

Summary
Vi presenterer en metodikk for å etablere pollineringskravene til aprikos (Prunus armeniaca L.) sorter som kombinerer fastsettelse av selv-(i)kompatibilitet ved fluorescensmikroskopi med identifisering av S-genotype ved PCR-analyse.
Abstract
Selvinkompatibilitet i Rosaceae bestemmes av et Gametophytic Self-Incompatibility System (GSI) som hovedsakelig styres av multiallelic locus S. I aprikos er fastsettelsen av selv- og inter-(in)kompatibilitetsforhold stadig viktigere, siden utgivelsen av et viktig antall nye sorter har resultert i økningen av sorter med ukjente pollineringskrav. Her beskriver vi en metodikk som kombinerer fastsettelse av selv-(i)kompatibilitet ved hånd-pollineringer og mikroskopi med identifisering av S-genotype etter PCR-analyse. For selvbestemmelse av kompatibilitet ble blomster på ballongstadiet fra hver sortiator samlet i feltet, håndpollinert i laboratoriet, fast og farget med anilinblå for observasjon av pollenrøratferd under fluorescensmikroskopien. For etablering av inkompatibilitetsforhold mellom sorter ble DNA fra hver sort ble hentet fra unge blader og S-alleles identifisert av PCR. Denne tilnærmingen gjør det mulig å etablere inkompatibilitetsgrupper og belyse inkompatibilitetsforhold mellom sorter, som gir en verdifull informasjon for å velge egnede pollinizers i utformingen av nye frukthager og å velge passende foreldre i avlsprogrammer.
Introduction
Selvinkompatibilitet er en strategi for blomstrende planter for å hindre selvpollinering og fremme outcrossing1. I Rosaceae, denne mekanismen bestemmes av en Gametophytic Self-Incompatibility System (GSI) som hovedsakelig styres av multiallelic locus S2. I stil, RNase genet koder S-stylar determinant, en RNase3, mens en F-boks protein, som bestemmer S-pollen determinant, er kodifisert av SFB genet4. Den selvinkompatibilitet interaksjon finner sted gjennom hemming av pollen rør vekst langs stilen hindrer befruktning av ovule5,6.
I aprikos har en variantfornyelse funnet sted over hele verden i de siste to tiårene7,8. Denne innføringen av et viktig antall nye sorter, fra ulike offentlige og private avlsprogrammer, har resultert i økningen av aprikoskulter med ukjente pollineringskrav8.
Ulike metoder har blitt brukt til å bestemme pollineringskrav i aprikos. I feltet kan selvkompatibilitet etableres ved kontrollerte pollineringer i burtrær eller i emasculated blomster og deretter registrere prosentandelen av frukt sett9,10,11,12. I tillegg har kontrollerte pollineringer blitt utført i laboratoriet av semi-in vivo kultur av blomster og analyse av pollen rør atferd under fluorescens mikroskopi8,13,14,15,16,17. Nylig har molekylære teknikker, som PCR-analyse og sekvensering, tillatt karakterisering av inkompatibilitetsrelasjoner basert på studiet av RNase- og SFB-genene 18,19. I aprikos, trettitre S-alleles har blitt rapportert (S1 til S20, S22 til S30, S52, S53, Sv, Sx), inkludert en allele relatert til selvkompatibilitet (Sc)12,18,20,21,22,23,24. Hittil har 26 inkompatibilitetsgrupper blitt knivstukket i denne arten i henhold til S-genotype8,9,17,25,26,27. Sorter med samme S-alleles er inter-uforenlige, mens sorter med minst en annen S-allele og dermed tildelt i forskjellige inkompatible grupper, er inter-kompatible.
For å definere pollineringskravene til aprikoskuler, beskriver vi en metodikk som kombinerer fastsettelse av selv-(i)kompatibilitet ved fluorescensmikroskopi med identifisering av S- genotype etter PCR-analysei aprikoskuler. Denne tilnærmingen gjør det mulig å etablere inkompatibilitetsgrupper og belyse inkompatibilitetsrelasjoner mellom sorter.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. Bestemmelse av selvkompatibilitet
- Prøv blomstene i feltet. Det er nødvendig å samle blomstene på ballongstadiet (Figur 1A), tilsvarende trinn 58 på BBCH-skalaen for aprikos28, for å unngå uønsket tidligere pollinering.
-
Selv- og krysspollineringer i laboratoriet
- Fjern anthers av blomstene på ballong scenen og plassere dem på et stykke papir for å tørke ved laboratorietemperatur.
- Etter 24 timer kan du sile pollenkornene ved hjelp av et fint nett (0,26 mm) (Figur 1B).
- Emasculate en gruppe på 30 blomster på samme ballong scenen for hver selv-pollinering og kryss-pollinering og plassere pistils på blomsterhandler skum i vann ved laboratorietemperatur (Figur 1C).
- Hånd polliner pistils ved hjelp av en pensel med pollen fra blomster av samme sorta 24 t etter emasculation. I tillegg pollinerer et annet sett med støvkjeringer av hver sort med pollen fra blomster av en kompatibel pollinizer som kontroll (Figur 1D).
- Etter 72 timer, fest støvlene i en fikseringsløsning av etanol/eddiksyre (3:1) i minst 24 timer ved 4 °C29. Kast deretter fikseringen og tilsett 75% etanol som sikrer at prøvene er helt nedsenket i løsningen. Prøver kan bevares i denne oppløsningen ved 4 °C til bruk8,,17,,30,,31,,32.
-
Evaluering av pollen levedyktighet gjennom in vitro pollen spiring
- For å forberede spiringsmediet, vekt 25 g sukrose, 0,075 g borsyre (H3BO3) og 0,075 g kalsiumnitrat (Ca(NO3)2)33.
- Tilsett komponentene i mediet i 250 ml destillert vann og oppløses helt.
- Stivner mediet og tilsett 2 g agarose og bland ved å virvle.
- Kontroller pH-verdien på mediet ved hjelp av en pH-måler, og juster verdien til 7,0 med NaOH- eller HCl-oppløsning.
- Autoklav blandingen for å sterilisere mediet.
- Etter autoklavering, avkjøl mediet og fordel det i Petri retter i en steril laminær flythette.
- Spre pollenkornene til de samme sortiene som brukes til de kontrollerte pollineringene i det størknede pollenspiringsmediet og observer dem under mikroskopet etter 24 t6.
MERK: For å sterilisere laminærstrømningshetten, rengjør du overflaten med 70 % etanol og slår på UV-lampen i løpet av 10 minutter. - Oppbevar Petri-rettene i kjøleskap ved 4 °C til bruk.
