

Summary
We beschrijven de collectie van onbevruchte
Abstract
Veel organismen lokaliseren mRNA om specifieke subcellulaire bestemmingen aan ruimte en tijd controle genexpressie. Recente studies hebben aangetoond dat de meerderheid van de transcriptoom is gelokaliseerd op een niet-willekeurige positie in cellen en embryo's. Een benadering om gelokaliseerde mRNA's te identificeren is om biochemisch zuiveren een cellulaire structuur van de rente en alle bijbehorende transcripties identificeren. Met behulp van recent ontwikkelde high-throughput sequencing technieken is het nu eenvoudig om alle RNA's in verband met een subcellulaire structuur te identificeren. Om transcript identificatie te vergemakkelijken is het noodzakelijk om te werken met een organisme met een volledig gesequenced genoom. Een aantrekkelijk systeem voor de biochemische zuivering van subcellulaire structuren zijn ei uittreksels uit de kikker Xenopus laevis. Echter, X. laevis heeft momenteel geen een volledig gesequenced genoom, waarvan transcript identificatie belemmert. In dit artikel beschrijven we een werkwijzevoor ei uittreksels uit een verwante kikker, X. tropicalis, dat een volledig gesequenced genoom. Wij bieden informatie voor microtubuli polymerisatie, zuivering en transcript isolement. Hoewel dit artikel beschrijft een specifieke werkwijze voor identificatie van microtubule-geassocieerde transcripten, geloven wij dat het gemakkelijk kan worden toegepast op andere subcellulaire structuren en een krachtige techniek voor de identificatie van gelokaliseerde RNA's.
Introduction
Ruimtelijke en temporele regulatie van genexpressie is van belang voor alle cellen, en is vooral belangrijk voor de controle van de vroege embryonale kletteren 1. Ruimtelijke regulatie van genexpressie wordt bereikt door de actieve lokalisatie van mRNA naar specifieke bestemmingen binnen cellen of embryo. In vele zeer grote celtypen (bijv. oöcyten, embryo's en neuronen) mRNA lokalisatie wordt gebruikt voor eiwitexpressie de plaats van werking van het gecodeerde eiwit te beperken. Aangezien een gelokaliseerde mRNA vele rondes van eiwitproductie kan katalyseren is efficiënter om een mRNA lokaliseren op afzonderlijke eiwitmoleculen lokaliseren. Gelokaliseerde mRNA's gewoonlijk translatie onderdrukt totdat ze hun bestemming, dat dient om de lokalisatie van het gecodeerde eiwit 2 verder te beperken bereiken. Naast de vele goed gedocumenteerde gevallen van RNA lokalisatie embryonale patroonvorming beheersen, hebben verschillende studies gedocumenteerd mRNAs die gelocaliseerdop de plaats van werking van het gecodeerde eiwit. Prominente voorbeelden zijn lokalisatie van de β-actine en Arp2 3/3 4 mRNAs de voorrand beweeglijke fibroblasten en lokalisatie van de mRNAs voor veel mitotische regelgevers meiose en mitose spindels 5-7.
Veel van de klassieke voorbeelden van gelokaliseerde mRNA's werden geïdentificeerd door middel van genetische screens voor maternale effect mutaties en werden later bepaald om gelokaliseerde RNA's coderen. Echter, recente genoom-brede studies begonnen om breder inzicht te geven in de omvang van gelokaliseerde RNA's. Een recent in situ hybridisatie gebruik in Drosophila embryo's aangetoond dat ~ 70% van alle mRNAs een specifieke lokalisatie, waaronder vele nieuwe bestemmingen 8. Zuivering van pseudopodia uit muizen fibroblasten die een diverse groep van gelokaliseerde mRNA 9. Werken vanuit onze groep met biochemische zuivering van microtubuli uit meiose Xenopus ei extracten die honderden mRNAs die copurify de as 5,7. Ons werk toonde aan dat de meerderheid van microtubule-gelokaliseerde mRNA's coderen voor eiwitten die functioneren in de controle van de mitose, hetgeen het idee ondersteunt dat mRNA's gelokaliseerd op de plaats van werking van het gecodeerde eiwit. Bovendien is de mogelijkheid om mRNA invloeden te onderscheiden in een subcellulaire fractie die door biochemische zuivering benadrukt de kracht van deze benadering voor de identificatie van gelokaliseerde mRNA.
De meeste gelokaliseerde RNA's gebruiken actief transport op het cytoskelet, hetzij actine of microtubuli, om het vervoer te komen naar hun eindbestemming 10. Om een beter begrip van de omvang en de aard van RNAs die zijn gelokaliseerd aan specifieke bestemmingen met een biochemische benadering moet een hebben in vitro systeem dat cytoskelet processen recapituleren krijgen. Een van de belangrijkste systemen voor het bestuderen van het cytoskelet biologie is ei extracten geproduceerduit onbevruchte eieren van de kikker Xenopus laevis. X. laevis ei extracten zijn gebruikt voor decennia om een breed scala van cytoskelet processen te bestuderen en hebben veel bijgedragen aan ons begrip van de mechanismen en moleculen die cytoskelet assemblage en dynamiek 11 beheersen. Voorts X. laevis ei extracten vatbaar zijn voor grootschalige zuiveringen van microtubuli geassocieerde eiwitten en 12,13 en zijn goed ontworpen werkwijzen voor de productie van verschillende soorten ei extracten 14-16. Voor genomische studies er verschillende nadelen aan het gebruik van X. laevis als modelsysteem.
