

Summary
Flüoresan mikroskopi ile öz-genotiyon un belirlenmesini birleştiren kayısı(Prunus armeniaca L.) çeşitlerinin tozlaşma gereksinimlerini niçin pcr analizi ile s-genotipinin tanımlanması ile biraraya getiren bir metodoloji salıyoruz.
Abstract
Rosaceae'de kendi kendine uyumsuzluk, esas olarak multiallelik lokus S tarafından kontrol edilen Gametofitik Kendi Kendine Uyumsuzluk Sistemi (GSI) tarafından belirlenir. Kayısıda, önemli sayıda yeni çeşidinin serbest bırakılması bilinmeyen tozlaşma gereksinimlerine sahip çeşitlerin artmasına yol açtığı ndan, öz ve inter-(in) uyumluluk ilişkilerinin belirlenmesi giderek daha önemlidir. Burada, el-polenler ve mikroskopi ile öz-(in)uyumluluğunun belirlenmesini PCR analizi ile S-genotipinin tanımlanması ile birleştiren bir metodolojiyi açıklıyoruz. Uyumluluk tespiti için, her bir çeşitten balon aşamasında çiçekler toplanmış, laboratuvarda elle tozlaşmış, sabit lenmiş ve floresan mikroskopisi altında polen tüpü davranışlarının gözlemi için anilin mavisi ile boyanmıştır. Çeşitleri arasında uyumsuzluk ilişkilerinin kurulması için, her çeşitdna genç yapraklarından elde edildi ve S-alel PCR tarafından tespit edildi. Bu yaklaşım, yeni meyve bahçeleri tasarımında uygun tozlaştırıcıları seçmek ve Üreme programlarında uygun ebeveynleri seçmek için değerli bilgiler sağlayan çeşitler arasındaki uyumsuzluk ilişkilerini açıklığa kavuşturmaya ve uyumsuzluk ilişkileri ortaya çıkarmasına olanak sağlar.
Introduction
Kendi kendine uyumsuzluk kendi kendine tozlaşma önlemek ve1outcrossing teşvik çiçekli bitkilerin bir stratejidir. Rosaceae'de bu mekanizma, esas olarak multiallelic locus S2tarafından kontrol edilen Gametofitik Kendi Kendine Uyumsuzluk Sistemi (GSI) tarafından belirlenir. Tarzında, RNase geni S-stylar determinant kodlar, bir RNase3, Bir F-box protein, S-polenbelirleyici belirler, SFB gen4tarafından kodlanmış iken . Kendi kendine uyumsuzluk etkileşimi ovule döllenme engelleyen tarzı boyunca polen tüpü büyüme inhibisyonu yoluyla gerçekleşir5,6.
Kayısı, bir varietal yenileme son yirmi yılda dünya çapında yer almıştır7,8. Yeni çeşitlerin önemli bir sayı bu giriş, farklı kamu ve özel üreme programlarından, bilinmeyen tozlaşma gereksinimleri ile kayısı çeşitlerinin artmasına neden olmuştur8.
Kayısıda tozlaşma gereksinimlerini belirlemek için farklı metodolojiler kullanılmıştır. Alanında, kendini-(in)uyumluluk kafesli ağaçlarda veya hadım edilmiş çiçeklerde kontrollü pollinations tarafından kurulmuş ve daha sonra meyve seti yüzdesi kayıt9,10,11,12. Buna ek olarak, kontrollü pollinations çiçek yarı-in vivo kültür ve floresan mikroskopi8altında polen tüp davranış analizi tarafından laboratuvarda yapılmıştır 8,13,14,15,16,17. Son zamanlarda, pcr analizi ve dizileme gibi moleküler teknikler, RNase ve SFB genlerinin çalışmasına dayalı uyumsuzluk ilişkilerinkarakterizasyonu izin var18,19. Kayısıda, otuz üç S-alel bildirilmiştir (S1 - S20, S22 - S30, S52, S53, Sv, Sx), kendi kendine uyumluluk ile ilgili bir alel dahil (Sc)12,18,20,21,22,23,24. Şimdiye kadar, 26 uyumsuzluk grupları S-genotip8,9,17,,25,26,27göre bu tür bıçaklanmış edilmiştir. Aynı Salellerine sahip çeşitleri birbirine uyumsuz, en az bir farklı S-alel ile ve dolayısıyla farklı uyumsuz gruplara ayrılan çeşitleri birbirine uyumludur.
Kayısı çeşitlerinin tozlaşma gereksinimlerini tanımlamak için, flüoresan mikroskobu ile öz-(in)uyumluluk tayinini, kayısı çeşitlerinde PCR analizi ile S-genotipinin tanımlanması ile birleştiren bir metodoloji yi tanımlıyoruz. Bu yaklaşım uyumsuzluk gruplarının oluşturulmasına ve tarikatlar arasındaki uyumsuzluk ilişkilerinin açıklığa kavuşturulmasına olanak sağlar.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. Kendi kendine uyumluluk tayini
- Alandaki çiçekleri tadına var. İstenmeyen önceki tozlaşmayıFigure 1Aönlemek için, kayısı28için BBCH ölçeğinde 58.
-
Laboratuvarda kendi kendine ve çapraz tozlaşmalara
- Balon aşamasında çiçek anthers çıkarın ve laboratuvar sıcaklığında kuruması için bir kağıt parçası üzerine yerleştirin.
- 24 saat sonra polen tanelerini ince bir örgü (0,26 mm)(Şekil 1B)kullanarak elek.
- Her kendi kendine tozlaşma ve çapraz tozlaşma için aynı balon aşamasında 30 çiçekten oluşan bir grubu hadım edin ve pistilleri laboratuvar sıcaklığında sudaki çiçekçi köpüğüne yerleştirin(Şekil 1C).
- El emasculation sonra aynı çati 24 saat çiçekpolen ile bir boya fırçası yardımıyla pistils tozlaşma. Buna ek olarak, kontrol olarak uyumlu bir tozlaştırıcı çiçeklerinden polen ile her çeşit pistils başka bir dizi tozlaşma (Şekil 1D).
- 72 saat sonra, etanol/asetik asit (3:1) fiksatif çözeltisi içinde, 4 °C29'daen az 24 saat boyunca pistilleri düzeltin. Daha sonra fiksatif atın ve örneklertamamen çözelti içinde batırılmış olduğundan emin% 75 etanol ekleyin. Numuneler bu çözeltide 4 °C'de8,17,30,,31,32kullanıma kadar saklanabilir.
-
İn vitro polen çimlenmesi ile polen canlılığının değerlendirilmesi
- Çimlenme ortamını hazırlamak için, ağırlık 25 g sakaroz, 0.075 g borik asit (H3BO3)ve 0.075 g kalsiyum nitrat (Ca(NO3)2)33.
- 250 mL distile su içinde orta bileşenleri ekleyin ve tamamen çözünür.
- 2 g agarose ekleyerek ortamı katılatın ve girdap ile karıştırın.