-
Mikroskopi observasjoner
- Vask støvlene tre ganger i 1 timer med destillert vann og la dem være i 5 % natriumsulfitt ved 4 °C. Etter 24 timer, autoklaver dem på 1 kg/ cm2 under 10 min i natriumsulfitt for å myke vevet34.
- Plasser de autoklavete pistils over et glasssklie, og ved hjelp av en skalpell, fjern trichomes rundt eggstokken for å få en bedre visualisering av pollenrørene. Deretter squash pistils med et dekk glass.
- Forbered 0,1 % (v/v) anilinblå beis: bland 0,1 ml anilinblå i 100 ml 0,1 N kaliumfosfattribasisk (K3PO4). Påfør en dråpe anilin blå over preparatene for å beis callose avsetninger under pollen rør vekst.
- Vær oppmerksom på pollenrørene langs stilen med et mikroskop med UV-epifluorescens ved hjelp av 340-380 båndpass og 425 longpassfiltre.
2. DNA-ekstraksjon
- Prøve 2-3 blader i feltet om våren. Det anbefales å prøve bladene i unge stadier siden DNA oppnådd er av høyere kvalitet og lavere nivåer av fenoliske forbindelser sammenlignet med gamle blader.
- Trekk ut genomisk DNA etter trinnene som er beskrevet i et kommersielt tilgjengelig sett (se Materialkommand).
- Analyser mengden og kvaliteten på DNA-konsentrasjoner ved hjelp av UV-vis spektrofotometer (260 nm).
3. SS-allele identifikasjon
-
Sette opp PCR-reaksjoner
- Forbered en 50 ng/μL fortynning i destillert vann av hver DNA-ekstraksjonsprøve.
- Tin ut PCR-reagensene sakte og hold dem på is. La DNA-polymerasen stå i fryseren til det er nødvendig.
- Forbered forsterkningsreaksjonene ved hjelp av de forskjellige kombinasjonene av primere. Opprett PCR-reaksjonsblandingen ved å kombinere komponentene i tabell 1. Vortex PCR-reaksjonen blandes godt og fordel volumet som er angitt for de forskjellige kombinasjonene av primere til hver brønn av PCR-platen. Deretter tilsett 1 μL av DNA-fortynningen i hver brønn.
- Plasser PCR-platen i termohjulet og kjør det tilsvarende PCR-programmet som vises i tabell 1.
- Analyser de forsterkede fragmentene. Det er hovedsakelig to forskjellige måter å analysere PCR forsterkede fragmenter: kapillær elektroforese (CE) med fluorescerende-merkede primere eller som visualisere amplicons av agarose gel elektroforese med ikke-merkede primere.
-
Kapillær elektroforese
- For å klargjøre lastebufferen, bland 35 μL av deionisert formamid med 0,45 μL merket størrelsesstandard. Vortex reagensen for å blande godt, og dispenser deretter 35,5 μL inn i brønnen på leserplaten.
- Tilsett 1 μL av PCR-produktet i brønnen. I tillegg legger du til en dråpe mineralolje for å forhindre vannfordampning.
- Klargjør separasjonsplaten som legger til separasjonsbuffer.
- Bruk den kommersielle programvaren som følger med genanalysatoren (se Materialkommand). Opprett en ny prøveplate og lagre prøvenavnene for alle brønner på platen.
- Velg analysemetoden. I dette tilfellet denaturerer prøvene ved 90 °C i 120 s, injiseres ved 2,0 kV i 30 s, og separerer ved 6,0 kV i 35 min.
- Sett de to platene inn i genanalysatoren. Fyll kapillærmatrisen med destillert vann.
- Last den patenterte lineære gelatroen (LPA). Til slutt klikker du Kjør.
-
Gel elektroforese
- Forbered en 1% agarose gel legge 1,5 g molekylærbiologi grade agarose i 150 ml 1x TAE (Tris-acetat-EDTA) elektroforese kjører buffer (40 mM Tris, 20 mM eddiksyre, og 1 mM EDTA på pH 8.0). Oppløs agarose ved mikrobølgeovn oppvarming for 2-3 min.
- For å visualisere DNA, tilsett 4 μL av en nukleinsyreflekk (se Tabell over materialer) og bland forsiktig.
- Legg til en gelkam, med tilstrekkelige brønner for stiger, kontroller og prøver, i et gelbrett. Hell deretter sakte blandingen i midten av gelbrettet og unngå bobler.
- La gelen avkjøles i 30-45 min ved romtemperatur til gelen er helt størknet. Introduser gelen i elektroforesekammeret, fjern gelkammen og fyll kammeret med nok 1x TAE-buffer for å dekke gelen.
MERK: Kontroller plasseringen av gelen. Brønnene skal plasseres nær den negative polen siden negativt ladet DNA migrerer mot katoden. - Tilsett 5 μL lastingsbuffer (0,1 % (v/v) bromofenol blå) i PCR-produktene og bland godt.
- For å estimere størrelsen på båndene, last 5 μL DNA molekylvekt stige (se Tabell over materialer).
- Last prøvene inn i de ekstra brønnene i gelen.
- Når alle prøvene og DNA molekylvekt stigen er lastet, kjøre gel på 90 V for 1-1,5 timer, til den blå fargelinjen er ca 75% lengden på gelen.
- Visualiser båndene i en transilluminator for nukleinsyrer.
-
Kapillær elektroforese
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Pollineringsstudier i aprikos krever bruk av blomster på sen ballongstadiet en dag før antese (figur 1A). Dette stadiet regnes som den mest gunstige for både pollen og støvbærersamling, siden blomsterstrukturer er nesten modne, men anther dehiscence har ennå ikke skjedd. Dette forhindrer interferens av uønsket pollen, ikke bare av pollen fra samme blomst, men også fra andre blomster, siden de lukkede kronbladene hindrer ankomsten av insekter som bærer ekstern pollen. Pollenkornene blir lett siktet gjennom et fint nett (Figur 1B) fra dehisced anthers tidligere plassert på et stykke papir i 24 timer ved romtemperatur eller med liten ekstra varme. På samme måte hentes pistils fra blomster på ballongstadiet etter fjerning av kronblader, sepals og pollenbærere ved hjelp av pinsett eller negler (Figur 1C). Støvbærere kan være selv- og krysspollinerte med en fin børste (Figur 1D).