Al decennia Xenopus laevis kikkers hebben een krachtig systeem voor de studie van de ontwikkelingsbiologie en celbiologie, gezien het grote eicel omvang en robuuste externe ontwikkeling 17 geweest. Bovendien is de ontwikkeling van ei extract systemen vele cellulaire processe kunnen recapitulerens in een reageerbuis heeft deze kikker heeft een krachtig experimenteel model. Echter, Xenopus laevis gehinderd door het ontbreken van een genomische sequentie, die is vertraagd door de aard van de allotetraploid genome.In contrast, een nauw verwante soorten, Xenopus tropicalis, een diploïd genoom die werd gesequenced 2010 18. Terwijl X. tropicalis is niet zo experimenteel handelbaar als X. laevis 17 van de beschikbaarheid van een gesequenced genoom maakt het een aantrekkelijk model systeem om het gehele genoom analyses uit te voeren.
In dit rapport beschrijven we een methode om meiose II-, cytostatica factor-gearresteerd extracten (CSF) te maken van X. tropicalis 19. Daarna beschrijven we een eenvoudige methode om microtubuli en bijbehorende RNA te zuiveren van dit extract. Het RNA kan vervolgens worden omgezet in bibliotheken vatbaar voor sequencing met behulp van recent ontwikkelde high throughput sequencing technologieën. Zodra de bibliothekenzijn sequentie kunnen worden aangepast aan het genoom van de kikker specifieke mRNAs die verrijkt in de microtubule monster vergeleken droge stof identificeren. Dit zorgt voor een krachtige methode om microtubuli-gerichte mRNA lokalisatie te detecteren op een genoom-brede schaal. Naast de mogelijkheid om gelokaliseerde mRNA te detecteren, het gebruik van high-throughput sequencing en gesequenced genoom bieden de mogelijkheid van het ontdekken van nieuwe transcripten die momenteel aanwezig zijn in openbare databank annotaties.
Protocol
1. Generatie X. tropicalis Eieren
Alle Xenopus tropicalis kikkers zijn gerangschikt van NASCO. Onze kikkers zijn gevestigd in een Aquatic Habitats recirculerend watersysteem bewaard bij 27 ° C. Er zijn vele mogelijkheden voor water-systemen voor de verzorging van X. tropicalis. Enkele goede algemene informatie over deze kikker soorten zijn te vinden op de websites van de Harland en Grainger labs ( http://tropicalis.berkeley.edu/home/ , http://www.faculty.virginia.edu/xtropicalis / ). Onze kikkers worden onderhouden in tankwater bestaande uit (0,4 g ciclid Lake Zouten, 0,6 g zeezout, 0.625 g NaHCO3 per liter water, pH 7,0) 20. Dit recept leidt tot een geleidbaarheid van ~ 1800 uS, wat een hoog zoutgehalte voor X. tropicalis. Wij hebben echter gevonden dat onze kikkers gedijen in this milieu en eicel kwaliteit wordt verbeterd. Alternatieve tankwater recepten hierboven te vinden op de lijst voor de algemene middelen X. tropicalis zorg.
- Kikkers worden geïnjecteerd met humaan choriongonadotrofine (hCG) op drie achtereenvolgende dagen op eieren te stimuleren leggen: Ten eerste, de voorbereiding van twee concentraties van hCG-oplossing. Resuspendeer 10.000 E hCG gelyofiliseerd poeder in 10 ml steriel, gedeïoniseerd H2O voor een uiteindelijke concentratie van 1000 U / ml. Vervolgens Verdun 1 ml van 1000 U / ml hCG oplossing in 9 ml H2O een eindconcentratie van 100 U / ml. Bewaar beide oplossingen bij 4 ° C.
- Op dag 1, bereiden 4-6 kikkers voor eileg door het injecteren met hCG tussen 2:00-03:00. Injecteer elke kikker in de dorsale lymfe weg nabij de cloaca met 0,2 ml 100 E / ml hCG-oplossing. Het hebben van de kikkers snel tijdens de daaropvolgende twee injecties zal de hoeveelheid kikker afval aanwezig te minimaliseren tijdens de eileg, maar is optioneel.
- Op dag 2, injecteren dezelfde kikkersmet 0,2 ml 100 E / ml hCG-oplossing tussen 2:00-03:00.
