

Summary
Vi beskriver samlingen av ubefruktet
Abstract
Mange organismer lokalisere mRNA til bestemte subcellular omgivelsen til romlig og tidsmessig kontroll genuttrykk. Nyere studier har vist at størstedelen av transcriptome er lokalisert til et nonrandom stilling i celler og embryoer. En måte å identifisere lokaliserte mRNA er å biokjemisk rense en cellestruktur av interesse og for å identifisere alle tilhørende transkripsjoner. Ved hjelp av nylig utviklede høy gjennomstrømming sekvensering teknologier er det nå enkelt å identifisere alle RNA assosiert med en subcellulære struktur. For å lette transkripsjon identifikasjon er det nødvendig å arbeide med en organisme med et fullt sekvensert genom. En attraktiv system for biokjemisk rensing av subcellular strukturer er egg ekstrakter produsert fra frosken Xenopus laevis. Men X. laevis for tiden ikke har en fullt sekvensert genomet, som vanskeliggjør transkripsjon identifikasjon. I denne artikkelen beskriver vi en metodeå produsere egg utdrag fra en relatert frosk, X. tropicalis, som har en fullt sekvensert genom. Vi gir detaljer for microtubule polymerisasjon, rensing og karakterutskrift isolasjon. Mens denne artikkelen beskriver en spesifikk metode for identifisering av mikrotubulus-assosierte transkripter, tror vi at det vil være lett brukes på andre subcellulære strukturer og vil gi en kraftig metode for identifisering av lokaliserte RNAer.
Introduction
Romlig og tidsmessig kontroll av genekspresjon er viktig for alle celler, og er spesielt viktig for kontroll av tidlig fosterdød pattering en. Romlig kontroll av genuttrykk oppnås gjennom aktiv lokalisering av mRNA til bestemte destinasjoner innenfor celler eller embryoer. I mange svært store celletyper, (f.eks egg, embryoer, og nevroner) mRNA lokalisering brukes til å begrense protein uttrykk i stedet for handling av det kodede protein. Siden en lokalisert mRNA kan katalysere mange runder med protein produksjon er det mer effektivt å lokalisere en mRNA enn å lokalisere enkelte protein molekyler. Lokaliserte mRNA er vanligvis translationally undertrykt før de når sin destinasjon, som tjener til å ytterligere begrense lokalisering av det kodede protein to. I tillegg til de mange godt dokumenterte hendelser av RNA lokalisering for å kontrollere embryonale rutemønster som har flere studier dokumentert mRNA som er lokaliserttil åsted for handlingen i den kodede protein. Prominent eksempler er lokalisering av β-actin 3 og Arp2 / 3 4 mRNA til forkanten av bevegelige fibroblaster og lokalisering av mRNA for mange mitotiske regulatorer å meiotisk og mitosetråder 5-7.
Mange av de klassiske eksemplene på lokaliserte mRNA ble identifisert gjennom genetiske skjermer for mors effekt mutasjoner og ble senere fastslått å kode lokaliserte RNA. Imidlertid har nyere genome-wide studier begynt å gi bredere innsikt i omfanget av lokaliserte RNA. En nylig in situ hybridisering skjerm i Drosophila embryo viste at ~ 70% av alle mRNA har en bestemt lokalisering, inkludert mange nye destinasjoner 8. Rensing av pseudopodia fra musefibroblaster identifisert en mangfoldig gruppe av lokaliserte mRNA ni. Arbeid fra vår gruppe ved hjelp av biokjemiske rensing av mikrotubuli fra meiotisk Xenopus egg ekstrakter identifisert hundrevis av mRNA som copurify med spindelen 5,7. Vårt arbeid viste at flertallet av microtubule-lokaliserte mRNA koder proteiner som fungerer i kontroll av mitose, støtter ideen om at mRNA er lokalisert til området av handlingen i den kodede protein. Videre fremhever evnen til å oppdage mRNA berikelse i en subcellular brøkdel av biokjemisk rensing kraften i denne tilnærmingen for identifisering av lokaliserte mRNA.