Hermaphroditic blomster av aprikos har fem mørke røde sepals, fem hvite kronblad(Figur 1A),en enkelt pistil (Figur 2A) og 25-30 pollenbærere. Pistil har tre hovedstrukturer: stigma, stil og eggstokk. Eggstokken har to eggstokker, og befruktning av minst en av dem er nødvendig for fruktinnstilling. Under pollinering overfører insekter, hovedsakelig bier, pollenkorn til stigmaet (figur 1A), hvor de spirer (figur 2B) innen 24 timer etter pollinering. Et pollenrør produseres fra hver spirende pollenkorn, som vokser gjennom støvesteinstrukturene for å nå eggstokken etter 3-4 dager og befrukte en av de to ovuleene etter rundt 7 dager. I selvkompatibel sorter der S allel av pollenkornet er den samme som en av de to pistils, pollen rør slutter å vokse i den øvre stilen, hindre befruktning (Figur 2C). Pollenrørene fra en kompatibel sort, med en annen S allele, kan imidlertid vokse gjennom stilen (Figur 2D), nå eggstokken (Figur 2E) og befrukte en av de to ovuleene.
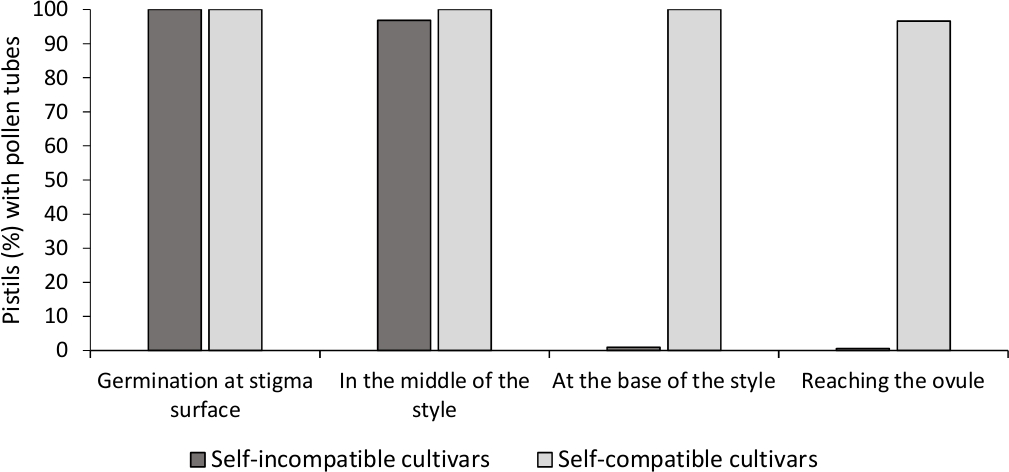
Analysen av in vitro pollen spiring viste god pollen levedyktighet i alle sorter analysert her, siden de fleste pollen rør var lengre enn lengden på pollen korn etter 24 h i kulturmediet. Germinerte pollenkorn ble observert på stigmaoverflaten (figur 2B) i støvler fra alle pollineringer, noe som indikerer tilstrekkelig pollinering (figur 3).
For å bestemme selvkompatibiliteten for hver sort, pollen rør atferd i selv- og krysspollineringer gjort i laboratoriekontrollerte forhold ble observert under fluorescens mikroskopi. Pollen rør vekst ble registrert langs stilen i alle pistils undersøkt. Cultivars ble ansett som selvkompatibel når pollen rør vekst ble arrestert langs stilen i de fleste selvpollinerte pistils (Figur 2C, Figur 3) og selvkompatibel når minst ett pollen rør nådde bunnen av stilen i de fleste av pistils undersøkt (Figur 2E, Figur 3).
Studien av S-locus ved PCR-analyse tillot å karakterisere S-genotypen til hver sort. For det Sførste ble S-allelene identifisert ved forsterkning av den første S-RNase intron ved hjelp av primers SRc-F/SRc-R (Tabell 2). Størrelsen på de forsterkede fragmentene ble analysert av kapillærelektroforese (figur 4A) og ble brukt til å klassifisere genotypene som ble analysert i deres tilsvarende inkompatibilitetsgruppe (I.G.) (Tabell 3).
Noen par alleler, som S1 og S7 eller S6 og S9,viste lignende fragmentstørrelser for den første intronen. Dermed ble differensieringen av disse allelene gjort ved å forsterke en region av den andre intron av RNase med primers Pru-C2 / PruC4R, SHLM1 / SHLM2 og SHLM3 / SHLM4 (Tabell 2). PruC2/PruC4R primer kombinasjonen ble brukt til å skille mellom S6 og S9. For S6ble det forsterket et fragment på 1300 bp, mens et fragment på rundt 700 bp ble observert for S9 allel ( figur4B, tabell 3). De spesifikke primers SHLM1/SHLM2 og SHLM3/SHLM4 forsterket et fragment på ca. 650 bp i S1 allele og 413 bp i S7 allele ( Figur4C, Tabell 3).
Primers AprFBC8-(F/R) som forsterker V2 og HVb variable regioner av SFB genet ble brukt til å skille Sc og S8 alleler siden begge allelene viser identiske RNase sekvens. S8 allelet viste et PCR-fragment på ca. 150 bp, mens et 500 bp-fragment tilsvarte Sc allele ( figur4D,tabell 3). Når S-genotypen ble bestemt for alle sortiene, ble selvkompatibele kultivar tildelt deres tilsvarende inkompatibilitetsgrupper basert på deres S-alleles (Tabell 3).
