Native collagen
The reaction mixture of ~0.3 mg/ml collagen monomer in 100 mM phosphate and 100 mM KCl at pH 7 left at 37 °C for 3-4 hr will yield a solution containing native type collagen fibrils with clear ~67 nm D-banding, without unbanded fibrils.
Under an optical microscope at 200× magnification, clumps of several fibrils can be normally seen on the mica substrate particularly when viewed by differential interference contrast (DIC) microscopy (Figure 2). In a typical sample, a random 100 × 100 μm2 scan by AFM will usually show at least a few fibrils that are 5-50 microns long (Figures 3a-c). Individual, separated collagen fibrils can be easily identified at this stage. With a 512 × 512 pixel2 image scan, zooming into a 10 × 10 μm2 scan size shows that all the fibrils are banded (Figures 3d-f). In order to accurately measure the banding periodicity, it is best to zoom into a 2 × 2 μm2 scan size (Figures 3g-i).
Banding periodicity can be determined by simply measuring and averaging the longitudinal distance between several peaks or valleys. Alternatively, if the AFM software allows, a one dimensional Fourier transform along a longitudinal cross-section can also be used. Figure 4 shows an AFM height image of a fibril and a corresponding longitudinal cross-section.
FLS collagen
The reaction mixture of 1 mg/ml μ1-acid glycoprotein and ~1 mg/ml collagen monomer in water left at room temperature for 30 min will yield a solution of FLS collagen fibrils.
Clumps of fibrils, similar to what is observed in the case of native collagen, can be easily seen on the mica substrate under an optical microscope at 200× magnification. In a typical sample, a random 100 × 100 μm2 scan by AFM will usually show at least a few fibrils. With a 512 × 512 pixel2 image scan size, the banding periodicity can be measured by zooming into a 10 × 10 μm2 scan (Figure 5a) or more accurately with a 2 × 2 μm2 scan (Figure 5b). Figure 5c shows a longitudinal cross-section of a FLS fibril.
SLS collagen
The reaction mixture of 2 mg/ml ATP and ~0.5 mg/ml of collagen monomer in 100 mM glycine-HCl buffer at pH 3.3 left at room temperature for 2 hr will yield a solution of SLS collagen.
Typically, SLS crystallites will not be visible under an optical microscope. An AFM is required to confirm the presence of SLS crystallites. In a typical sample, a random 100 × 100 μm2 scan by AFM will usually show many dots. Zooming into a 10 × 10 μm2 scan will usually show several SLS crystallites (Figure 6a). And a 2 × 2 μm2 scan (Figure 6b) shows the finer structure of an SLS crystallite.
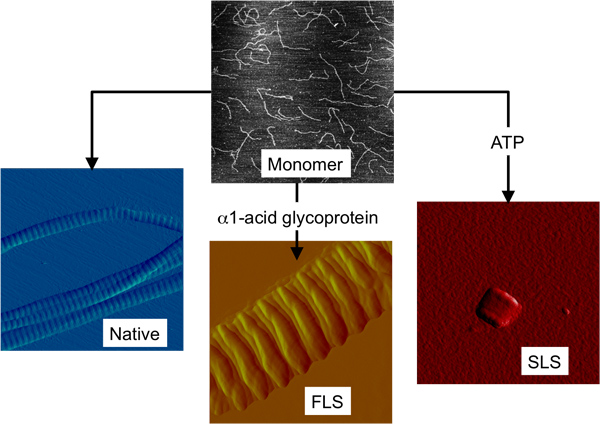
Figure 1. The three structurally different forms of collagen fibrils that can be formed in vitro from collagen monomer. AFM images depicting collagen monomer and the three higher order collagen structures all on the same size scale (2 × 2 μm2).
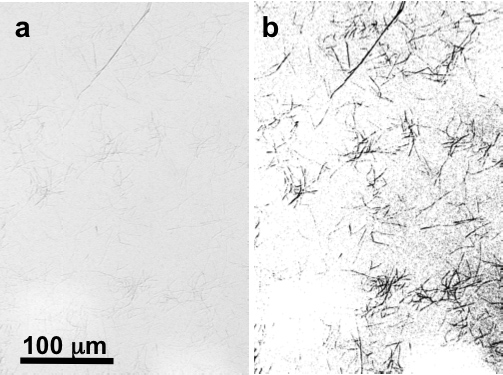
Figure 2. Digital image at ~200× magnification by optical DIC microscopy of native collagen fibrils on mica (a) untouched and (b) thresholded and sharpened to highlight the fibrils.

Figure 3. AFM images of native collagen fibrils on mica. Intermittent contact mode AFM amplitude images are shown.
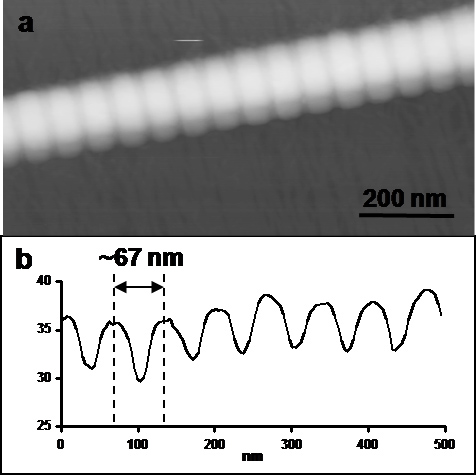
Figure 4. (a) 0.5 × 1 μm2 intermittent contact mode AFM height image (vertical scale = 100 nm) and (b) longitudinal cross-section of a native collagen fibril.

Figure 5. AFM images of FLS collagen fibrils on mica. (a) 10 × 10 μm2 contact mode AFM deflection image, (b) 2 × 2 μm2 contact mode AFM deflection image and (c) longitudinal cross-section of a FLS collagen fibril.

Figure 6. AFM images of SLS collagen on mica. Intermittent contact mode AFM amplitude images are shown.