- Op dag 3, injecteren dezelfde kikkers met 0,2 ml 1000 E / ml hCG-oplossing, tussen 7:00-10:00. Opgezet kikkers om eieren te leggen: vul een 6-liter plastic emmer met verse tankwater, kikkers en plaats toe te voegen in het donker bij 25 ° C. Na deze injectie zal eileg begint na 4 uur en zal door 7 uur compleet zijn. Kikkers moeten eieren in een omgeving die wordt gehandhaafd op een minimum van 25 ° C. lay
- Maak extraheren oplossingen en hebben apparatuur klaar direct voor het verzamelen van eieren.
20X MMR: 100 mM HEPES, pH 7,8, 2 mM EDTA pH 7,8, 2 M NaCl, 40 mM KCl, 20 mM MgCl2, 40 mM CaCl2. Autoclaaf en bewaar bij kamertemperatuur. Bereid 1 L van 1X MMR net voor de voorbereiding extraheren.
10X XB: 100 mM HEPES, pH7.7, 10 mM MgCl2, 1 mM CaCl2, 1 M KCl, 500 mM sucrose. Autoclaaf en bewaar bij 4 ° C. Bereid 1 L van 1X XB just voorafgaand aan de voorbereiding extraheren. Dejelly oplossing: Bereid 250 ml 3% cysteïne-oplossing in gedeïoniseerd H2O en pH op 7,8-8,0 met 10 N NaOH. Net voor het opstellen halen.
CSF-XB: neem 200 ml 1X XB en voeg 2 ml 0,5 M EGTA pH 7,7 en 200 ui 1 M MgCl2. Net voor het opstellen halen.
CSF-XB +: neem 50 ml van CSF-XB en voeg 50 ul van LPC (10 mg / ml elk bestand van Leupeptin, Pepstatine en chymostatine in DMSO). Voeg 50 ul cytochalasine D (10 mg / ml in DMSO). Net voor het opstellen halen.
Bereid een 0,2% gelatine-oplossing in gedeïoniseerd H2O, magnetron op te lossen en filteren steriliseren. Bewaar bij kamertemperatuur.
Reserve 2 Beckman 2 x ½ inch ultracentrifugebuizen.
Bereid twee 15 ml glazen ronde bodem centrifuge buizen met 0,5 ml H 2 O in elke aan het opvangen van deultracentrifuge buis.
Maak brand-gepolijst glas Pasteur pipetten. Snap het einde af van 5 ¾ inch glazen pipetten tot een brede opening bloot te leggen, en bloot aan vuur aan het nieuwe blootgestelde pipettip glad.
- Bereid een bekerglas 500 ml voor opslag van eieren door zwenken een 0,2% gelatine oplossing rond het bekleden van de wanden van het bekerglas. Gooi gelatine oplossing van bekerglas na gebruik.
- Verzamel eieren uit het plastic emmer gebruikt voor het leggen van 6-7 uur na de derde injectie op dag 3. Indien gewenst, knijp elke kikker een keer om de resterende eieren krijgen. Was de eieren eenmaal met verse tankwater en transfer naar de 500 ml bekerglas bekleed met 0,2% gelatine-oplossing.
2. Bereiding van het extract uit X. tropicalis Eieren
Alle stappen extract preparaat kan worden uitgevoerd bij kamertemperatuur, ongeveer 25 ° C. Gedurende het wassen is het belangrijk om het ei te houdens ondergedompeld onder vloeistof, zodat ze nat blijven. Blootstelling aan lucht kan ertoe leiden dat de eieren aan de celcyclus te ontsnappen of te lyseren.
- Decanteren zoveel tankwater mogelijk terwijl het reserveren van voldoende vloeistof om de eitjes nat te houden. Tilt de beker die eieren aan de zijkant en voeg langzaam ~ 300 ml 1X MMR aan de wand van de beker wordt zo dat fysieke beweging van de eieren geminimaliseerd. Laat eieren te regelen, dan giet af supernatant bevattende puin. X. tropicalis eieren zijn vezelig bij deze stap, dus het verwijderen van geactiveerde eieren gebeurt na dejellying. Herhaal dit voor een totaal van drie 1X MMR wasbeurten.
- Dejelly de eieren. Giet af zoveel MMR mogelijk en voeg de helft van de dejelly oplossing. Werveling continu gedurende ongeveer 5 minuten. Oplossen gelei jassen zal in het supernatant zichtbaar zijn na een paar minuten. Giet af en voeg de resterende dejelly oplossing. Blijven swirl continu tot de eieren inpakken zeer strak en al oriënteren met hun vegetatieve pool (de paalmet wit pigment) naar de bodem van de schaal. Snel decanteer zoveel dejelly oplossing mogelijk. Nadat de eieren dejellied ze zeer gevoelig voor mechanische manipulaties.
- Voeg XB zorgvuldig om de eieren. In de eerste XB wasbeurt, verwijder eieren die CSF arrestatie zijn ontsnapt door het verwijderen van gelyseerde, gezwollen, wit, en pseudocleavage eieren. Geactiveerde X. tropicalis eieren hebben de neiging om zich te vestigen in het midden boven, dus gebruik een plastic overdracht pipet om deze uit te trekken. Verwijder ook stukken huid en kikker afval. Wassen eieren in totaal driemaal met -300 ml 1X XB oplossing voorzichtig schudden eieren tussen wast en waardoor ze op de bodem van het bekerglas. Als voorheen, giet zoveel van elke wasbeurt oplossing mogelijk terwijl eieren nat.