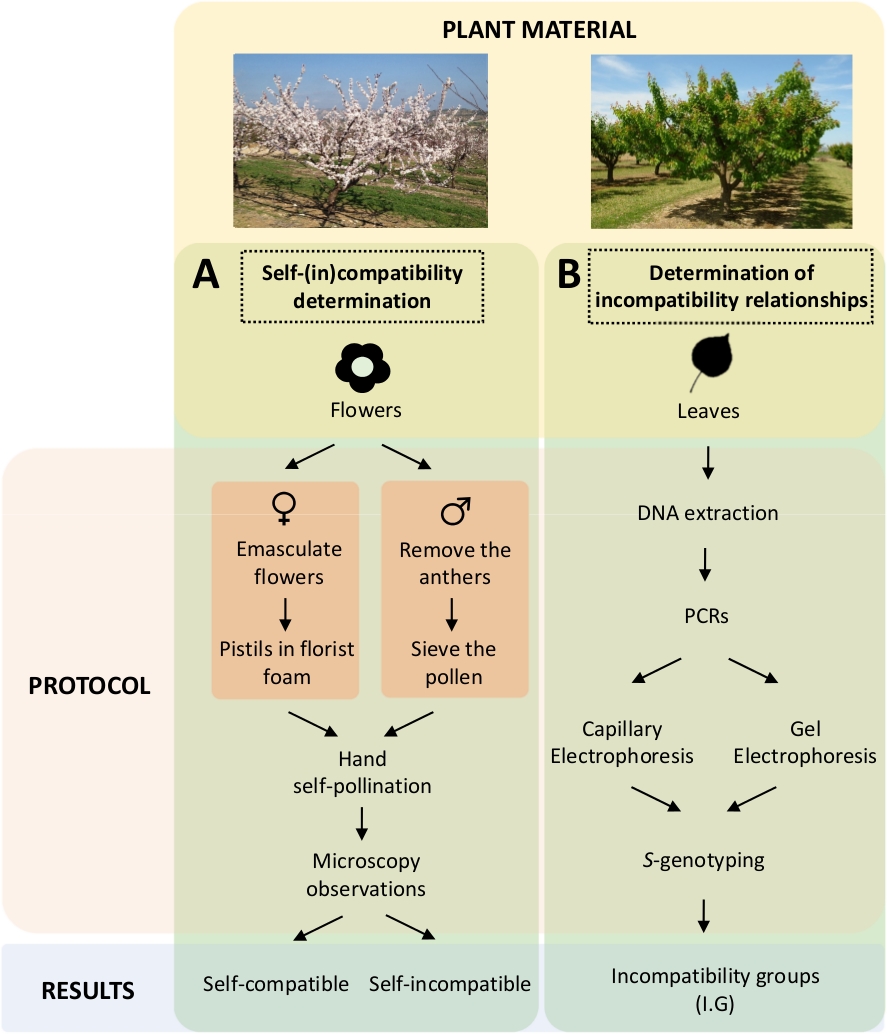
Denne tilnærmingen krever å bestemme selv-(i)kompatibiliteten til hver sortivar ved kontrollerte selv- og krysspollineringer i laboratoriet (Figur 5A) samtidig med karakterisering av S-genotype ved genetisk analyse ( Figur5B). Som et resultat kan pollineringskravene til hver sort og inkompatibilitetsforholdene mellom aprikoskulter bestemmes.

Figur 1. Eksperimentell satt opp for bestemmelse av selv-(i)kompatibilitet i aprikos.
(A)Blomster på ballongscenen (svarte piler) i feltet. (B) Sil av pollen korn ved hjelp av et fint nett. (C) Støvbærere plassert på blomsterhandlerskum i vann. (D)Håndpollinering av pistils ved hjelp av en pensel. Vennligst klikk her for å se en større versjon av dette tallet.

Figur 2. Diagrammatisk representasjon av gametophytic inkompatibilitet relasjoner i aprikos blomster.
(A)I Gametophytic Self-Incompatibility (GSI) spirer både kompatible og inkompatible pollenkorn på stigmaet. Pollenkornet bærer en av to S-alleler av den opprinnelige genotypen, i dette tilfellet enten S1 eller S2. Hvis S-allelet av pollen korn matcher en av de to S-alleles av støvsugeren, i dette tilfellet S1S3, pollen tube vekst er hemmet i den øvre en tredjedel av stilen. (B) Spiring av pollenkorn på stigmaoverflaten. (C)Pollen rør arrestert i stil som indikerer en uforenlig oppførsel. Pollenrørenevokser langs stilen. Pollenrøreneved foten av stilen indikerer en kompatibel oppførsel. Vektlinjer, 100 μm. Vennligst klikk her for å se en større versjon av dette tallet.

Figur 3. Representative resultater av pollen spiring og pollen tube vekst gjennom stil for selvkompatibel og selvkompatibel sorter etter selvpollinering.
Prosentandel av støvbærere med pollenkorn som spirer på stigmaoverflaten, med pollenrør halvveis i stilen, ved foten av stilen, og når eggløsningen. Vennligst klikk her for å se en større versjon av dette tallet.

Figur 4. PCR fragment forsterkning ved hjelp av fem primer par kombinasjoner for identifisering av S-alleles.
(A) Genanalysator utgang for SRc-(F/R) primere som viser størrelsen på de to forsterkede fragmenter av RNase første intron regionen tilsvarende S-alleles. (B) PCR-forsterkning ved hjelp av primers PruC2/ PruC4R for identifisering av S6 og S9 alleler. (C) PCR-produkter oppnådd ved hjelp av de spesifikke primerne SHLM1 og SHLM2 for differensiering av S1 allel og SHLM3 og SHLM4 for å skille mellom S7-allelet. (D) PCR forsterkning med AprFBC8-(F/R) primere for å identifisere Sc og S8 alleler. MI: 1 kb DNA Stige. MII: 100 bp DNA Stige. Vennligst klikk her for å se en større versjon av dette tallet.

Figur 5. Ordningen med eksperimentell design for å belyse selv- og inter-(in) kompatibilitetsrelasjoner i aprikoskuler.
(A) Arbeidsflyt for selvkompatibilitetsbestemmelse ved kontrollerte pollineringer i laboratoriet. (B)Arbeidsflyt av S-allele identifikasjon ved molekylære tilnærminger. Vennligst klikk her for å se en større versjon av dette tallet.