- Bir pH metre kullanarak ortamın pH'ını kontrol edin ve NaOH veya HCl çözeltisi ile değeri 7.0 olarak ayarlayın.
- Otoklav karışımı orta sterilize etmek.
- Otoklavlama sonra, orta soğutun ve steril bir laminar akış başlık Petri yemekleri içine dağıtmak.
- Katılaşmış polen çimlenme ortamında kontrollü polenler için kullanılan aynı çeşitlerin polen tanelerini dağıtır ve 24 saat6'dansonra mikroskop altında gözlemleyin.
NOT: Laminar akış başlığını sterilize etmek için yüzeyi %70 etanol ile temizleyin ve 10 dakika boyunca UV lambasını açın. - Petri yemeklerini kullanıma kadar 4 °C'de buzdolabında saklayın.
-
Mikroskopi gözlemleri
- Pistilleri 1 saat boyunca distile suyla üç kez yıkayın ve 4 °C'de %5 sodyum sülfit bırakın. 24 saat sonra, 1 kg/cm2 sodyum sülfit 10 dakika boyunca dokuları yumuşatmak için onları otoklav34.
- Bir cam slayt üzerinde otoklavlı pistils yerleştirin ve, bir neşter yardımıyla, polen tüpleri daha iyi bir görselleştirme elde etmek için yumurtalık etrafında trichomes kaldırın. Daha sonra pistibir camla ezin.
- %0.1 (v/v) anilin mavisi lekesini hazırlayın: 0,1 mL'lik anilin mavisinin 0,1 mL'lik 0,1 N potasyum fosfat tribasic karışımını (K3PO4). Polen tüpü büyümesi sırasında kalkoz depozisyonlarını lekelemek için preparatların üzerine bir damla aniline mavisi uygulayın.
- 340-380 bandpass ve 425 longpass filtreleri kullanarak UV epifloresans ile bir mikroskop ile tarzı boyunca polen tüpleri gözlemleyin.
2. DNA çıkarma
- Örnek 2-3 ilkbaharda alanda yaprakları. Elde edilen DNA eski yapraklara göre fenolik bileşiklerin daha yüksek kalite ve daha düşük seviyelerde olduğundan, genç evrelerde yaprakların örnekalınması tavsiye edilir.
- Ticari olarak kullanılabilen bir kitte açıklanan adımları izleyerek Genomik DNA ayıklayın (Bkz. Malzeme Tablosu).
- UV-vis spektrofotometre (260 nm) kullanarak DNA konsantrasyonlarının miktarını ve kalitesini analiz edin.
3. S-alel tanımlama
-
PCR Reaksiyonlarının Ayarlanması
- Her DNA çıkarma örneğinin distile suyunda 50 ng/μL seyreltme hazırlayın.
- PCR reaktiflerini yavaşça eritin ve buzda tutun. DNA polimeraz'ı ihtiyaç duyulana kadar dondurucuda bırakın.
- Astarların farklı kombinasyonlarını kullanarak amplifikasyon reaksiyonlarını hazırlayın. Tablo 1'dekibileşenleri birleştirerek PCR reaksiyon karışımıoluşturun. Vortex PCR reaksiyonu iyi karıştırın ve PCR plaka her kuyuya astar farklı kombinasyonları için belirtilen hacmi dağıtmak. Daha sonra, her kuyuda 1 μL DNA seyreltme ekleyin.
- PCR plakasını termocycler'a yerleştirin ve Tablo 1'degösterilen ilgili PCR programını çalıştırın.
- Güçlendirilmiş parçaları analiz edin. PCR amplifiye parçaları analiz etmek için başlıca iki farklı yolu vardır: kapiller elektroforezi (CE) floresan etiketli astarlar ile veya etiketli olmayan astar ile agarose jel elektroforez amfisi görselleştirmek gibi.
-
Kılcal Elektroforez
- Yükleme tamponu hazırlamak için 35 μL deiyonize formamid'i 0,45 μL etiketli boyutlandırma standardı ile karıştırın. Girdap reaktif iyi karıştırmak ve daha sonra okuyucu plaka kuyusuna 35,5 μL dağıtmak.
- Kuyuya 1 μL PCR ürün ekleyin. Buna ek olarak, su buharlaşmasını önlemek için mineral yağ bir damla ekleyin.
- Ayırma plakasını ayırma tamponu ekleyerek hazırlayın.
- Gen analizörüyle birlikte verilen ticari yazılımı kullanın (bkz. Malzemeler Tablosu). Yeni bir örnek plaka oluşturun ve örnek adları plakadaki tüm kuyulara kaydedin.
- Çözümleme yöntemini seçin. Bu durumda, numuneleri 120 s için 90 °C'de denatüre, 30 s için 2,0 kV enjekte edin ve 35 dk için 6,0 kV'da ayırın.
- İki plakayı gen analizörüne yerleştirin. Kılcal damar dizisini distile suyla doldurun.
- Patentli lineer poliakrilamid (LPA) jeli yükleyin. Son olarak, Çalıştır'ıtıklatın.
-
Jel Elektroforez
- 150 mL'lik 1x TAE (Tris-asetat-EDTA) elektroforezinde 150 mL'lik moleküler biyoloji sınıfı agarose 1.5 g ekleyerek %1 agarose jel (40 mM Tris, 20 mM asetik asit ve pH 8.0'da 1 mM EDTA) hazırlayın. 2-3 dk mikrodalga ısıtma ile agarose çözünür.
- DNA'yı görselleştirmek için, bir nükleik asit lekesi 4 μL ekleyin (Bkz. Malzemeler Tablosu) ve hafifçe karıştırın.
- Bir jel tepsiiçine, merdivenler, kontroller ve numuneler için yeterli kuyular ile bir jel tarak ekleyin. Sonra, jel tepsinin ortasına yavaş yavaş karışımı dökün ve kabarcıklar kaçının.
- Jel tamamen katılaşmış kadar oda sıcaklığında 30-45 dakika boyunca jel soğumasını bekleyin. Elektroforez odasında jel tanıtın, jel tarak çıkarın ve jel kapsayacak kadar 1x TAE tampon ile oda doldurun.
NOT: Jelin yerleşimini kontrol edin. Negatif yüklü DNA katota doğru göç ettiği için kuyular negatif kutbuna yakın yerleştirilmelidir. - PCR ürünlerine 5 μL yükleme tamponu (%0,1 (v/v) bromofenol mavisi ekleyin ve iyice karıştırın.
- Bantların boyutunu tahmin etmek için 5 μL DNA moleküler ağırlık merdiveni yükleyin (bkz. Malzeme Tablosu).
- Numuneleri jelin ek kuyularına yükleyin.
- Tüm numuneler ve DNA moleküler ağırlık merdiveni yüklendikten sonra jeli 90 V'de 1-1,5 saat çalıştırın, ta ki mavi boya hattı jelin uzunluğunun yaklaşık %75'ine kadar.
- Nükleik asitler için bir transilluminator bantları görselleştirin.