Mest lokaliserte RNA bruke aktiv transport på cytoskjelettet, enten actin eller mikrotubuli, for å oppnå transport til det endelige målet 10. For å få en bedre forståelse av omfanget og typene av RNA som er lokalisert til bestemte destinasjoner ved hjelp av en biokjemisk metode er det nødvendig å ha et in vitro-system som kan recapitulate cytoskeletal prosesser. En av de fremste systemer for å studere cytoskeletal biologi er egg ekstrakter produsertfra ubefruktede egg fra frosken Xenopus laevis. X. laevis egg ekstrakter har blitt brukt i flere tiår for å studere et bredt spekter av cytoskeletal prosesser og har bidratt mye til vår forståelse av mekanismene og molekyler som styrer cytoskeletal montering og dynamikk 11. Videre X. laevis egg ekstrakter er mottagelig for store purifications av mikrotubuli og tilhørende proteiner 12,13 og det er godt designet metoder for produksjon av ulike typer egg ekstrakter 14-16. Men for genomisk studier er det flere ulemper til bruk av X. laevis som modellsystem.
I flere tiår Xenopus laevis frosker har vært et kraftig system for studier av utviklings-og cellebiologi, på grunn av den store eggcelle størrelse og robust ytre utvikling 17. Videre utvikling av egg ekstrakt systemer som kan rekapitulere mange mobilnettet processes i et reagensrør har gjort dette frosk en kraftig eksperimentell modell. Imidlertid har Xenopus laevis vært hemmet av mangel på et komplett genom sekvens, som har blitt bremset av allotetraploid natur genome.In kontrast, har et nært beslektede arter, Xenopus tropicalis, en diploid genom som ble sekvensert i 2010 18. Mens X. tropicalis er ikke så eksperimentelt medgjørlig som X. laevis 17 tilgjengeligheten av en sekvensert genom gjør det til en attraktiv modell system for å utføre genom brede analyser.
I denne rapporten beskriver vi en metode for å gjøre meiose II-, cytostatiske factor-arrestert ekstrakter (CSF) fra X. tropicalis 19. Vi beskriver så en enkel metode for å rense mikrotubuli og tilhørende RNA'er fra denne ekstrakt. De RNA kan deretter konverteres til bibliotekene mottagelig for sekvensering bruker nyutviklede høy gjennomstrømming sekvensering teknologier. Når bibliotekeneer sekvensert de kan bli justert til genomet av frosken å identifisere spesifikke mRNA som er beriket i microtubule prøven i forhold til total ekstrakt. Dette gir en kraftig metode for å detektere microtubule-målrettet mRNA lokalisering på et genom-wide skala. I tillegg til å være i stand til å oppdage lokaliserte mRNA, bruk av high-throughput sekvensering og sekvensert genomet tilbyr muligheten for å oppdage nye vitnemål som for øyeblikket ikke er til stede i offentlig database merknader.
Protocol
En. Generasjon X. tropicalis egg
Alle Xenopus tropicalis frosker er bestilt fra NASCO. Våre frosker er plassert i en akvatiske habitater resirkulering av vann system holdt ved 27 ° C. Det er mange alternativer for vann-systemer for pleie av X. tropicalis. Noen god generell informasjon om denne frosken arter kan finnes på nettstedene til Harland and Grainger laboratorier ( http://tropicalis.berkeley.edu/home/ , http://www.faculty.virginia.edu/xtropicalis / ). Våre frosker blir opprettholdt i tankwater bestående av (0,4 g Ciclid Lake salter, 0,6 g marin salt, 0,625 g NaHCO3 pr liter vann, pH 7.0) 20. Denne oppskriften gir en ledningsevne på ~ 1800 mS, noe som er et høyt saltinnhold for X. tropicalis. Men vi har funnet ut at våre frosker trives i this miljø og eggcelle kvaliteten er forbedret. Alternative tankwater oppskrifter finner du over på de ressursene som er oppført for generell X. tropicalis omsorg.
- Frosker blir injisert med humant koriongonadotropin (hCG) på tre påfølgende dager for å stimulere egglegging: Først utarbeide to konsentrasjoner av hCG løsning. Resuspender 10.000 U av hCG lyofilisert pulver i 10 ml sterilt, avionisert H2O til en endelig konsentrasjon på 1000 U / ml. Deretter fortynnes 1 ml av 1000 U / ml hCG oppløsning i 9 ml H2O til en endelig konsentrasjon på 100 U / ml. Lagre begge løsninger ved 4 ° C.