| PCR Master Mix | Termosyklusforhold | ||||||
| Komponenter | Endelig konsentrasjon | 15 μL reaksjon | Syklus trinn | Temperatur | Tid | Sykluser | |
| 10x NH4 Reaksjon buffer | 10x (andre) | 1,5 μL | Innledende denaturasjon | 94 °C | 3 min. | 1 | |
| 50 mM MgCl2 Oppløsning | 25 mM (andre) | 1,2 μL | Denaturing (Nær Denaturing) | 94 °C | 1 min. | 35 | |
| 100 mM dNTP | 2,5 mM (andre) | 0,6 μL | Annealing | 55 °C | 1 min. | ||
| Primer SRc-F | 10 μM | 0,6 μL | Forlengelsen | 72 °C (72 °F) | 3 min. | ||
| Primer SRc-R | 10 μM | 0,6 μL | Endelig utvidelse | 72 °C (72 °F) | 5 min. | 1 | |
| 500 U Taq DNA Polymerase | 0,5 U | 0,2 μL | 4 °C (4 °F) | Holde | |||
| H 2 O(andre) | 8,3 μL | ||||||
| Komponenter | Endelig konsentrasjon | 25 μL reaksjon | Syklus trinn | Temperatur | Tid | Sykluser | |
| 10x PCR-buffer | 10x (andre) | 2,5 μL | Innledende denaturasjon | 94 °C | 2 min. | 1 | |
| 5x Q-løsning | 5x (andre) | 5 μL | Denaturing (Nær Denaturing) | 94 °C | 10 s | 10 | |
| 100 mM dNTP | 2,5 mM (andre) | 0,5 μL | Annealing | 55 °C | 2 min. | ||
| Primer PruC2 | 10 μM | 0,2 μL | Forlengelsen | 68 °C (68 °F) | 2 min. | ||
| Primer C4R | 10 μM | 0,2 μL | Denaturing (Nær Denaturing) | 94 °C | 10 s | 25 | |
| 250 U Taq DNA Polymerase | 10 U (andre) | 0,13 μL | Annealing | 58 °C (58 °F) | 2 min. | ||
| H 2 O(andre) | 15,5 μL | Utvidelse* | 68 °C (68 °F) | 2 min. | |||
| Endelig utvidelse | 72 °C (72 °F) | 5 min. | 1 | ||||
| 4 °C (4 °F) | Holde | ||||||
| * med 10 s lagt hver syklus til 68 %C forlengelse trinn. | |||||||
| Komponenter | Endelig konsentrasjon | 25 μL reaksjon | Syklus trinn | Temperatur | Tid | Sykluser | |
| 10x PCR-buffer | 10x (andre) | 2,5 μL | Innledende denaturasjon | 94 °C | 2 min. | 1 | |
| 5x Q-løsning | 5x (andre) | 5 μL | Denaturing (Nær Denaturing) | 94 °C | 30 s | 35 | |
| 100 mM dNTP | 2,5 mM (andre) | 0,5 μL | Annealing | 62 °C (62 °F) | 1.5 min. | ||
| Primer SHLM1 | 10 μM | 0,2 μL | Forlengelsen | 72 °C (72 °F) | 2 min. | ||
| Primer SHLM2 | 10 μM | 0,2 μL | Endelig utvidelse | 72 °C (72 °F) | 5 min. | 1 | |
| 250 U Taq DNA Polymerase | 10 U (andre) | 0,13 μL | 4 °C (4 °F) | Holde | |||
| H 2 O(andre) | 15,5 μL | ||||||
| Komponenter | Endelig konsentrasjon | 20 μL reaksjon | Syklus trinn | Temperatur | Tid | Sykluser | |
| 5x PCR-buffer | 5x (andre) | 4 μL | Innledende denaturasjon | 98 °C | 30 s | 1 | |
| DNTP (andre) | 2,5 mM (andre) | 1,6 μL | Denaturing (Nær Denaturing) | 98 °C | 10 s | 35 | |
| Primer SHLM3 | 10 μM | 1 μL | Annealing | 51 °C (51 °F) | 30 s | ||
| Primer SHLM4 | 10 μM | 1 μL | Forlengelsen | 72 °C (72 °F) | 1 min. | ||
| 100 U DNA Polymerase | 5 U (andre) | 0,2 μL | Endelig utvidelse | 72 °C (72 °F) | 5 min. | 1 | |
| H 2 O(andre) | 12,4 μL | 4 °C (4 °F) | Holde | ||||
| Komponenter | Endelig konsentrasjon | 25 μL reaksjon | Syklus trinn | Temperatur | Tid | Sykluser | |
| 10x PCR-buffer | 10x (andre) | 2,5 μL | Innledende denaturasjon | 94 °C | 2 min. | 1 | |
| 100 mM dNTP | 2,5 mM (andre) | 2 μL | Denaturing (Nær Denaturing) | 94 °C | 30 s | 35 | |
| Primer FBC8-F | 10 μM | 1 μL | Annealing | 55 °C | 1.5 min. | ||
| Primer FBC8-R | 10 μM | 1 μL | Forlengelsen | 72 °C (72 °F) | 2 min. | ||
| 250 U Taq DNA Polymerase | 10 U (andre) | 0,125 μL | Endelig utvidelse | 72 °C (72 °F) | 5 min. | 1 | |
| H 2 O(andre) | 17,4 μL | 4 °C (4 °F) | Holde | ||||
Tabell 1. Reaksjons- og sykkelforhold for ulike primerkombinasjoner som brukes i denne protokollen.
| Primere | Sekvens | Referanse |
| Src-F (andre) | 5'-CTCGCTTTCCTTGTTCTTGC-3' | 18 |
| SRc-R (andre) | 5'-GGCCATTGTTGCACCCCTTG-3' | 18 |
| Pru-C2 (andre) | 5'-CTTTGGCCAAGTAATTATTCAAACC-3' | 35 |
| Pru-C4R (andre) | 5'-GGATGTGGTACGATTGAAGCG-3' | 35 |
| Shlm1-F (andre) | 5'-GGTGGAGGTGATAAGGTAGCC-3' | 17 |
| Shlm2-R (andre) | 5'-GGCTGCATAAGGAAGCTGTAGG-3' | 17 |
| Shlm3-F (andre) | 5'-TATATCTTACTCTTTGGC-3' | 17 |
| Shlm4-R (andre) | 5'-CACTATGATAATGTGTATG-3' | 17 |
| AprilFBC8-F | 5'-CATGGAAAAAGCTGACTTATGG-3' | 26 |
| AprilFBC8-R | 5'-GCCTCTAATGTCATCTACTCTTAG-3' | 26 |
Tabell 2. Primere som brukes i denne protokollen, sekvens og referanse for S-genotype karakterisering i Prunus armeniaca.
| Kultivaren | SRc-(F/R) (bp) | PruC2/PruC4R (bp) | SHLM1/SHLM2 (bp) | SHLM3/SHLM4 (bp) | AprFBC8-(F/R) (bp) | S-Genotype (andre) | Inkompatibilitetsgruppe (I.G) |
| Lurer på barneseng8 | 420, 420 | 749, 1386 | S6S9 (andre) | Viii | |||
| Magisk barneseng8 | 334, 420 | 749 | S2S9 (andre) | Xx | |||
| Gullstrike8 | 334, 420 | 749 | S2S9 (andre) | - | |||
| T06917 (andre) | 334, 408 | 650 | S1S2 (andre) | I | |||
| T12017 (andre) | 334, 408 | 650 | S1S2 (andre) | - | |||
| C-6 (andre) | 334, 408 | 413 | S 2 S7 (andre) | Iv | |||
| Cooper Barneseng8 | 274, 408 | 650 | S1S3 (andre) | Xviii | |||
| Apriqueen (andre) | 358, 358 | 500 | ScSc | - | |||
| Bergecot8 (Andre) | 334, 358 | 500 | S2Sc | - | |||
| Vårrødme8 | 274, 358 | 150 | S 3 S8 (andre) | Xxi |
Tabell 3. S-genotyping av aprikos kultivar med fem primer par som brukes i denne protokollen og inkompatibilitet gruppe tildeling. S De forskjellige polymerase kjedereaksjonsproduktstørrelsene til S-allelesforsterket ved hjelp av SRc-(F/R), PruC2/PruC4R, SHLM1/ SHLM2, SHLM3/SHLM4 og AprFBC8-(F/R) primere er vist i tabellen.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Tradisjonelt var de fleste kommersielle aprikoseuropeiske sorter selvkompatibele36. Likevel har bruken av nordamerikanske selvkompatibele sorter som foreldre i avlsprogrammer de siste tiårene resultert i utgivelsen av et økende antall nye selvkompatibele kultigjenstander med ukjente pollineringskrav7,8,37. Dermed er fastsettelsen av selv- og inter-(in)kompatibilitetsforhold i aprikoskuler stadig viktigere. Dette fremheves i de områdene hvor vinteren chilling er avtagende, siden høy år til år variasjoner i blomstringstidspunktet hindrer tilfeldighet i blomstring av sorter og deres pollenizers i mange tilfeller, spesielt i sorter med høye skremmende krav38. Metodikken som er beskrevet her, som kombinerer håndpollinering, mikroskopi og genetiske analyser, har vært svært nyttig for å bestemme selvkompatibiliteten til hver sort og for å etablere sine potensielle pollinizer-sorter.