- Wassen eieren tweemaal met CSF-XB en decanteren.
- Voeg CSF-XB + naar eieren. Met behulp van een gelatine-behandelde brand-gepolijste Pasteur pipet, overdracht eieren naar Ultra-centrifuge buizen met CSF-XB +, zorg ervoor dat u de e blootggs aan de lucht. Plaats in de 15 ml glazen centrifuge buizen met het water kussen.
- Spin eieren in een klinische centrifuge bij 200 xg gedurende 1 min, de snelheid wanneer 800 xg en centrifugeren gedurende 30 sec.
- Gebruik een aspirator om zo veel buffer te verwijderen mogelijk van eieren. Ze moeten bijna droog op de top. Snel eieren naar een Sorvall RC-6 centrifuge met een rotor HB-6 (of equivalent) en spin 17.000 xg gedurende 15 min bij 20 ° C.
- Verwijder de gele cytoplasmatische laag tussen het pigment en het lipide lagen met behulp van een 18 gauge naald bevestigd aan een 1 ml spuit. Prik de zijkant van de buis en trek de spuit langzaam naar het cytoplasma-extract laag te verkrijgen. Vermijd pigment granules zoveel mogelijk.
- Overdragen cytoplasma naar nieuwe ultracentrifugering buis. Het is normaal dat het extract te verschijnen licht bewolkt bij deze stap. Plaats in de 15 ml glazen centrifugebuis met water kussen. Nogmaals spelen 17.000 xg gedurende 10 min bij 20 ° C. Herhaal extraction met 18 gauge naald.
- Overdragen cytoplasma naar een ml microfugebuis 1.5. Schatten het extract volume en verdun Cytochalasine D en LPC 1:1000 in het extract. Goed mengen met 1 ml pipet, en neer pipetteren vaak zonder introductie van luchtbellen. Een typisch rendement van een gezonde kikker kolonie is ongeveer 300-500 ul extract / kikker. Om maximale activiteit te behouden, is het noodzakelijk om het extract te slaan en uit te voeren experimentele manipulaties bij kamertemperatuur (20-25 ° C).
3. Zuivering Taxol-gestabiliseerde Microtubules van X. tropicalis Extract
- Voeg Taxol een 100-200 pi aliquot van het extract in een eindconcentratie van 10 uM en incuberen bij kamertemperatuur gedurende 30 minuten. Voor controlereacties, behandelen een gelijkwaardige hoeveelheid extract met de microtubuli-destablilzing drug Nocodazole (10 uM). Reserve 100 ui onbehandeld extract voor analyse.
- Verdun het geneesmiddel behandeldeextraheer met 10 volumes BRB-80 (80 mM PIPES pH 6,8, 1 mM MgCl2, 1 mM EGTA) + 30% glycerol. Monteer 14 ml rondbodem polypropyleen buizen met 10 ml van BRB-80 + 60% glycerol kussen. Met een brede boring pipetpunt, de laag geneesmiddel behandelde extract reactie zachtjes bovenop de BRB-80 + 60% glycerol kussen. Centrifugeer 10 minuten bij 17.000 xg bij 20 ° C in een Sorvall RC-6 centrifuge uitgerust met een HB-6 rotor (of equivalent) en buisadapter.
- Zuig het supernatant die unsedimented extract materiaal, en was de interface tweemaal met gedemineraliseerd H 2 O. Zuig het resterende kussen volume langzaam, het verzorgen van de gel-achtige pellet met microtubuli, microtubuli-geassocieerde eiwitten, en microtubuli geassocieerde RNA's in de Taxol-behandelde monster niet te storen. De Nocodazole behandelde monster niet zichtbare materiaal bevatten. Resuspendeer de pellet in 1 ml TRIzol en ga verder met de instructies van de fabrikant voor het isoleren van RNA. Untreated extract (tot 100 pi) direct worden geresuspendeerd in 1 ml TRIzol.
- Er zijn nu commercieel verkrijgbare kits voor het bereiden transcriptome bibliotheken geschikt voor RNA-seq. Deze kunnen worden gekocht via http://www.illumina.com/ en http://www.454.com/ .