-
Kılcal Elektroforez
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Kayısıda tozlaşma çalışmaları, antezten bir gün önce geç balon evresinde çiçek kullanılmasını gerektirir (Şekil 1A). Çiçek yapıları neredeyse olgun olduğundan bu aşama hem polen hem de pistil koleksiyon için en uygun evre olarak kabul edilir, ancak anher ayrılması henüz gerçekleşmemiştir. Kapalı yaprakları dış polen taşıyan böceklerin gelişini engellediği için, bu sadece aynı çiçekten değil, diğer çiçeklerden de istenmeyen polenlerin karışmasını önler. Polen taneleri, daha önce oda sıcaklığında 24 saat veya hafif ekstra ısı ile bir kağıt parçasına yerleştirilen dehisced anthers ince bir örgü(Şekil 1B)ile kolayca elek. Aynı şekilde, pistils cımbız veya tırnak yardımı ile yaprakları, sepals ve stamens kaldırılmasından sonra balon aşamasında çiçekler elde edilir(Şekil 1C). Pistils kendi kendine olabilir ve ince bir fırça ile çapraz tozlaşma (Şekil 1D).
Kayısı hermafrodit çiçekleri beş koyu kırmızı sepals, beş beyaz yaprakları(Şekil 1A),tek bir pistil (Şekil 2A) ve 25-30 stamens var. Pistil üç ana yapıya sahiptir: stigma, stil ve yumurtalık. Yumurtalıkta iki ovules vardır ve bunlardan en az birinin döllenmesi meyve ayarı için gereklidir. Tozlaşma sırasında, böcekler, özellikle arılar, polen tanelerini tozlaşmadan sonra 24 saat içinde çimlendikleri stigmayaB(Şekil1A)aktarırlar. Bir polen tüpü 3-4 gün sonra yumurtalık ulaşmak için pistil yapılar aracılığıyla yetişen ve yaklaşık 7 gün sonra iki ovules birini döllemek her çimlenmiş polen tahıl, üretilir. Polen tanelerinin S alelinin iki pistilden biri ile aynı olduğu kendi kendine uyumsuz çeşitlerinde polen tüpü üst tarzda büyümeyi durdurarak döllenmeyi önler (Şekil 2C). Ancak, uyumlu bir çeşitten polen tüpleri, farklı bir S alel ile, tarzı ile büyüyebilir (Şekil 2D), yumurtalık ulaşmak (Şekil 2E) ve iki ovules birini döllemek.
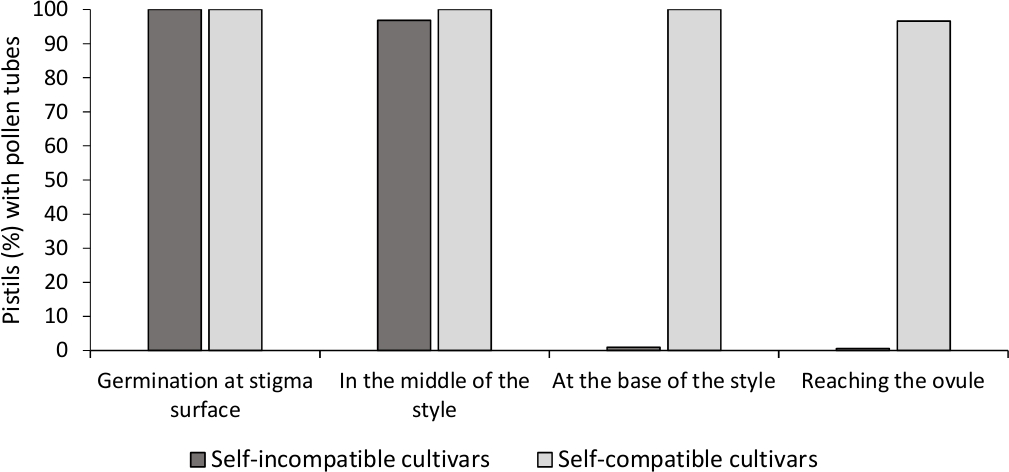
İn vitro polen çimlenme analizi, çoğu polen tüpleri kültür orta 24 saat sonra polen tahıl uzunluğu daha uzun olduğundan, burada analiz tüm çeşitleri iyi polen canlılığı gösterdi. Tüm polenlerden gelen rilerle'lerde, yeterli tozlaşmayı gösteren, çimlenmiş polen taneleri stigma yüzeyinde(Şekil 2B)gözlenmiştir (Şekil 3).
Floresan mikroskopide her bir çeşitiçin öz-(in)uyumluluğunu belirlemek için, laboratuvar kontrollü koşullarda yapılan kendi kendine ve çapraz polenlerde polen tüpü davranışı gözlendi. Polen tüpü büyümesi incelenen tüm pistillerde stil boyunca kaydedildi. Polen tüpü büyümesi çoğu kendi kendine tozlaşmış pistillerde(Şekil 2C, Şekil 3) ve en az bir polen tüpü incelenen pistillerin çoğunda stilin tabanına ulaştığında kendi kendine uyumlu olarak değerlendirildiğinde, çeşitleri kendi kendine uyumsuz olarak kabul edilmiştir (Şekil 2E, Şekil 3).
S-locus'un PCR analizi ile incelenmesi, her bir çeşidin S-genotipinin karakterizasyonuna olanak sağladı. İlk olarak, S-alelleri ilk S-RNase intronunun src-f/src-r(Tablo 2)kullanılarak amplifikasyonu ile tanımlanmıştır. Güçlendirilmiş parçaların büyüklüğü kapiller elektroforez(Şekil 4A)ile analiz edilmiş ve ilgili uyumsuzluk grubunda (I.G.) analiz edilen genotipleri sınıflandırmak için kullanılmıştır. (Tablo 3).
S1 ve S7 veya S6 ve S9gibi alellerin bazı çiftleri, ilk intron için benzer parça boyutları gösterdi. Böylece, bu alellerin farklılaşması, RNase'nin ikinci intron bölgesinin pru-C2/PruC4R, SHLM1/SHLM2 ve SHLM3/SHLM4(Tablo 2)ile yükseltilmesi yle yapılmıştır. PruC2/PruC4R astar kombinasyonu S6 ve S9'uayırt etmek için kullanılmıştır. S6için 1300 bp'lik bir parça yükseltilirken, S9 alel için yaklaşık 700 bp'lik bir parça gözlendi ( Şekil4B, Tablo 3). Özel astarlar SHLM1/SHLM2 ve SHLM3/SHLM4, S1 alelinde yaklaşık 650 bp ve S7 alelinde 413 bp'lik bir parçayı güçlendirdi ( Şekil4C, Tablo 3).
SFB geninin V2 ve HVb değişken bölgelerini yükselten AprFBC8-(F/R) astarları, her iki alel de aynı RNase dizilimini gösterdiğinden Sc ve S8 alellerini ayırt etmek için kullanıldı. S8 alel yaklaşık 150 bp pcr-parçası gösterirken, 500 bp'lik bir parça Sc aleline karşılık gelirken ( Şekil4D, Tablo 3). Tümçeşitlerin S -genotip'i belirlendikten sonra, kendi kendine uyumsuz çeşitleri, S-alelleri temel alınerek karşılık gelen uyumsuzluk gruplarına atanmıştır(Tablo 3).