Pollineringskrav kan bestemmes gjennom feltkontrolleksperimenter i frukthageforhold11,,39. Imidlertid kan utstillingen til eksterne faktorer, inkludert meteorologiske ugunstige forhold, forårsake pollineringsfeil10, noe som kan føre til feilaktige diagnoser av selvinkompatibilitet. Metodikken som er beskrevet her, gjør det mulig å evaluere selvkompatibilitet mer nøyaktig ved mikroskopiobservasjoner av håndpollinerte blomster i laboratoriekontrollerte forhold, og unngår miljøpåvirkning. Videre gjør denne tilnærmingen det mulig å analysere et høyere antall sorter per år, siden bare et lite antall blomster er nødvendig i stedet for flere voksne trær for hver sort som kreves i felteksperimenter40.
Inkompatibilitetsrelasjoner kan etableres som kombinerer håndpollinering og mikroskopi14. Imidlertid kan pollineringer bare utføres i en kort periode i blomstringsperioden om våren, og voksne trær i nærheten av laboratoriet er nødvendig, siden levetiden til blomstene samlet er svært kort. Dermed er antall inkompatibilitetsforhold som kan analyseres av kontrollerte håndpollineringer i hver sesong svært lavt. Karakteriseringen av genene kodet av S-locus har gjort det mulig å utvikle PCR-baserte metoder for S-allele genotyping18,41. Denne tilnærmingen akselererer S-allele identifikasjon siden det ikke krever blomster, og forsøkene kan utføres med noen vegetative vev42. Dette forlenger perioden der plantemateriale, vanligvis unge blader, kan samles43. Videre kan bladene være lyofilisert eller frosset, slik at analysen kan gjøres når som helst på året, i motsetning til pollineringer som bare kan gjøres på friske blomster i blomstringssesongen44. En ekstra fordel er at blader kan samles inn fra unge trær selv før du går inn blomstrende alder, lette innsamling av prøver og tidlig innhenting av resultater45.
Den genetiske analysen gir en bedre differensiering av selvinkompatibilitetslyser siden det gir nøyaktige resultater av forsterkede fragmentstørrelser21,,46. Hittil har trettitre S-alleler blitt identifisert i aprikos12,18,20,21,22,23,24, som har lov til å etablere 36 inkompatibilitetsgrupper basert på S-genotype8,9,17,25,26,27. På den annen side er en ulempe med denne metodikken at forskjellige alleler i samme rekkeviddestørrelse eller mutasjoner kan feilaktig identifiseres som samme allel. Dermed er Sc og S8 alleler identiske for RNase-sekvensen, men en 358-bp innsetting finnes i SFB-genet sc19. På samme måte er den første intron-regionen i allelene S1 og S7 identiske og kan ikke skilles ved hjelp av primerne SRc-F/SRc-R. I tillegg har flere homologier, som S6 og S528 eller S20 og S S55, og S7, S13 (EF062341) og S4617, blitt funnet fordi noen av disse allelene har blitt delvis sekvensert eller ved feil under PCR-forsterkning, og derfor er det nødvendig med ytterligere arbeid for å skille dem riktig.
PCR analyse og S-RNase sekvensering er tilstrekkelig for å etablere inkompatibilitet relasjoner gjennom identifisering av S-allelesog tildeling av sorter i deres tilsvarende Incompatibility Group8,17,26,27. Denne metodikken har imidlertid begrensningen av å forhindre fastsettelse av selvkompatibiliteten for bestemte aprikoskuler. Selvkompatibilitet (SC) har vært knyttet til bestemte S-alleleri andre Prunus arter47, som mandel (Sf)48,,49 eller søt kirsebær (S4')50,51. Men i aprikos, Sc allele, som har vært knyttet til SC21, kan feilaktig identifiseres som S8, en selvkompatibel allel19,22, og mulige mutasjoner ikke knyttet til S locus, som M-locus12,52, conferring SC har blitt identifisert. Nylig har M-locus blitt genotyped ved hjelp av SSR-markører12. Derfor trenger den genetiske identifiseringen av SC for aprikos genotyper videre forskning, og for å unngå feil på grunn av faktorer som ikke er knyttet til S locus, i dette arbeidet karakterisering av selv-(i)kompatibilitet har blitt bestemt også ved fenotyping oppførselen til pollen rør gjennom pistiled blomster.
Metodikken som er beskrevet her i å kombinere fastsettelsen av selv-(i)kompatibilitet ved hånd-pollineringer i laboratorieforhold med påfølgende observasjon av oppførselen til pollenrør i pistil av Skontrollerte selvpollineringer under fluorescensmikroskopi og identifisering av S-genotype ved PCR-analyse gjør det mulig å etablere pollineringskravene til aprikoskuler. Dette gir en verdifull informasjon for dyrkere og oppdrettere, siden det gjør det mulig å etablere inkompatibilitetsforhold mellom sorter å velge egnede pollinizers i utformingen av nye frukthager samt å velge passende foreldre til å designe nye kors i aprikos avlsprogrammer.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Forfatterne har ingenting å avsløre.
Acknowledgments
Denne forskningen ble finansiert av Ministerio de Ciencia, Innovación y Universidades-European Regional Development Fund, Eu (AGL2016-77267-R, og AGL2015-74071-JIN); Instituto Nacional de Investigación y Tecnología Agraria y Alimentaria (RFP2015-00015-00, RTA2017-00003-00); Gobierno de Aragón-European Social Fund, Eu (Grupo Consolidado A12_17R), Fundación Biodiversidad og Agroseguro S.A.
