なる対立遺伝子が同じ対立遺伝子として誤って同定され得る点である。したがって、SccおよびS88対立遺伝子はRNase配列について同一であるが、Sc1919のSFB遺伝子に358bp挿入が見られる。同様に、対立胞S11およびS77の第1イントロン領域は同一であり、プライマーSRc-F/SRc-Rを用いて区別がつかない。また、S6、S6 S528、S20、S55、S7、S8 S20 S7 S5513(EF062341)、S S46 S17などのいくつかの相同性は、これらの対立遺伝子の一部が部分的に配列されているか、PCR増幅中の障害によって検出され、その結果、それらを正しく区別するためにさらなる作業が必要です。17
PCR分析およびS-RNaseシーケンシングは、S-対立体の同定および対応する非適合性グループ,8、17、26、2717における品種の割り当てを通8じて非互換性関係を確立するのに十分である。26,27しかし、この方法論には、特定のアプリコット品種に対する自己(in)互換性の決定を阻止するという制限がある。自己適合性(SC)は、アーモンド(Sff)48、49または49スウィートチェリー(S44')50、51として、他のプルナス種47の特定50のS-対立アレスに51関連付けられている。48しかし、アプリコットでは、SC21に関連付けられているScアレスは、誤ってS8、自己非互換アレスレ19、22、およびS遺伝子座19,22にリンクされていない変異として誤って同定され得る、M-locus12、52、SCを授与するように、SCを与える。12,52最近、M-locusはSSRマーカー12を用いて遺伝子型化されている。したがって、アプリコット遺伝子型のSCの遺伝子同定にはさらなる研究が必要であり、S遺伝子座に関連しない要因によるミスを避けるために、この研究では自己受粉花のピスチを通した花粉管の挙動をフェノタイピングすることによって自己(in)の相性の特徴付けも決定されている。
本明細書に記載されている方法論は、実験室条件における手受けによる自己受光性の決定と、その後の蛍光顕微鏡下での制御された自己受粉の病態における花粉管の挙動の観察と、PCR分析によるS-genotypeの同定を組Sみ合わせることにより、アプリコット品種の受粉要件を確立することを可能にする。これは、新しい果樹園の設計に適切な受粉者を選択するだけでなく、アプリコットの繁殖プログラムで新しい十字架を設計するために適切な親を選択するために品種間の非互換性関係を確立することができるので、生産者やブリーダーのための貴重な情報を提供します。
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
著者らは開示するものは何もない。
Acknowledgments
この研究は、シエンシア大臣、イノベルシダーデス・イ・ウニバーシダーデス・ヨーロピアン地域開発基金、欧州連合(AGL2016-77267-R、AGL2015-74071-JIN)によって資金提供されました。インスティトゥート・ナシオナル・デ・インベスティガシオン・イ・テクノロジア・アグラリア・イ・アリメンタリア(RFP2015-00015-00,RTA2017-00003-00);ゴビエルノ・デ・アラゴン・ヨーロピアン・ソーシャル・ファンド、欧州連合(グルポ・コンソリダド・A12_17R)、フンダシオン・バイオディビジダード、アグロゼグロS.A.