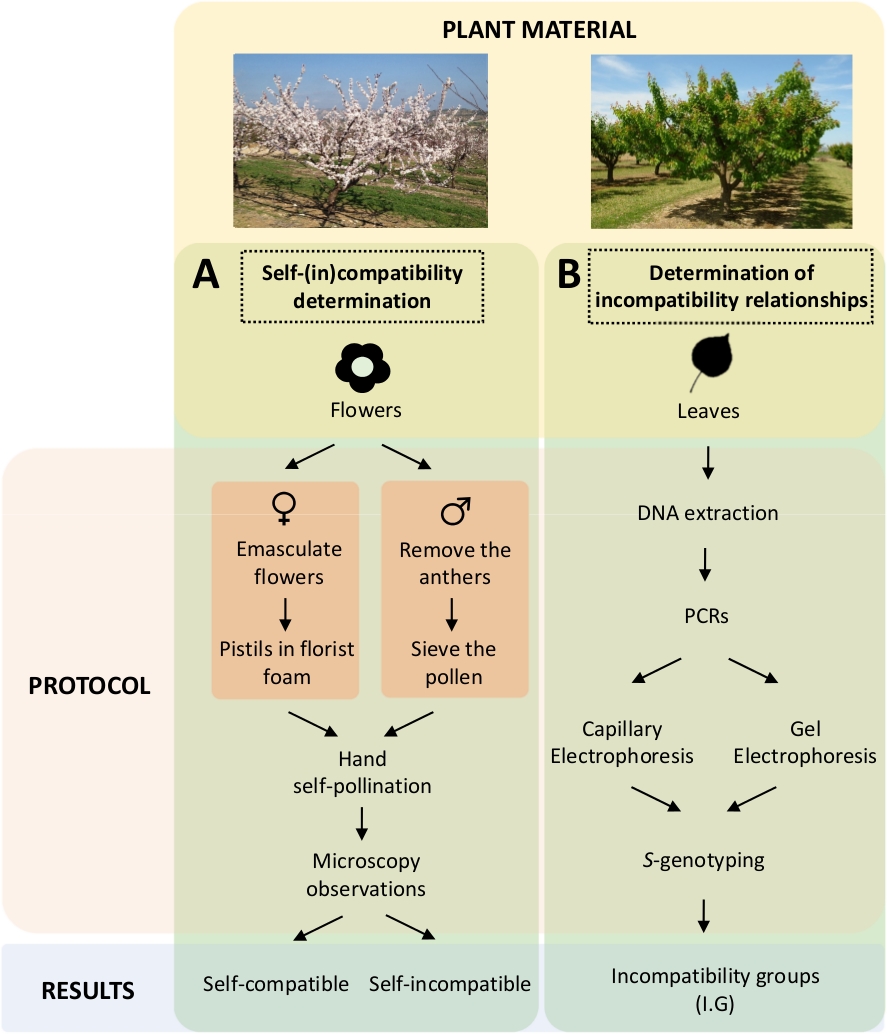
Bu yaklaşım, her bir çeşidinin laboratuvarda kontrollü öz ve çapraz polenlerle(Şekil 5A)s-genotipin genetik Sanalizle karakterizasyonuile birlikte kendi kendine uyumluluğunun belirlenmesini gerektirir (Şekil 5B). Sonuç olarak, her bir çeşidinin tozlaşma gereksinimleri ve kayısı çeşitleri arasındaki uyumsuzluk ilişkileri belirlenebilir.

Şekil 1. Kayısıda kendi kendine uyumluluk tespiti için deneysel kurulum.
(A) Balon aşamasında çiçekler (siyah oklar) alanında. (B) Polen tanelerinin ince bir örgü kullanarak elek. (C) Suda çiçekçi köpüğüüzerine yerleştirilen pistiller. (D) Bir boya fırçası yardımıyla pistillerin elle tozlaşması. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.

Şekil 2. Kayısı çiçekleri gametofitik uyumsuzluk ilişkilerinin diyagramatik gösterimi.
(A) Gametofitik Kendi Kendine Uyumsuzluk (GSI) olarak, hem uyumlu hem de uyumsuz polen taneleri stigma üzerinde çimlenir. Polen tanesi orijinal genotipin iki Salelinden birini taşır, bu durumda s1 veya S2. Polen tanelerinin Salelleri pistil'in iki Salelinden biriyle eşleşirse, bu durumda S1S3, polen tüpü büyümesi stilin üçte birinde inhibe edilir. (B) Stigma yüzeyinde polen tanelerinin çimlenmesi. (C) Polen tüpü uyumsuz bir davranış gösteren tarzında tutuklandı. (D) Polen tüpleri tarzı boyunca büyüyen. (E) Polen tüpleri tarzının tabanında uyumlu bir davranış gösteren. Ölçek çubukları, 100 μm. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.

Şekil 3. Polen çimlenme ve polen tüpü büyüme temsilcisi sonuçları kendi kendine pollinations sonra kendini uyumlu ve kendini uyumsuz çeşitleri için tarzı ile.
Stigma yüzeyinde çimlenmiş polen taneleri ile pistils yüzdesi, yarı yolda polen tüpleri ile tarzı, tarzı tabanında, ve ovule ulaşan. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.

Şekil 4. S-alellerinin tanımlanması için beş astar çifti kombinasyonu nu kullanarak PCR parça amplifikasyonu.
(A) SRc-(F/R) astarları için gen analizörü çıkışı, S-alellerine karşılık gelen ilk intron bölgesinin iki güçlendirilmiş parçasının boyutunu gösterir. (B) S6 ve S9 alellerinin tanımlanması için PruC2/PruC4R astarlarını kullanarak PCR amplifikasyonu. (C) PCR ürünleri S7 7 alel ayırt etmek için S1 alel ve SHLM3 ve SHLM4 farklılaşması için özel astar SHLM1 ve SHLM2 kullanılarak elde edin. (D) Sc ve S8 alellerinin tanımlanması için AprFBC8-(F/R) astarları ile PCR amplifikasyonu. MI: 1 kb DNA Merdiveni. MII: 100 bp DNA Merdiveni. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.

Şekil 5. Kayısı çeşitlerinde öz-ve inter-(in) uyumluluk ilişkilerini açıklığa kavuşturmak için deneysel tasarım şeması.