Materials
| Name | Company | Catalog Number | Comments |
| Agarose D1 Low EEO | Conda | 8010.22 | |
| BIOTAQ DNA Polymerase kit | Bioline | BIO-21060 | |
| Bright field microscope | Leica Microsystems | DM2500 | |
| CEQ System Software | Beckman Coulter | ||
| DNeasy Plant Mini Kit | QIAGEN | 69106 | |
| dNTP Set, 4 x 25 µmol | Bioline | BIO-39025 | |
| GenomeLab DNA Size Standard Kit - 400 | Beckman Coulter | 608098 | |
| GenomeLab GeXP Genetic Analysis System | Beckman Coulter | ||
| GenomeLab Separation Buffer | Beckman Coulter | 608012 | |
| GenomeLab Separation Gel LPA-1 | Beckman Coulter | 391438 | |
| HyperLadder 100bp | Bioline | BIO-33029 | |
| HyperLadder 1kb | Bioline | BIO-33025 | |
| Image Analysis System | Leica Microsystems | ||
| Molecular Imager VersaDoc MP 4000 system | Bio-Rad | 170-8640 | |
| NanoDrop One Spectrophotometer | Thermo Fisher Scientific | 13-400-518 | |
| pH-Meter BASIC 20 | Crison | ||
| Phusion High-Fidelity PCR Kit | Thermo Fisher Scientific | F553S | |
| Power Pack P 25 T | Biometra | ||
| Primer Forward | Isogen Life Science | ||
| Primer Reverse | Isogen Life Science | ||
| Quantity One Software | Bio-Rad | ||
| Stereoscopic microscope | Leica Microsystems | MZ-16 | |
| Sub-Cell GT | Bio-Rad | ||
| SYBR Safe DNA Gel Stain | Thermo Fisher Scientific | S33102 | |
| T100 Thermal Cycler | Bio-Rad | 1861096 | |
| Taq DNA Polymerase | QIAGEN | 201203 | |
| Vertical Stand Autoclave | JP Selecta |
References
- Silva, N. F., Goring, D. R. Mechanisms of self-incompatibility in flowering plants. Cellular and Molecular Life Sciences. 58, 1988-2007 (2001).
- Charlesworth, D., Vekemans, X., Castric, V., Glémin, S. Plant self-incompatibility systems: A molecular evolutionary perspective. New phytologist. 168, 61-69 (2005).
- Tao, R., et al. Identification of stylar RNases associated with gametophytic self-incompatibility in almond (Prunus dulcis). Plant and Cell Physiology. 38, 304-311 (1997).
- Ushijima, K., et al. Structural and transcriptional analysis of the self-incompatibility locus of almond: Identification of a pollen-expressed F-box gene with haplotype-specific polymorphism. The Plant cell. 15, 771-781 (2003).
- Bedinger, P. A., Broz, A. K., Tovar-Mendez, A., McClure, B. Pollen-Pistil Interactions and Their Role in Mate Selection. Plant Physiology. 173, 79-90 (2017).
- Guerra, M. E., Rodrigo, J. Japanese plum pollination: A review. Scientia Horticulturae. 197, 674-686 (2015).
- Zhebentyayeva, T., Ledbetter, C., Burgos, L., Llacer, G. Apricot. Fruit Breeding. Badenes, M. L., Byrne, D. , Springer. Boston, MA. 415-458 (2012).
- Herrera, S., Lora, J., Hormaza, J. I., Herrero, M., Rodrigo, J. Optimizing Production in the New Generation of Apricot Cultivars: Self-incompatibility, S-RNase Allele Identification, and Incompatibility Group Assignment. Frontiers in Plant Science. 9, 527 (2018).
- Egea, J., Burgos, L. Detecting Cross-incompatibility of Three North American Apricot Cultivars and Establishing the First Incompatibility Group in Apricot. Journal of the American Society for Horticultural Science. 121, 1002-1005 (1996).
- Rodrigo, J., Herrero, M. Effects of pre-blossom temperatures on flower development and fruit set in apricot. Scientia Horticulturae. 92, 125-135 (2002).
- Julian, C., Herrero, M., Rodrigo, J. Flower bud differentiation and development in fruiting and non-fruiting shoots in relation to fruit set in apricot (Prunus armeniaca). Trees. 24, 833-841 (2010).
- Muñoz-Sanz, J. V., Zuriaga, E., López, I., Badenes, M. L., Romero, C. Self-(in)compatibility in apricot germplasm is controlled by two major loci, S and M. BMC Plant Biology. 17, 82 (2017).
- Burgos, L., Berenguer, T., Egea, J. Self- and Cross-compatibility among Apricot Cultivars. HortScience. 28, 148-150 (1993).
- Rodrigo, J., Herrero, M. Evaluation of pollination as the cause of erratic fruit set in apricot "Moniqui". Journal of Horticultural Science. 71, 801-805 (1996).
- Milatović, D., Nikolić, D., Krška, B. Testing of self-(in)compatibility in apricot cultivars from European breeding programmes. Horticultural Science. 40 (2), 65-71 (2013).
- Milatović, D., Nikolić, D., Fotirić-Aksić, M., Radović, A. Testing of self-(in)compatibility in apricot cultivars using fluorescence microscopy. Acta Scientiarum Polonorum, Hortorum Cultus. 12 (6), 103-113 (2013).
- Herrera, S., Rodrigo, J., Hormaza, J. I., Lora, J. Identification of Self-Incompatibility Alleles by Specific PCR Analysis and S-RNase Sequencing in Apricot. Int J Mol Sci. 19, 3612 (2018).
- Romero, C., et al. Analysis of the S-locus structure in Prunus armeniaca L. Identification of S-haplotype specific S-RNase and F-box genes. Plant Molecular Biology. 56, 145-157 (2004).
- Halász, J., Pedryc, A., Hegedus, A. Origin and dissemination of the pollen-part mutated SC haplotype which confers self-compatibility in apricot (Prunus armeniaca). New Phytologist. 176, 792-803 (2007).
- Halász, J., Hegedus, A., Hermán, R., Stefanovits-Bányai, É, Pedryc, A. New self-incompatibility alleles in apricot (Prunus armeniaca L.) revealed by stylar ribonuclease assay and S-PCR analysis. Euphytica. 145, 57-66 (2005).
- Vilanova, S., Romero, C., Llacer, G., Badenes, M. L., Burgos, L. Identification of Self-(in)compatibility Alleles in Apricot by PCR and Sequence Analysis. Journal of the American Society for Horticultural Science. 130, 893-898 (2005).