Representative Results
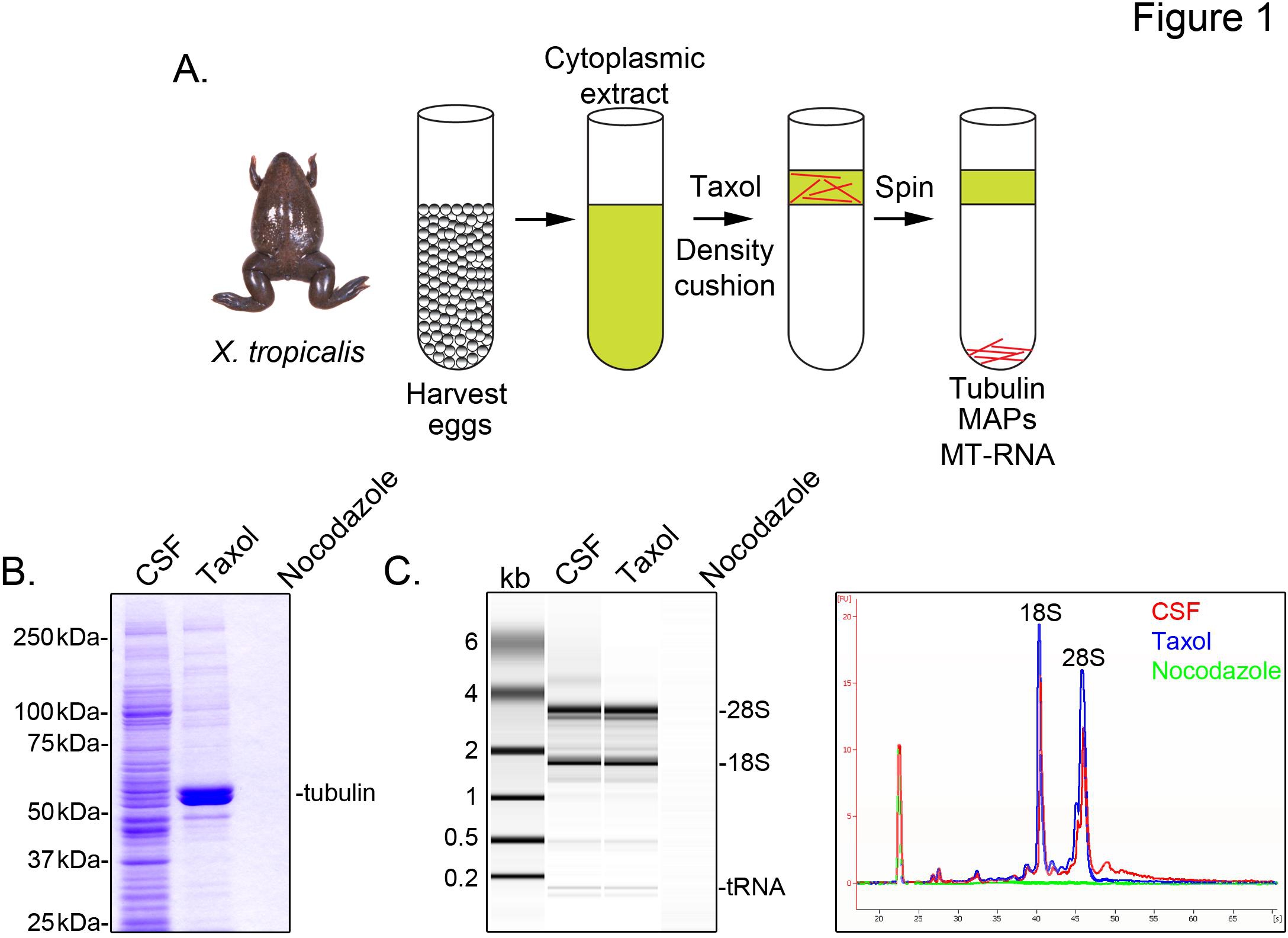
Om X. identificeren tropicalis transcripten in verband met microtubuli, bereiden we een cytosolextract uit onbevruchte eitjes gearresteerd in metafase van meiose II (CSF). Behandeling van dit extract met taxol maakt de vorming van stabiele microtubules die kunnen worden gezuiverd door sedimentatie door een glycerol kussen (figuur 1A). Coomassie gel analyse bevestigt dat α / β-tubuline sedimenten in een taxol-afhankelijke wijze, en vertegenwoordigt het belangrijkste eiwit species teruggewonnen in deze preparaten (Figuur 1B). Lagere niveaus van andere eiwitten zijn ook aanwezig in de pellet taxol, maar niet in preparaten die met de microtubuli depolymeriseren drug nocodazole, wat aangeeft dat eiwitten in de taxol fractie specifiek geassocieerd met microtubules (MAP's).
Een Agilent Bioanalyzer wordt gebruikt om algemene RNA preparaat in alle onderzoeken X. tropicalis extract fracties (figuur 1C X. zijn laevis ei extract 5,21. Een lijn spoor van de gel toont het uitsteeksel mRNA signaal aanzienlijk lager in de microtubule-taxol bevattende pellet, met name in het gebied migrerende bovenstaande 28S rRNA, wat aangeeft dat een deel van mRNA cosediment met microtubules in X. tropicalis. RNA geïsoleerd op deze manier is geschikt voor RNA-seq experimenten gebruikmaking van commercieel verkrijgbare reagentia.