Materials
| Name | Company | Catalog Number | Comments |
| Agarose D1 Low EEO | Conda | 8010.22 | |
| BIOTAQ DNA Polymerase kit | Bioline | BIO-21060 | |
| Bright field microscope | Leica Microsystems | DM2500 | |
| CEQ System Software | Beckman Coulter | ||
| DNeasy Plant Mini Kit | QIAGEN | 69106 | |
| dNTP Set, 4 x 25 µmol | Bioline | BIO-39025 | |
| GenomeLab DNA Size Standard Kit - 400 | Beckman Coulter | 608098 | |
| GenomeLab GeXP Genetic Analysis System | Beckman Coulter | ||
| GenomeLab Separation Buffer | Beckman Coulter | 608012 | |
| GenomeLab Separation Gel LPA-1 | Beckman Coulter | 391438 | |
| HyperLadder 100bp | Bioline | BIO-33029 | |
| HyperLadder 1kb | Bioline | BIO-33025 | |
| Image Analysis System | Leica Microsystems | ||
| Molecular Imager VersaDoc MP 4000 system | Bio-Rad | 170-8640 | |
| NanoDrop One Spectrophotometer | Thermo Fisher Scientific | 13-400-518 | |
| pH-Meter BASIC 20 | Crison | ||
| Phusion High-Fidelity PCR Kit | Thermo Fisher Scientific | F553S | |
| Power Pack P 25 T | Biometra | ||
| Primer Forward | Isogen Life Science | ||
| Primer Reverse | Isogen Life Science | ||
| Quantity One Software | Bio-Rad | ||
| Stereoscopic microscope | Leica Microsystems | MZ-16 | |
| Sub-Cell GT | Bio-Rad | ||
| SYBR Safe DNA Gel Stain | Thermo Fisher Scientific | S33102 | |
| T100 Thermal Cycler | Bio-Rad | 1861096 | |
| Taq DNA Polymerase | QIAGEN | 201203 | |
| Vertical Stand Autoclave | JP Selecta |
References
- Silva, N. F., Goring, D. R. Mechanisms of self-incompatibility in flowering plants. Cellular and Molecular Life Sciences. 58, 1988-2007 (2001).
- Charlesworth, D., Vekemans, X., Castric, V., Glémin, S. Plant self-incompatibility systems: A molecular evolutionary perspective. New phytologist. 168, 61-69 (2005).
- Tao, R., et al. Identification of stylar RNases associated with gametophytic self-incompatibility in almond (Prunus dulcis). Plant and Cell Physiology. 38, 304-311 (1997).
- Ushijima, K., et al. Structural and transcriptional analysis of the self-incompatibility locus of almond: Identification of a pollen-expressed F-box gene with haplotype-specific polymorphism. The Plant cell. 15, 771-781 (2003).
- Bedinger, P. A., Broz, A. K., Tovar-Mendez, A., McClure, B. Pollen-Pistil Interactions and Their Role in Mate Selection. Plant Physiology. 173, 79-90 (2017).
- Guerra, M. E., Rodrigo, J. Japanese plum pollination: A review. Scientia Horticulturae. 197, 674-686 (2015).
- Zhebentyayeva, T., Ledbetter, C., Burgos, L., Llacer, G. Apricot. Fruit Breeding. Badenes, M. L., Byrne, D. , Springer. Boston, MA. 415-458 (2012).
- Herrera, S., Lora, J., Hormaza, J. I., Herrero, M., Rodrigo, J. Optimizing Production in the New Generation of Apricot Cultivars: Self-incompatibility, S-RNase Allele Identification, and Incompatibility Group Assignment. Frontiers in Plant Science. 9, 527 (2018).
- Egea, J., Burgos, L. Detecting Cross-incompatibility of Three North American Apricot Cultivars and Establishing the First Incompatibility Group in Apricot. Journal of the American Society for Horticultural Science. 121, 1002-1005 (1996).
- Rodrigo, J., Herrero, M. Effects of pre-blossom temperatures on flower development and fruit set in apricot. Scientia Horticulturae. 92, 125-135 (2002).
- Julian, C., Herrero, M., Rodrigo, J. Flower bud differentiation and development in fruiting and non-fruiting shoots in relation to fruit set in apricot (Prunus armeniaca). Trees. 24, 833-841 (2010).
- Muñoz-Sanz, J. V., Zuriaga, E., López, I., Badenes, M. L., Romero, C. Self-(in)compatibility in apricot germplasm is controlled by two major loci, S and M. BMC Plant Biology. 17, 82 (2017).
- Burgos, L., Berenguer, T., Egea, J. Self- and Cross-compatibility among Apricot Cultivars. HortScience. 28, 148-150 (1993).
- Rodrigo, J., Herrero, M. Evaluation of pollination as the cause of erratic fruit set in apricot "Moniqui". Journal of Horticultural Science. 71, 801-805 (1996).
- Milatović, D., Nikolić, D., Krška, B. Testing of self-(in)compatibility in apricot cultivars from European breeding programmes. Horticultural Science. 40 (2), 65-71 (2013).