(A) Laboratuvarda kontrollü pollinations tarafından kendi kendine uyumluluk belirlenmesi iş akışı. (B) Moleküler yaklaşımlarla S-alel tanımlamasının iş akışı. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
| PCR Master Mix | Termocycler koşulları | ||||||
| Bileşen | Son Konsantrasyon | 15 μL reaksiyonu | Döngü Adımı | Sıcaklık | Zaman | Döngü | |
| 10x NH4 Reaksiyon Tamponu | 10x | 1,5 μL | İlk denatürasyon | 94 °C | 3 dk | 1 | |
| 50 mM MgCl2 Çözeltisi | 25mm | 1.2 μL | Denatüre | 94 °C | 1 dk | 35 | |
| 100 mM dNTP | 2,5 mM | 0,6 μL | Tavlama | 55 °C | 1 dk | ||
| Astar SRc-F | 10 μM | 0,6 μL | Uzantısı | 72 °C | 3 dk | ||
| Astar SRc-R | 10 μM | 0,6 μL | Son Uzatma | 72 °C | 5 dk | 1 | |
| 500 U Taq DNA Polimeraz | 0.5 U | 0,2 μL | 4 °C | Tutun | |||
| H2O | 8.3 μL | ||||||
| Bileşen | Son Konsantrasyon | 25 μL reaksiyonu | Döngü Adımı | Sıcaklık | Zaman | Döngü | |
| 10x PCR tampon | 10x | 2,5 μL | İlk denatürasyon | 94 °C | 2 dk | 1 | |
| 5x Q-çözeltisi | 5 x | 5 μL | Denatüre | 94 °C | 10 s | 10 | |
| 100 mM dNTP | 2,5 mM | 0,5 μL | Tavlama | 55 °C | 2 dk | ||
| Astar PruC2 | 10 μM | 0,2 μL | Uzantısı | 68 °C | 2 dk | ||
| Astar C4R | 10 μM | 0,2 μL | Denatüre | 94 °C | 10 s | 25 | |
| 250 U Taq DNA Polimeraz | 10 U | 0,13 μL | Tavlama | 58 °C | 2 dk | ||
| H2O | 15,5 μL | Uzantı* | 68 °C | 2 dk | |||
| Son Uzatma | 72 °C | 5 dk | 1 | ||||
| 4 °C | Tutun | ||||||
| * 10 s ile% 68 C uzatma adımına her döngüsü eklendi. | |||||||
| Bileşen | Son Konsantrasyon | 25 μL reaksiyonu | Döngü Adımı | Sıcaklık | Zaman | Döngü | |
| 10x PCR tampon | 10x | 2,5 μL | İlk denatürasyon | 94 °C | 2 dk | 1 | |
| 5x Q-çözeltisi | 5 x | 5 μL | Denatüre | 94 °C | 30 s | 35 | |
| 100 mM dNTP | 2,5 mM | 0,5 μL | Tavlama | 62 °C | 1,5 dk | ||
| Astar SHLM1 | 10 μM | 0,2 μL | Uzantısı | 72 °C | 2 dk | ||
| Astar SHLM2 | 10 μM | 0,2 μL | Son Uzatma | 72 °C | 5 dk | 1 | |
| 250 U Taq DNA Polimeraz | 10 U | 0,13 μL | 4 °C | Tutun | |||
| H2O | 15,5 μL | ||||||
| Bileşen | Son Konsantrasyon | 20 μL reaksiyonu | Döngü Adımı | Sıcaklık | Zaman | Döngü | |
| 5x PCR Tampon | 5 x | 4 μL | İlk denatürasyon | 98 °C | 30 s | 1 | |
| dNTP | 2,5 mM | 1,6 μL | Denatüre | 98 °C | 10 s | 35 | |
| Astar SHLM3 | 10 μM | 1 μL | Tavlama | 51 °C | 30 s | ||
| Astar SHLM4 | 10 μM | 1 μL | Uzantısı | 72 °C | 1 dk | ||
| 100 U DNA Polimeraz | 5 U | 0,2 μL | Son Uzatma | 72 °C | 5 dk | 1 | |
| H2O | 12.4 μL | 4 °C | Tutun | ||||
| Bileşen | Son Konsantrasyon | 25 μL reaksiyonu | Döngü Adımı | Sıcaklık | Zaman | Döngü | |
| 10x PCR tampon | 10x | 2,5 μL | İlk denatürasyon | 94 °C | 2 dk | 1 | |
| 100 mM dNTP | 2,5 mM | 2 μL | Denatüre | 94 °C | 30 s | 35 | |
| Astar FBC8-F | 10 μM | 1 μL | Tavlama | 55 °C | 1,5 dk | ||
| Astar FBC8-R | 10 μM | 1 μL | Uzantısı | 72 °C | 2 dk | ||
| 250 U Taq DNA Polimeraz | 10 U | 0,125 μL | Son Uzatma | 72 °C | 5 dk | 1 | |
| H2O | 17.4 μL | 4 °C | Tutun | ||||
Tablo 1. Bu protokolde kullanılan farklı astar kombinasyonları için reaksiyon ve bisiklet koşulları.
| Astar | Sıra | Başvuru |
| SRc-F | 5'-CTCGCTTTCCTGTTCTTGC-3' | 18 |
| SRc-R | 5'-GGCCATTGTTGCACCCCTTG-3' | 18 |
| Pru-C2 | 5'-CTTTGGCCAAGTAATTATTCAAACC-3' | 35 |
| Pru-C4R | 5'-GGATGTGGTACGATTGAAGCG-3' | 35 |
| SHLM1-F | 5'-GGTGGAGGTGATAAGGTAGCC-3' | 17 |
| SHLM2-R | 5'-GGCTGCATAAGGAAGCTGTAGG-3' | 17 |
| SHLM3-F | 5'-TATATCTTACTCTTTGGC-3' | 17 |
| SHLM4-R | 5'-CACTATGATAATGTGTATG-3' | 17 |
| AprFBC8-F | 5'-CATGGAAAAAGCTGACTTATGG-3' | 26 |
| AprFBC8-R | 5'-GCCTCTAATGTCATCTCTTAG-3' | 26 |
Tablo 2. Bu protokolde kullanılan astarlar, Prunus ermenistanca'da S-genotip karakterizasyonu için sıra ve referans .