Figuur 1. Zuivering van MT-RNA voor RNA-seq. (A) Zuivering regeling aan MT-RNA te isoleren. Eieren worden geoogst van vrouwelijke X. tropicalis kikkers. Na bereiding van een cytoplasmatisch extract wordt toegevoegd aan taxol microtubuluspolymerisatie induceren. Microtubuli en MT-RNA wordt gezuiverd door sedimentatie door een glycerol vulling. (B) Coomassie gel van eiwitten die met de regeling (A) beschreven. Totaal CSF extract tegenover eiwitten neergeslagen in aanwezigheid van taxol of nocodazole. (C) Bioanalyzer gel analyse van RNA geïsoleerd met behulp van de regeling (A) beschreven. RNA geïsoleerd uit CSF extract tegenover RNA neergeslagen in aanwezigheid van taxol of nocodazole. Zowel de gel projectie en de lijn sporen worden getoond. Overgenomen met toestemming van Sharp, et al.., (2011). Klik hier voor grotere afbeelding .
Discussion
In dit rapport hebben we een eenvoudige methode beschreven om CSF-gearresteerd ei uittreksels uit X. tropicalis 19 en het gebruik van dit extract aan microtubuli geassocieerde RNA's te bestuderen 7. De basisprocedure voor het produceren van CSF-gearresteerd ei uittreksels uit X. tropicalis is hetzelfde als voor X. laevis met een paar belangrijke verschillen. Een van de meest uitdagende aspecten aan het werken met X. tropicalis kikkers is het verkrijgen van voldoende hoge kwaliteit eieren om een uittreksel te maken met microtubuli nucleatie of spilstelsel activiteit vergelijkbaar met X. laevis ei haalt. Om optimale eileg voorwaarden te bereiken, terwijl het voorkomen slippen van meiose II celcyclus, het interval tussen hormoon injecties voor X. tropicalis korter zijn dan voor X. laevis en de timing van de derde hCG injectie aan het begin van eileggen is veel korter. Met X. Laevis de timing van de hCG injectie aan thij begin eileggen is zodanig dat het handig en efficiënt voor eieren nachts worden gelegd in buffer. Vanwege de korte tijd tussen hCG injectie en eieren leggen met X. tropicalis is het vaak noodzakelijk om de eieren van kikkers handmatig drukken. Een ander significant verschil tussen het maken ei extract uit de twee verschillende kikkers is dejellying stap. Met X. laevis de eieren zo groot dat het gemakkelijk is te bepalen wanneer het gelei coat door observeren hoe nauw de eieren zijn verdeeld in het bekerglas opgelost. Zoals de dejellying reactie aanvangt, de eieren beginnen te dichter pakken. Echter, X. tropicalis eieren zijn veel kleiner en het kan heel moeilijk zijn om te bepalen wanneer de gelei coat door ei pakkingdichtheid alleen al heeft opgelost. Wij hebben gevonden dat de meest betrouwbare methode om te bepalen wanneer de gelei laag opgelost is de oriëntatie van het dier (zwart) en plantaardig (wit) polen controleren. Wanneer alle plantaardige polen orient naar de bodem van het bekerglas de gelei laag is verwijderd genoeg te gaan met het extract. Tot slot, terwijl X. laevis ei-extract kan bij koele temperaturen worden opgeslagen (4-12 ° C) hebben wij geconstateerd dat het cruciaal is om X. behouden tropicalis ei extract bij kamertemperatuur (20-25 ° C) tijdens de bereiding en experimentele manipulaties biochemische activiteit behouden. Vanwege de verschillen in gebruiksgemak wij bij voorkeur X te laevis kikkers voor de productie van ei extract. Echter, voor de experimenten die vereisen of worden vergemakkelijkt door een organisme met een gesequenced genoom, X. tropicalis is een uitstekend alternatief systeem.
De methode die we in dit rapport zijn beschreven maakt gebruik van taxol als microtubuli-stabiliserende agent om microtubuli polymerisatie veroorzaken. We kozen deze methode omdat taxol is een robuuste microtubule-stabiliserende middel dat de grootschalige isolatie van gezuiverd microtubules vergemakkelijkt. Werkwijze thten we beschreven kunnen waarschijnlijk worden verbeterd door de eiwitten en RNAs verbonden microtubules alternatieve microtubuluspolymerisatie methoden vergelijken. Alternatieven kunnen onder polymerisatie met GTP-geïnduceerde polymerisatie (een klassieke techniek), 22 of met behulp van Ran-GTP als een microtubule polymerisator aan de microtubuli veroorzaakt door chromatine-driven spilstelsel 23 na te bootsen. Gebruik tenslotte gezuiverd spermakernen te induceren microtubuluspolymerisatie zou het dichtst bootsen de typen microtubuli die gekiemd tijdens de mitose (centrosome, chromatine en kinetochore gemedieerde). Nadelen van deze alternatieve microtubule kiemvorming zijn dat de kiemvormende middelen niet zo gemakkelijk verkrijgbaar als taxol en zij niet kiemen of stabiliseren microtubuli efficiënt taxol. Derhalve elk van deze methoden echter moeilijk te gebruiken voor grootschalige zuiveringen zijn. Het voordeel van het vergelijken van meerdere verschillende microtubule kiemvormendeis dat het mogelijk zou kunnen zijn om eiwitten en / of RNA's die specifiek zijn voor elke route van microtubule nucleatie identificeren.