- Milatović, D., Nikolić, D., Fotirić-Aksić, M., Radović, A. Testing of self-(in)compatibility in apricot cultivars using fluorescence microscopy. Acta Scientiarum Polonorum, Hortorum Cultus. 12 (6), 103-113 (2013).
- Herrera, S., Rodrigo, J., Hormaza, J. I., Lora, J. Identification of Self-Incompatibility Alleles by Specific PCR Analysis and S-RNase Sequencing in Apricot. Int J Mol Sci. 19, 3612 (2018).
- Romero, C., et al. Analysis of the S-locus structure in Prunus armeniaca L. Identification of S-haplotype specific S-RNase and F-box genes. Plant Molecular Biology. 56, 145-157 (2004).
- Halász, J., Pedryc, A., Hegedus, A. Origin and dissemination of the pollen-part mutated SC haplotype which confers self-compatibility in apricot (Prunus armeniaca). New Phytologist. 176, 792-803 (2007).
- Halász, J., Hegedus, A., Hermán, R., Stefanovits-Bányai, É, Pedryc, A. New self-incompatibility alleles in apricot (Prunus armeniaca L.) revealed by stylar ribonuclease assay and S-PCR analysis. Euphytica. 145, 57-66 (2005).
- Vilanova, S., Romero, C., Llacer, G., Badenes, M. L., Burgos, L. Identification of Self-(in)compatibility Alleles in Apricot by PCR and Sequence Analysis. Journal of the American Society for Horticultural Science. 130, 893-898 (2005).
- Feng, J., et al. Detection and transcript expression of S-RNase gene associated with self-incompatibility in apricot (Prunus armeniaca L.). Molecular Biology Reports. 33, 215-221 (2006).
- Zhang, L., et al. Identification of self-incompatibility (S-) genotypes of Chinese apricot cultivars. Euphytica. 160, 241-248 (2008).
- Wu, J., et al. Identification of S-haplotype-specific S-RNase and SFB alleles in native Chinese apricot (Prunus armeniaca L). Journal of Horticultural Science and Biotechnology. 84, 645-652 (2009).
- Szabó, Z., Nyéki, J. Blossoming, fructification and combination of apricot varieties. Acta Horticulturae. 293, 295-302 (1991).
- Halász, J., Pedryc, A., Ercisli, S., Yilmaz, K. U., Hegedűs, A. S-genotyping supports the genetic relationships between Turkish and Hungarian apricot germplasm. Journal of the American Society for Horticultural Science. 135, 410-417 (2010).
- Lachkar, A., et al. Identification of self-(in)compatibility S-alleles and new cross-incompatibility groups in Tunisian apricot (Prunus armeniaca L.) cultivars. The Journal of Horticultural Science and Biotechnology. 88, 497-501 (2013).
- Pérez-Pastor, A., Ruiz-Sánchez, M. C., Domingo, R., Torrecillas, A. Growth and phenological stages of Búlida apricot trees in South-East. Agronomie. 24, 93-100 (2004).
- Williams, J. H., Friedman, W. E., Arnold, M. L. Developmental selection within the angiosperm style: using gamete DNA to visualize interspecific pollen competition. Proceedings of the National Academy of Sciences of the United States of America. 96, 9201-9206 (1999).
- Julian, C., Herrero, M., Rodrigo, J. Anther meiosis time is related to winter cold temperatures in apricot (Prunus armeniaca L.). Environmental and Experimental Botany. 100, 20-25 (2014).
- Guerra, M. E., López-Corrales, M., Wünsch, A., Rodrigo, J. Lack of Fruit Set Caused by Ovule Degeneration in Japanese Plum. Journal of the American Society for Horticultural Science. 136 (6), 375-381 (2011).
- Guerra, M. E., Wünsch, A., López-Corrales, M., Rodrigo, J. Flower Emasculation as the Cause for Lack of Fruit Set in Japanese Plum Crosses. Journal of the American Society for Horticultural Science. 135 (6), 556-562 (2010).
- Hormaza, J. I., Pinney, K., Polito, V. S. Correlation in the tolerance to ozone between sporophytes and male gametophytes of several fruit and nut tree species (Rosaceae). Sexual Plant Reproduction. 9, 44-48 (1996).