| Çeşitleri | SRc-(F/R) (bp) | PruC2/PruC4R (bp) | SHLM1/SHLM2 (bp) | SHLM3/SHLM4 (bp) | AprFBC8-(F/R) (bp) | S-Genotip | Uyumsuzluk grubu (I.G) |
| Wonder Cot8 | 420, 420 | 749, 1386 | S6S9 | Vııı | |||
| Sihirli Karyola8 | 334, 420 | 749 | S2S9 | Xx | |||
| Goldstrike8 | 334, 420 | 749 | S2S9 | - | |||
| T06917 | 334, 408 | 650 | S1S2 | Ⅰ | |||
| T12017 | 334, 408 | 650 | S1S2 | - | |||
| C-6 | 334, 408 | 413 | S2S7 | ıv | |||
| Cooper Cot8 | 274, 408 | 650 | S1S3 | Xvııı | |||
| Apriqueen | 358, 358 | 500 | ScSc | - | |||
| Bergecot8 | 334, 358 | 500 | S2Sc | - | |||
| Bahar Allık8 | 274, 358 | 150 | S3S8 | Xxı |
Tablo 3. Bu protokolde kullanılan beş astar çifti ile kayısı çeşitlerinin S-genotipleme ve uyumsuzluk grubu ataması. SRc-(F/R), PruC2/PruC4R, SHLM1/ SHLM2, SHLM3/SHLM4 ve AprFBC8-(F/R) astarları kullanılarak güçlendirilmiş S-alellerin farklı polimeraz zincir reaksiyonu ürün boyutları tabloda gösterilmiştir.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Geleneksel olarak, en ticari kayısı Avrupa çeşitleri kendi kendine uyumlu36edildi. Bununla birlikte, son yıllarda üreme programlarında ebeveyn olarak Kuzey Amerika kendini uyumsuz çeşitlerin kullanımı bilinmeyen tozlaşma gereksinimleri7ile yeni kendini uyumsuz çeşitlerin artan sayıda serbest bırakılması ile sonuçlandı7 ,8,37. Bu nedenle kayısı çeşitlerinde öz ve inter-(in) uyumluluk ilişkilerinin belirlenmesi giderek önem kazanmaktadır. Bu kış soğutma azalan bu alanlarda vurgulanır, çiçeklenme zamanında yıllık varyasyonları yüksek yıl beri birçok durumda çeşitleri ve pollenizers çiçeklenme tesadüf engelliyor, özellikle yüksek soğutma gereksinimleri ile çeşitleri38. Burada açıklanan yöntem, el-tozlaşma, mikroskopi ve genetik analizleri birleştirerek her bir çeşidinin kendi kendine uyumluluğunu belirlemek ve potansiyel tozlaştırıcı çeşitlerini oluşturmak için çok yararlı olmuştur.
Tozlaşma gereksinimleri meyve bahçesi koşullarında alan kontrol deneyleri ile belirlenebilir11,39. Ancak, meteorolojik olumsuz koşullar da dahil olmak üzere dış faktörlere maruz tozlaşma yetmezliği neden olabilir10, kendi kendine uyumsuzluk hatalı tanılar neden olabilir. Burada açıklanan metodoloji, laboratuvar kontrollü koşullarda elle tozlanmış çiçeklerin mikroskopi gözlemleri ile kendi kendine uyumluluk daha doğru bir şekilde değerlendirilmesine olanak sağlayarak, çevresel etkilerden kaçınArak. Ayrıca, bu yaklaşım yılda daha fazla sayıda çeşit analizisağlar, çünkü sadece az sayıda çiçek yerine alan deneyleri40gerekli olan her çeşit için birkaç yetişkin ağaçları gereklidir.
Uyumsuzluk ilişkileri el-pollinations ve mikroskopi birleştirerek kurulabilir14. Ancak, polenler sadece ilkbaharda çiçeklenme mevsiminde kısa bir süre için yapılabilir ve toplanan çiçeklerin ömrü çok kısa olduğundan, laboratuvar ayakın yetişkin ağaçlar gereklidir. Böylece, her mevsim kontrollü el polenleri ile analiz edilebilir uyumsuzluk ilişkilerinin sayısı çok düşüktür. S-locus tarafından kodlanan genlerin karakterizasyonu S-alel genotipleme18,41için PCR tabanlı yöntemlerin geliştirilmesini sağlamıştır. Bu yaklaşım çiçek gerektirmediğinden S-alel tanımlamasını hızlandırır ve deneyler herhangi bir bitkisel doku42ile yapılabilir. Bu, genellikle genç yapraklar olan bitki materyalinin43'te toplayabildiği süreyi uzatamam. Ayrıca, yaprakları lyophilized veya dondurulmuş olabilir, böylece analiz yılın herhangi bir zamanında yapılabilir, sadece çiçekli sezon44sırasında taze çiçekler üzerinde yapılabilir pollinations aksine . Ek bir yararı yaprakları bile çiçeklenme yaş girmeden önce genç ağaçlardan toplanabilir, örneklerin toplanması kolaylaştırılması ve sonuçların erken elde45.
Bu güçlendirilmiş parça boyutları21,46hassas sonuçlar sağlar beri genetik analiz kendi kendine uyumsuzluk aleldaha iyi bir farklılaşma sağlar. Bugüne kadar, otuz üç S-alel kayısı tespit edilmiştir12,18,20,21,22,23,24, Hangi S dayalı 36 uyumsuzluk grupları kurmak için izin verdi-genotip8,9,17,25,26,27. Öte yandan, bu metodolojinin bir dezavantajı, aynı aralık boyutunda farklı alellerin hatalı bir şekilde aynı alel olarak tanımlanabildiğidir. Böylece, Sc ve S8 alelR dizisi için aynıdır ancak ScSFB geni nde 358 bp'lik bir ekleme bulunur19. RNase Aynı şekilde, alel S1 ve S S7'nin ilk intron bölgesi aynıdır ve SRc-F/SRc-R astarları kullanılarak ayırt edilemez. Buna ek olarak, s6 ve S528 veya S20 ve S55, ve S7, S13 (EF062341) ve S4617gibi çeşitli homologlar, bu alellerin bazıları kısmen sıralanmış veya PCR amplifikasyon sırasında başarısızlıklar nedeniyle bulundu ve sonuç olarak, daha fazla çalışma onları ayırt etmek için gereklidir.
PCR analizi ve S-RNase sıralaması, S-alellerinin tanımlanması ve ilgili Uyumsuzluk Grubu8,17,,26,27'deçeşitlerin tahsisi yoluyla uyumsuzluk ilişkileri kurmak için yeterlidir. Ancak bu metodoloji, belirli kayısı çeşitleri için öz-(in)uyumluluk belirlenmesini önleme sınırlaması vardır. Kendi kendine uyumluluk (SC) diğer Prunus türlerinde belirli S-alel ile ilişkili olmuştur47, badem gibi (Sf)48,49 veya tatlı kiraz (S4')50,51. Ancak, kayısı, SC 21ile ilişkili olan Sc alel, yanlışlıkla S S8olarak tanımlanabilir , kendi kendine uyumsuz alel19,22, ve olası mutasyonlar S locus bağlı olmayan, M-locus12olarak,52, SC conferring tespit edilmiştir. Son zamanlarda, M-locus SSR işaretleri kullanılarak genotipli olmuştur12. Bu nedenle, kayısı genotipleri için SC'nin genetik tanımlaması daha fazla araştırma gerektirir ve S lokusu ile bağlantılı olmayan faktörlerden kaynaklanan hataları önlemek için, bu çalışmada polen tüplerinin davranışını kendi kendine tozlaşmış çiçeklerin pistil'i aracılığıyla fenotipleme ile de belirlenir.
Burada tanımlanan metodoloji, laboratuvar koşullarında el polemasyonları ile öz-polenlerin belirlenmesini, floresan mikroskopisi altında kontrollü öz polemasyonların pistil'deki davranışlarının daha sonra gözlemlemesi Sve S-genotipinin PCR analizi ile belirlenmesi kayısı çeşitlerinin tozlaşma gereksinimlerinin belirlenmesine olanak sağlamaktadır. Bu yetiştiriciler ve yetiştiriciler için değerli bir bilgi sağlar, çünkü yeni meyve bahçeleri tasarımında uygun tozlaştırıcılar seçmek için çeşitleri arasında uyumsuzluk ilişkileri kurulması sağlar yanı sıra kayısı ıslah programlarında yeni haçlar tasarlamak için uygun veliler seçmek için.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Yazarların açıklayacak bir şeyi yok.