De methode die we hier hebben beschreven maakt gebruik van cytoplasmatische extracten van amfibieën. Echter, deze benadering kan worden uitgebreid naar het gebruik van extractiesysteem van andere organismen. Mitotische extracten werden beschreven van gesynchroniseerde menselijke weefselkweek cellen 24 die trouw recapituleren veel aspecten van de assemblage van microtubuli. We hebben met succes gebruik gemaakt van deze extracten om microtubuli geassocieerde RNA's te identificeren uit HeLa-cellen 5. Soortgelijke microtubule zuiveringsschema's zijn beschreven voor verschillende organismen 25,26, hoewel het microtubule geassocieerde RNAs niet onderzocht. De hier beschreven benadering kan worden gebruikt met een organisme dat een geconcentreerde cytoplasmatisch extract kan kiemvormende microtubules kan produceren.
Tenslotte, hoewel de aanpak die we deschrijver hier bespreekt de zuivering van microtubuli en geassocieerde eiwitten en RNAs, kan deze benadering worden gegeneraliseerd naar andere subcellulaire structuren. Terwijl de meeste gelokaliseerde mRNA werden niet geïdentificeerd met biochemische werkwijzen de recente vorderingen in DNA en RNA sequencing technologie biedt een aantrekkelijke aanpak methode om gelokaliseerde RNAs identificeren. In deze benadering elke subcellulaire of sub-embryo structuur van belang zou kunnen worden geïsoleerd of gezuiverd. Dan is de bijbehorende eiwitten en RNA's kunnen worden geïdentificeerd op een genoom-wijde schaal. RNA kan dan worden vergeleken met de RNA inhoud van het totale aantal cellen of embryo verrijkte gelokaliseerde RNAs identificeren. Deze benadering kan worden gebruikt met hele eieren (dierlijk en plantaardig scheiding, vergelijkbaar met de benadering die de eerste gelokaliseerde RNA geïdentificeerd in Xenopus 27), actine geassocieerde RNAs, ER-geassocieerde RNAs, mitochondria geassocieerde RNAs, of een subcellulaire structuur die kan worden gezuiverd met bijbehorende RNA intact. Gebaseerd opons werk microtubule-geassocieerde RNA voorspellen we dat dit een uitstekende methode om nieuwe eiwitten die functioneren op een bepaalde locatie te ontdekken zijn. Bovendien zal de identificatie van de locatie en de omvang van alle gelokaliseerde RNA's inzicht geven in hoe cellen en embryo's te gebruiken mRNA lokalisatie om genexpressie te controleren.
Disclosures
Geen belangenconflicten verklaard.
Materials
| Name | Company | Catalog Number | Comments |
| REAGENTS | |||
| Xenopus tropicalis | NASCO | LM00823MX | |
| human Chorionic Gonadotropin | Sigma-Aldrich | CG10 | |
| HEPES | Sigma-Aldrich | H4034 | |
| EDTA | Sigma-Aldrich | E5134 | |
| NaCl | Sigma-Aldrich | S3014 | |
| KCl | Sigma-Aldrich | P9541 | |
| MgCl2 | Sigma-Aldrich | M8266 | |
| CaCl2 | Sigma-Aldrich | C8106 | |
| sucrose | Sigma-Aldrich | S0389 | |
| NaOH | Sigma-Aldrich | S5881 | |
| EGTA | Sigma-Aldrich | E3889 | |
| Leupeptin | Sigma-Aldrich | L9783 | |
| Pepstatin | Sigma-Aldrich | P5318 | |
| Chymostatin | Sigma-Aldrich | C7268 | |
| Cytochalasin D | Sigma-Aldrich | C8273 | |
| Gelatin, porcine skin | Sigma-Aldrich | G1890 | |
| PIPES | Sigma-Aldrich | P6757 | |
| Taxol | Sigma-Aldrich | T7191 | |
| Nocodazole | Sigma-Aldrich | M1404 | |
| Trizol | Invitrogen | 15596-026 | |
| L-Cysteine, free base | USB Corporation | 14030 | |
| Cichlid Lake Salt | Seachem | 47894 | |
| Marine salt | Seachem | SC7111 | |
| NaHCO3 | Sigma-Aldrich | S6014 | |
| EQUIPMENT | |||
| 1 ml syringes | BD Biosciences | 309659 | |
| 18 gauge needles | BD Biosciences | 305195 | |
| 30 gauge needles | BD Biosciences | 305106 | |
| Rubbermaid Plastic bucket | Amazon | 6306 | |
| Beckman Polyallomer 2 x ½ inch Ultracentrifuge tubes | Beckman | 326819 | |
| 15 ml round-bottomed glass centrifuge tubes | Fisher Scientific | 45500-15 | |
| Rubber adapter sleeves for 15 ml tubes | Kimble-Chase | 45550-15 | |
| 5 ¾ inch glass Pasteur pipettes | Fisher Scientific | 13-678-20A | |
| 14 ml polypropylene round-bottom tube | BD Biosciences | 352059 | |
| Sorvall HB-6 rotor | Thermo Scientific | 11860 | |
| Sorvall RC-6 centrifuge | Thermo Scientific | 46910 | |
References
- Martin, K. C., Ephrussi, A. mRNA localization: gene expression in the spatial dimension. Cell. 136, 719-730 (2009).