- På dag 1, forberede 4-6 frosker for egglegging ved å injisere med hCG mellom 02:00-15:00. Injisere hver frosk i dorsal lymfe sac nær cloaca med 0,2 ml 100 U / ml hCG løsning. Å ha frosker raskt i løpet av de påfølgende to injeksjoner vil minimere mengden av frosk avfall til stede under egglegging, men er valgfritt.
- På dag to, injiserer de samme froskermed 0,2 ml 100 U / ml hCG løsning mellom 2:00 til 15:00.
- På dag 3, injiserer de samme frosker med 0,2 ml 1000 U / ml hCG løsning, mellom 07:00-10:00. Sett opp frosker å legge egg: fylle en 6-liter plastbøtte med fersk tankwater, legge til frosker og sted i mørket ved 25 ° C. Etter dette injeksjon, vil egg legging begynner etter 4 timer og vil være ferdig innen 7 hr. Frosker bør legge egg i et miljø som er opprettholdt på minimum 25 ° C.
- Gjør trekke løsninger og har utstyret klart umiddelbart før innsamling av egg.
20X MMR: 100 mMHEPES, pH 7,8, 2 mM EDTA pH 7,8, 2 M NaCl, 40 mM KCl, 20 mM MgCl2, 40 mM CaCl 2. Autoklav og oppbevares ved romtemperatur. Forbered en L 1X MMR like før trekke forberedelse.
10X XB: 100 mMHEPES, pH7.7, 10 mm MgCl 2, 1 mM CaCl 2, 1 M KCl, 500 mM sukrose. Autoklav og oppbevar ved 4 ° C. Forbered en L 1X XB jUst før trekke forberedelse. Dejelly løsning: Forbered 250 ml 3% cystein løsning i avionisert H 2 O og pH til 7.8-8.0 med 10 N NaOH. Forbered like før trekke forberedelse.
CSF-XB: ta 200 ml 1X XB og tilsett 2 ml 0,5 M EGTA pH 7,7 og 200 pl 1 M MgCl2. Forbered like før trekke forberedelse.
CSF-XB +: ta 50 ml av CSF-XB og tilsett 50 pl av LPC (10 mg / ml i hver lager leupeptin, pepstatin, og Chymostatin i DMSO). Tilsett 50 mL Cytochalasin D (10 mg / ml i DMSO). Forbered like før trekke forberedelse.
Forbered en 0,2% gelatin løsning i avionisert H 2 O, mikrobølgeovn å oppløse og filtrere sterilisere. Oppbevar ved romtemperatur.
Reserve 2 Beckman 2 x ½ tommers Ultrasentrifuger rør.
Forbered to 15 ml glass round-bunn sentrifugerør med 0,5 ml H 2 O i hver for å dempeUltrasentrifuger tube.
Gjør brann-polert glass Pasteur pipetter. Smekk slutten av av 5 ¾ tomme glass pipetter for å avsløre en bred åpning, og utsettes for ild for å jevne den nye eksponert pipette tips.
- Tilbered en 500 ml begerglass for lagring egg ved å svinge en 0,2% gelatinoppløsning rundt for å belegge veggene av begeret. Forkast gelatinoppløsning fra begerglasset etter bruk.
- Samle egg fra plastbøtte brukes til legging 6-7 time etter den tredje injeksjonen på dag tre. Hvis ønskelig, forsiktig klem hver frosk en gang for å få eventuelle gjenværende egg. Vask en gang med frisk egg tankwater og overføring til 500 ml begerglass belagt med 0,2% gelatin-løsning.
2. Utarbeidelse av Utdrag fra X. tropicalis Eggs
Alle trinnene av ekstrakt preparatet kan utføres ved romtemperatur, omtrent 25 ° C. Gjennom vaskinger, er det viktig å holde eggets senkes under væske, slik at de holdes våte. Eksponering for luft kan føre til at eggene å unnslippe cellesyklus arrest eller lyse.