Acknowledgments
Bu araştırma Ministerio de Ciencia, Innovación y Universidades-European Development Fund, Avrupa Birliği (AGL2016-77267-R ve AGL2015-74071-JIN) tarafından finanse edilmiştir; Instituto Nacional de Investigación y Tecnología Agraria y Alimentaria (RFP2015-00015-00, RTA2017-00003-00); Gobierno de Aragón-Avrupa Sosyal Fonu, Avrupa Birliği (Grupo Consolidado A12_17R), Fundación Biodiversidad ve Agroseguro S.A.
Materials
| Name | Company | Catalog Number | Comments |
| Agarose D1 Low EEO | Conda | 8010.22 | |
| BIOTAQ DNA Polymerase kit | Bioline | BIO-21060 | |
| Bright field microscope | Leica Microsystems | DM2500 | |
| CEQ System Software | Beckman Coulter | ||
| DNeasy Plant Mini Kit | QIAGEN | 69106 | |
| dNTP Set, 4 x 25 µmol | Bioline | BIO-39025 | |
| GenomeLab DNA Size Standard Kit - 400 | Beckman Coulter | 608098 | |
| GenomeLab GeXP Genetic Analysis System | Beckman Coulter | ||
| GenomeLab Separation Buffer | Beckman Coulter | 608012 | |
| GenomeLab Separation Gel LPA-1 | Beckman Coulter | 391438 | |
| HyperLadder 100bp | Bioline | BIO-33029 | |
| HyperLadder 1kb | Bioline | BIO-33025 | |
| Image Analysis System | Leica Microsystems | ||
| Molecular Imager VersaDoc MP 4000 system | Bio-Rad | 170-8640 | |
| NanoDrop One Spectrophotometer | Thermo Fisher Scientific | 13-400-518 | |
| pH-Meter BASIC 20 | Crison | ||
| Phusion High-Fidelity PCR Kit | Thermo Fisher Scientific | F553S | |
| Power Pack P 25 T | Biometra | ||
| Primer Forward | Isogen Life Science | ||
| Primer Reverse | Isogen Life Science | ||
| Quantity One Software | Bio-Rad | ||
| Stereoscopic microscope | Leica Microsystems | MZ-16 | |
| Sub-Cell GT | Bio-Rad | ||
| SYBR Safe DNA Gel Stain | Thermo Fisher Scientific | S33102 | |
| T100 Thermal Cycler | Bio-Rad | 1861096 | |
| Taq DNA Polymerase | QIAGEN | 201203 | |
| Vertical Stand Autoclave | JP Selecta |
References
- Silva, N. F., Goring, D. R. Mechanisms of self-incompatibility in flowering plants. Cellular and Molecular Life Sciences. 58, 1988-2007 (2001).
- Charlesworth, D., Vekemans, X., Castric, V., Glémin, S. Plant self-incompatibility systems: A molecular evolutionary perspective. New phytologist. 168, 61-69 (2005).
- Tao, R., et al. Identification of stylar RNases associated with gametophytic self-incompatibility in almond (Prunus dulcis). Plant and Cell Physiology. 38, 304-311 (1997).
- Ushijima, K., et al. Structural and transcriptional analysis of the self-incompatibility locus of almond: Identification of a pollen-expressed F-box gene with haplotype-specific polymorphism. The Plant cell. 15, 771-781 (2003).
- Bedinger, P. A., Broz, A. K., Tovar-Mendez, A., McClure, B. Pollen-Pistil Interactions and Their Role in Mate Selection. Plant Physiology. 173, 79-90 (2017).
- Guerra, M. E., Rodrigo, J. Japanese plum pollination: A review. Scientia Horticulturae. 197, 674-686 (2015).
- Zhebentyayeva, T., Ledbetter, C., Burgos, L., Llacer, G. Apricot. Fruit Breeding. Badenes, M. L., Byrne, D. , Springer. Boston, MA. 415-458 (2012).
- Herrera, S., Lora, J., Hormaza, J. I., Herrero, M., Rodrigo, J. Optimizing Production in the New Generation of Apricot Cultivars: Self-incompatibility, S-RNase Allele Identification, and Incompatibility Group Assignment. Frontiers in Plant Science. 9, 527 (2018).
- Egea, J., Burgos, L. Detecting Cross-incompatibility of Three North American Apricot Cultivars and Establishing the First Incompatibility Group in Apricot. Journal of the American Society for Horticultural Science. 121, 1002-1005 (1996).
- Rodrigo, J., Herrero, M. Effects of pre-blossom temperatures on flower development and fruit set in apricot. Scientia Horticulturae. 92, 125-135 (2002).
- Julian, C., Herrero, M., Rodrigo, J. Flower bud differentiation and development in fruiting and non-fruiting shoots in relation to fruit set in apricot (Prunus armeniaca). Trees. 24, 833-841 (2010).
- Muñoz-Sanz, J. V., Zuriaga, E., López, I., Badenes, M. L., Romero, C. Self-(in)compatibility in apricot germplasm is controlled by two major loci, S and M. BMC Plant Biology. 17, 82 (2017).
- Burgos, L., Berenguer, T., Egea, J. Self- and Cross-compatibility among Apricot Cultivars. HortScience. 28, 148-150 (1993).
- Rodrigo, J., Herrero, M. Evaluation of pollination as the cause of erratic fruit set in apricot "Moniqui". Journal of Horticultural Science. 71, 801-805 (1996).
- Milatović, D., Nikolić, D., Krška, B. Testing of self-(in)compatibility in apricot cultivars from European breeding programmes. Horticultural Science. 40 (2), 65-71 (2013).
- Milatović, D., Nikolić, D., Fotirić-Aksić, M., Radović, A. Testing of self-(in)compatibility in apricot cultivars using fluorescence microscopy. Acta Scientiarum Polonorum, Hortorum Cultus. 12 (6), 103-113 (2013).
- Herrera, S., Rodrigo, J., Hormaza, J. I., Lora, J. Identification of Self-Incompatibility Alleles by Specific PCR Analysis and S-RNase Sequencing in Apricot. Int J Mol Sci. 19, 3612 (2018).
- Romero, C., et al. Analysis of the S-locus structure in Prunus armeniaca L. Identification of S-haplotype specific S-RNase and F-box genes. Plant Molecular Biology. 56, 145-157 (2004).
- Halász, J., Pedryc, A., Hegedus, A. Origin and dissemination of the pollen-part mutated SC haplotype which confers self-compatibility in apricot (Prunus armeniaca). New Phytologist. 176, 792-803 (2007).
- Halász, J., Hegedus, A., Hermán, R., Stefanovits-Bányai, É, Pedryc, A. New self-incompatibility alleles in apricot (Prunus armeniaca L.) revealed by stylar ribonuclease assay and S-PCR analysis. Euphytica. 145, 57-66 (2005).