- Besse, F., Ephrussi, A. Translational control of localized mRNAs: restricting protein synthesis in space and time. Nat. Rev. Mol. Cell Biol. 9, 971-980 (2008).
- Lawrence, J. B., Singer, R. H. Intracellular localization of messenger RNAs for cytoskeletal proteins. Cell. 45, 407-415 (1986).
- Mingle, L. A., et al. Localization of all seven messenger RNAs for the actin-polymerization nucleator Arp2/3 complex in the protrusions of fibroblasts. J. Cell Sci. 118, 2425-2433 (2005).
- Blower, M. D., Feric, E., Weis, K., Heald, R. Genome-wide analysis demonstrates conserved localization of messenger RNAs to mitotic microtubules. J. Cell Biol. 179, 1365-1373 (2007).
- Eliscovich, C., Peset, I., Vernos, I., Mendez, R. Spindle-localized CPE-mediated translation controls meiotic chromosome segregation. Nat. Cell Biol. 10, 858-865 (2008).
- Sharp, J. A., Plant, J. J., Ohsumi, T. K., Borowsky, M., Blower, M. D. Functional analysis of the microtubule-interacting transcriptome. Mol. Biol Cell. 22, 4312-4323 (2011).
- Lecuyer, E., et al. Global analysis of mRNA localization reveals a prominent role in organizing cellular architecture and function. Cell. 131, 174-187 (2007).
- Mili, S., Moissoglu, K., Macara, I. G. Genome-wide screen reveals APC-associated RNAs enriched in cell protrusions. Nature. 453, 115-119 (2008).
- Holt, C. E., Bullock, S. L. Subcellular mRNA localization in animal cells and why it matters. Science. 326, 1212-1216 (2009).
- Desai, A., Murray, A., Mitchison, T. J., Walczak, C. E. The use of Xenopus egg extracts to study mitotic spindle assembly and function in vitro. Methods Cell Biol. 61, 385-412 (1999).
- Gache, V., Waridel, P., Luche, S., Shevchenko, A., Popov, A. V. Purification and mass spectrometry identification of microtubule-binding proteins from Xenopus egg extracts. Methods Mol. Med. 137, 29-43 (2007).
- Gache, V., et al. Xenopus meiotic microtubule-associated interactome. PLoS One. 5, e9248 (2010).
- Cross, M. K., Powers, M. Obtaining eggs from Xenopus laevis females. J. Vis. Exp. (18), e890 (2008).
- Cross, M. K., Powers, M. Preparation and fractionation of Xenopus laevis egg extracts. J. Vis. Exp. (18), e891 (2008).
- Cross, M., Powers, M. In vitro nuclear assembly using fractionated Xenopus egg extracts. J. Vis. Exp. (19), e908 (2008).
- Harland, R. M., Grainger, R. M. Xenopus research: metamorphosed by genetics and genomics. Trends Genet. 27, 507-515 (2011).
- Hellsten, U. The genome of the Western clawed frog Xenopus tropicalis. Science. 328, 633-636 (2010).
- Brown, K. S. Xenopus tropicalis egg extracts provide insight into scaling of the mitotic spindle. J. Cell Biol. 176, 765-770 (2007).
- Godfrey, E. W., Sanders, G. E. Effect of water hardness on oocyte quality and embryo development in the African clawed frog (Xenopus laevis). Comp. Med. 54, 170-175 (2004).
- Groisman, I., et al. CPEB, maskin, and cyclin B1 mRNA at the mitotic apparatus: implications for local translational control of cell division. Cell. 103, 435-447 (2000).
- Budde, P. P., Desai, A., Heald, R. Analysis of microtubule polymerization in vitro and during the cell cycle in Xenopus egg extracts. Methods. 38, 29-34 (2006).
- Kalab, P., Pu, R. T., Dasso, M. The ran GTPase regulates mitotic spindle assembly. Curr. Biol. 9, 481-484 (1999).
- Gaglio, T., Saredi, A., Compton, D. A. NuMA is required for the organization of microtubules into aster-like mitotic arrays. J. Cell Biol. 131, 693-708 (1995).
- Hughes, J. R., et al. A microtubule interactome: complexes with roles in cell cycle and mitosis. PLoS Biol. 6, e98 (2008).
- Suprenant, K. A., Tempero, L. B., Hammer, L. E. Association of ribosomes with in vitro assembled microtubules. Cell Motil. Cytoskeleton. 14, 401-415 (1002).
- Rebagliati, M. R., Weeks, D. L., Harvey, R. P., Melton, D. A. Identification and cloning of localized maternal RNAs from Xenopus eggs. Cell. 42, 769-777 (1985).

{kind=link}