- Alcaraz, M. L., Hormaza, J. I., Rodrigo, J. Pistil Starch Reserves at Anthesis Correlate with Final Flower Fate in Avocado (Persea americana). PLOS ONE. 8 (10), 78467 (2013).
- Tao, R., et al. Molecular typing of S-alleles through Identification, Characterization and cDNA cloning for S-RNases in Sweet Cherry. Journal of the American Society for Horticultural Science. 124, 224-233 (1999).
- Burgos, L., et al. The self-compatibility trait of the main apricot cultivars and new selections from breeding programmes. Journal of Horticultural Science. 72, 147-154 (1997).
- Hormaza, J. I., Yamane, H., Rodrigo, J. Apricot. Genome Mapping and Molecular Breeding in Plants, Volume 4 Fruits and Nuts. Kole, C. , Springer. New York, NY. 171-187 (2007).
- Benmoussa, H., Ghrab, M., Ben Mimoun, M., Luedeling, E. Chilling and heat requirements for local and foreign almond (Prunus dulcis Mill.) cultivars in a warm Mediterranean location based on 30 years of phenology records. Agricultural and Forest Meteorology. 239, 34-46 (2017).
- Rodrigo, J., Herrero, M., Hormaza, J. I. Pistil traits and flower fate in apricot (Prunus armeniaca). Annals of Applied Biology. 154, 365-375 (2009).
- Williams, R. R. Techniques used in fruit-set experiments. Towards Regulated Cropping. Williams, R. R., Wilson, D. , Grower Books. London. 57-61 (1970).
- Sutherland, B. G., Robbins, T. P., Tobutt, K. R. Primers amplifying a range of Prunus S-alleles. Plant Breeding. 123, 582-584 (2004).
- Murray, M. G., Thompson, W. F. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Research. 8, 4321-4325 (1980).
- Porebski, S., Bailey, L. G., Baum, B. R. Modification of a CTAB DNA Extraction Protocol for Plants Containing High Polysaccharide and Polyphenol Components. Plant Molecular Biology Reporter. 15 (1), 8-15 (1997).
- Rogers, S. O., Bendich, A. J. Extraction of DNA from milligram amounts of fresh, herbarium and mummified plant tissues. Plant Molecular Biology. 5 (2), 69-76 (1985).
- Hormaza, J. I. Molecular characterization and similarity relationships among apricot (Prunus armeniaca L.) genotypes using simple sequence repeats. Theoretical and Applied Genetics. 104, 321-328 (2002).
- Sonneveld, T., Tobutt, K. R., Robbins, T. P. Allele-specific PCR detection of sweet cherry self-incompatibility (S) alleles S1 to S16 using consensus and allele-specific primers. Theoretical and Applied Genetics. 107, 1059-1070 (2003).
- Hegedus, A., Lénárt, J., Halász, J. Sexual incompatibility in Rosaceae fruit tree species: molecular interactions and evolutionary dynamics. Biologia Plantarum. 56 (2), 201-209 (2012).
- Fernández i Martí, A., Gradziel, T. M., Socias i Company, R. Methylation of the Sf locus in almond is associated with S-RNase loss of function. Plant Molecular Biology. 86, 681-689 (2014).
- Company, R. S. i, Kodad, O., Martí, A. F. i, Alonso, J. M. Mutations conferring self-compatibility in Prunus species: From deletions and insertions to epigenetic alterations. Scientia Horticulturae. 192, 125-131 (2015).
- Boskovic, R., Tobutt, K. R. Correlation of stylar ribonuclease zymograms with incompatibility alleles in sweet cherry. Euphytica. 90, 245-250 (1996).
- Cachi, A. M., Wünsch, A. S-genotyping of sweet cherry varieties from Spain and S-locus diversity in Europe. Euphytica. 197 (2), 229-236 (2014).
- Zuriaga, E., et al. An S-locus Independent Pollen Factor Confers Self-Compatibility in "Katy" Apricot. PLoS ONE. 8 (1), 53947 (2013).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}