- Feng, J., et al. Detection and transcript expression of S-RNase gene associated with self-incompatibility in apricot (Prunus armeniaca L.). Molecular Biology Reports. 33, 215-221 (2006).
- Zhang, L., et al. Identification of self-incompatibility (S-) genotypes of Chinese apricot cultivars. Euphytica. 160, 241-248 (2008).
- Wu, J., et al. Identification of S-haplotype-specific S-RNase and SFB alleles in native Chinese apricot (Prunus armeniaca L). Journal of Horticultural Science and Biotechnology. 84, 645-652 (2009).
- Szabó, Z., Nyéki, J. Blossoming, fructification and combination of apricot varieties. Acta Horticulturae. 293, 295-302 (1991).
- Halász, J., Pedryc, A., Ercisli, S., Yilmaz, K. U., Hegedűs, A. S-genotyping supports the genetic relationships between Turkish and Hungarian apricot germplasm. Journal of the American Society for Horticultural Science. 135, 410-417 (2010).
- Lachkar, A., et al. Identification of self-(in)compatibility S-alleles and new cross-incompatibility groups in Tunisian apricot (Prunus armeniaca L.) cultivars. The Journal of Horticultural Science and Biotechnology. 88, 497-501 (2013).
- Pérez-Pastor, A., Ruiz-Sánchez, M. C., Domingo, R., Torrecillas, A. Growth and phenological stages of Búlida apricot trees in South-East. Agronomie. 24, 93-100 (2004).
- Williams, J. H., Friedman, W. E., Arnold, M. L. Developmental selection within the angiosperm style: using gamete DNA to visualize interspecific pollen competition. Proceedings of the National Academy of Sciences of the United States of America. 96, 9201-9206 (1999).
- Julian, C., Herrero, M., Rodrigo, J. Anther meiosis time is related to winter cold temperatures in apricot (Prunus armeniaca L.). Environmental and Experimental Botany. 100, 20-25 (2014).
- Guerra, M. E., López-Corrales, M., Wünsch, A., Rodrigo, J. Lack of Fruit Set Caused by Ovule Degeneration in Japanese Plum. Journal of the American Society for Horticultural Science. 136 (6), 375-381 (2011).
- Guerra, M. E., Wünsch, A., López-Corrales, M., Rodrigo, J. Flower Emasculation as the Cause for Lack of Fruit Set in Japanese Plum Crosses. Journal of the American Society for Horticultural Science. 135 (6), 556-562 (2010).
- Hormaza, J. I., Pinney, K., Polito, V. S. Correlation in the tolerance to ozone between sporophytes and male gametophytes of several fruit and nut tree species (Rosaceae). Sexual Plant Reproduction. 9, 44-48 (1996).
- Alcaraz, M. L., Hormaza, J. I., Rodrigo, J. Pistil Starch Reserves at Anthesis Correlate with Final Flower Fate in Avocado (Persea americana). PLOS ONE. 8 (10), 78467 (2013).
- Tao, R., et al. Molecular typing of S-alleles through Identification, Characterization and cDNA cloning for S-RNases in Sweet Cherry. Journal of the American Society for Horticultural Science. 124, 224-233 (1999).
- Burgos, L., et al. The self-compatibility trait of the main apricot cultivars and new selections from breeding programmes. Journal of Horticultural Science. 72, 147-154 (1997).
- Hormaza, J. I., Yamane, H., Rodrigo, J. Apricot. Genome Mapping and Molecular Breeding in Plants, Volume 4 Fruits and Nuts. Kole, C. , Springer. New York, NY. 171-187 (2007).
- Benmoussa, H., Ghrab, M., Ben Mimoun, M., Luedeling, E. Chilling and heat requirements for local and foreign almond (Prunus dulcis Mill.) cultivars in a warm Mediterranean location based on 30 years of phenology records. Agricultural and Forest Meteorology. 239, 34-46 (2017).
- Rodrigo, J., Herrero, M., Hormaza, J. I. Pistil traits and flower fate in apricot (Prunus armeniaca). Annals of Applied Biology. 154, 365-375 (2009).
- Williams, R. R. Techniques used in fruit-set experiments. Towards Regulated Cropping. Williams, R. R., Wilson, D. , Grower Books. London. 57-61 (1970).
- Sutherland, B. G., Robbins, T. P., Tobutt, K. R. Primers amplifying a range of Prunus S-alleles. Plant Breeding. 123, 582-584 (2004).
- Murray, M. G., Thompson, W. F. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Research. 8, 4321-4325 (1980).
- Porebski, S., Bailey, L. G., Baum, B. R. Modification of a CTAB DNA Extraction Protocol for Plants Containing High Polysaccharide and Polyphenol Components. Plant Molecular Biology Reporter. 15 (1), 8-15 (1997).
- Rogers, S. O., Bendich, A. J. Extraction of DNA from milligram amounts of fresh, herbarium and mummified plant tissues. Plant Molecular Biology. 5 (2), 69-76 (1985).
- Hormaza, J. I. Molecular characterization and similarity relationships among apricot (Prunus armeniaca L.) genotypes using simple sequence repeats. Theoretical and Applied Genetics. 104, 321-328 (2002).
- Sonneveld, T., Tobutt, K. R., Robbins, T. P. Allele-specific PCR detection of sweet cherry self-incompatibility (S) alleles S1 to S16 using consensus and allele-specific primers. Theoretical and Applied Genetics. 107, 1059-1070 (2003).
- Hegedus, A., Lénárt, J., Halász, J. Sexual incompatibility in Rosaceae fruit tree species: molecular interactions and evolutionary dynamics. Biologia Plantarum. 56 (2), 201-209 (2012).
- Fernández i Martí, A., Gradziel, T. M., Socias i Company, R. Methylation of the Sf locus in almond is associated with S-RNase loss of function. Plant Molecular Biology. 86, 681-689 (2014).
- Company, R. S. i, Kodad, O., Martí, A. F. i, Alonso, J. M. Mutations conferring self-compatibility in Prunus species: From deletions and insertions to epigenetic alterations. Scientia Horticulturae. 192, 125-131 (2015).
- Boskovic, R., Tobutt, K. R. Correlation of stylar ribonuclease zymograms with incompatibility alleles in sweet cherry. Euphytica. 90, 245-250 (1996).
- Cachi, A. M., Wünsch, A. S-genotyping of sweet cherry varieties from Spain and S-locus diversity in Europe. Euphytica. 197 (2), 229-236 (2014).
- Zuriaga, E., et al. An S-locus Independent Pollen Factor Confers Self-Compatibility in "Katy" Apricot. PLoS ONE. 8 (1), 53947 (2013).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}