- Vilanova, S., Romero, C., Llacer, G., Badenes, M. L., Burgos, L. Identification of Self-(in)compatibility Alleles in Apricot by PCR and Sequence Analysis. Journal of the American Society for Horticultural Science. 130, 893-898 (2005).
- Feng, J., et al. Detection and transcript expression of S-RNase gene associated with self-incompatibility in apricot (Prunus armeniaca L.). Molecular Biology Reports. 33, 215-221 (2006).
- Zhang, L., et al. Identification of self-incompatibility (S-) genotypes of Chinese apricot cultivars. Euphytica. 160, 241-248 (2008).
- Wu, J., et al. Identification of S-haplotype-specific S-RNase and SFB alleles in native Chinese apricot (Prunus armeniaca L). Journal of Horticultural Science and Biotechnology. 84, 645-652 (2009).
- Szabó, Z., Nyéki, J. Blossoming, fructification and combination of apricot varieties. Acta Horticulturae. 293, 295-302 (1991).
- Halász, J., Pedryc, A., Ercisli, S., Yilmaz, K. U., Hegedűs, A. S-genotyping supports the genetic relationships between Turkish and Hungarian apricot germplasm. Journal of the American Society for Horticultural Science. 135, 410-417 (2010).
- Lachkar, A., et al. Identification of self-(in)compatibility S-alleles and new cross-incompatibility groups in Tunisian apricot (Prunus armeniaca L.) cultivars. The Journal of Horticultural Science and Biotechnology. 88, 497-501 (2013).
- Pérez-Pastor, A., Ruiz-Sánchez, M. C., Domingo, R., Torrecillas, A. Growth and phenological stages of Búlida apricot trees in South-East. Agronomie. 24, 93-100 (2004).
- Williams, J. H., Friedman, W. E., Arnold, M. L. Developmental selection within the angiosperm style: using gamete DNA to visualize interspecific pollen competition. Proceedings of the National Academy of Sciences of the United States of America. 96, 9201-9206 (1999).
- Julian, C., Herrero, M., Rodrigo, J. Anther meiosis time is related to winter cold temperatures in apricot (Prunus armeniaca L.). Environmental and Experimental Botany. 100, 20-25 (2014).
- Guerra, M. E., López-Corrales, M., Wünsch, A., Rodrigo, J. Lack of Fruit Set Caused by Ovule Degeneration in Japanese Plum. Journal of the American Society for Horticultural Science. 136 (6), 375-381 (2011).
- Guerra, M. E., Wünsch, A., López-Corrales, M., Rodrigo, J. Flower Emasculation as the Cause for Lack of Fruit Set in Japanese Plum Crosses. Journal of the American Society for Horticultural Science. 135 (6), 556-562 (2010).
- Hormaza, J. I., Pinney, K., Polito, V. S. Correlation in the tolerance to ozone between sporophytes and male gametophytes of several fruit and nut tree species (Rosaceae). Sexual Plant Reproduction. 9, 44-48 (1996).
- Alcaraz, M. L., Hormaza, J. I., Rodrigo, J. Pistil Starch Reserves at Anthesis Correlate with Final Flower Fate in Avocado (Persea americana). PLOS ONE. 8 (10), 78467 (2013).
- Tao, R., et al. Molecular typing of S-alleles through Identification, Characterization and cDNA cloning for S-RNases in Sweet Cherry. Journal of the American Society for Horticultural Science. 124, 224-233 (1999).
- Burgos, L., et al. The self-compatibility trait of the main apricot cultivars and new selections from breeding programmes. Journal of Horticultural Science. 72, 147-154 (1997).
- Hormaza, J. I., Yamane, H., Rodrigo, J. Apricot. Genome Mapping and Molecular Breeding in Plants, Volume 4 Fruits and Nuts. Kole, C. , Springer. New York, NY. 171-187 (2007).
- Benmoussa, H., Ghrab, M., Ben Mimoun, M., Luedeling, E. Chilling and heat requirements for local and foreign almond (Prunus dulcis Mill.) cultivars in a warm Mediterranean location based on 30 years of phenology records. Agricultural and Forest Meteorology. 239, 34-46 (2017).
- Rodrigo, J., Herrero, M., Hormaza, J. I. Pistil traits and flower fate in apricot (Prunus armeniaca). Annals of Applied Biology. 154, 365-375 (2009).
- Williams, R. R. Techniques used in fruit-set experiments. Towards Regulated Cropping. Williams, R. R., Wilson, D. , Grower Books. London. 57-61 (1970).
- Sutherland, B. G., Robbins, T. P., Tobutt, K. R. Primers amplifying a range of Prunus S-alleles. Plant Breeding. 123, 582-584 (2004).
- Murray, M. G., Thompson, W. F. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Research. 8, 4321-4325 (1980).
- Porebski, S., Bailey, L. G., Baum, B. R. Modification of a CTAB DNA Extraction Protocol for Plants Containing High Polysaccharide and Polyphenol Components. Plant Molecular Biology Reporter. 15 (1), 8-15 (1997).
- Rogers, S. O., Bendich, A. J. Extraction of DNA from milligram amounts of fresh, herbarium and mummified plant tissues. Plant Molecular Biology. 5 (2), 69-76 (1985).
- Hormaza, J. I. Molecular characterization and similarity relationships among apricot (Prunus armeniaca L.) genotypes using simple sequence repeats. Theoretical and Applied Genetics. 104, 321-328 (2002).
- Sonneveld, T., Tobutt, K. R., Robbins, T. P. Allele-specific PCR detection of sweet cherry self-incompatibility (S) alleles S1 to S16 using consensus and allele-specific primers. Theoretical and Applied Genetics. 107, 1059-1070 (2003).
- Hegedus, A., Lénárt, J., Halász, J. Sexual incompatibility in Rosaceae fruit tree species: molecular interactions and evolutionary dynamics. Biologia Plantarum. 56 (2), 201-209 (2012).
- Fernández i Martí, A., Gradziel, T. M., Socias i Company, R. Methylation of the Sf locus in almond is associated with S-RNase loss of function. Plant Molecular Biology. 86, 681-689 (2014).
- Company, R. S. i, Kodad, O., Martí, A. F. i, Alonso, J. M. Mutations conferring self-compatibility in Prunus species: From deletions and insertions to epigenetic alterations. Scientia Horticulturae. 192, 125-131 (2015).
- Boskovic, R., Tobutt, K. R. Correlation of stylar ribonuclease zymograms with incompatibility alleles in sweet cherry. Euphytica. 90, 245-250 (1996).
- Cachi, A. M., Wünsch, A. S-genotyping of sweet cherry varieties from Spain and S-locus diversity in Europe. Euphytica. 197 (2), 229-236 (2014).
- Zuriaga, E., et al. An S-locus Independent Pollen Factor Confers Self-Compatibility in "Katy" Apricot. PLoS ONE. 8 (1), 53947 (2013).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}