- Dekanter så mye som mulig tankwater mens reservere tilstrekkelig væske til å holde eggene våt. Vippe beger inneholdende egg til siden og legge til ~ 300 ml 1X MMR langsomt til veggen av begerglasset, er slik at fysisk omrøring av eggene minimeres. La egg avgjøre, så dekanter supernatanten holdig avfall. X. tropicalis egg er trevlet på dette trinnet, slik at fjerning av aktiverte egg er gjort etter dejellying. Gjenta for totalt tre 1X MMR vasker.
- Dejelly eggene. Dekanter så mye MMR som mulig og legg halvparten av dejelly løsning. Swirl kontinuerlig i ca 5 min. Oppløsende gelé strøk vil være synlig i supernatanten etter et par minutter. Dekanter og tilsett resten dejelly løsning. Fortsett å virvle kontinuerlig til egg pakke svært tett og all orientere med sin vegetal pol (stangenmed hvitt pigment) mot bunnen av skålen. Raskt dekanter så mye dejelly løsning som mulig. Når eggene er dejellied de er svært følsomme for mekaniske manipulasjoner.
- Tilsett forsiktig XB til eggene. I den første XB vask, fjerne egg som har rømt CSF arrest ved å fjerne lysert, puffy, hvit, og pseudocleavage egg. Aktivert X. tropicalis egg tendens til å bosette seg i den øverste sentrum, så bruk en plast overføringspipetten å trekke disse ut. Også fjerne biter av hud og frosk avfall. Vask egg totalt tre ganger med ~ 300 ml 1X XB løsning, forsiktig virvlende egg mellom vasker og gi dem muligheten til å bosette seg på bunnen av begeret. Som før, dekanter så mye av hver vaskeoppløsning som mulig, og beholder egg våt.
- Vask egg to ganger med CSF-XB og dekanter.
- Legg CSF-XB + til egg. Ved hjelp av en gelatin-behandlet brann-polert Pasteur pipette, overføring egg til Ultra-sentrifugerør med CSF-XB +, tar seg ikke å avsløre eGGS til luft. Plass inne i 15 ml glass-sentrifugerør med vannet pute.
- Spinn egg i en klinisk sentrifuge ved 200 x g i 1 min, øker hastigheten til 800 xg og sentrifugering i 30 sek.
- Bruke en aspirator for å fjerne så mye som mulig fra buffer egg. De bør være nesten tørr på toppen. Raskt bevege egg til en Sorvall RC-6 sentrifuge utstyrt med en HB-6 rotor (eller tilsvarende) og spinn 17.000 xg i 15 min ved 20 ° C.
- Fjern den gule cytoplasmisk lag mellom pigment og lipid-lag ved hjelp av en 18 gauge nål festet til en 1 ml sprøyte. Punkter siden av røret og trekke sprøytesylinderen langsomt for å oppnå den cytoplasmiske ekstrakt laget. Unngå pigmentgranula så mye som mulig.
- Overføre cytoplasma til ny ultracentrifugation tube. Det er normalt at ekstrakt å vises litt regn på dette trinnet. Plass inne i 15 ml glass sentrifugerøret med vann pute. Spinn igjen 17.000 xg i 10 min ved 20 ° C. Gjenta ekstraction med 18 gauge nål.
- Overfør cytoplasma til et 1,5 ml mikro-sentrifugerør. Anslå ekstrakt volum og fortynne Cytochalasin D og LPC 1:1.000 inn ekstraktet. Bland godt med en 1 ml pipette spissen, pipettering opp og ned mange ganger uten innføring av luftbobler. En typisk avkastning fra en frisk frosk koloni er ca 300-500 mL av ekstrakt / frog. For å bevare maksimal aktivitet, er det nødvendig å lagre ekstraktet og utføre eksperimentelle manipulasjoner ved romtemperatur (20-25 ° C).
3. Rensing Taxol-stabilisert mikrotubuli fra X. tropicalis Extract
- Legg Taxol til en 100-200 pl alikvot av ekstraktet ved en endelig konsentrasjon på 10 uM og inkuber ved romtemperatur i 30 min. For kontroll reaksjoner, behandle et tilsvarende volum av ekstrakt med microtubule-destablilzing narkotika Nocodazole (10 mm). Reserve 100/il ubehandlet ekstrakt for analyse.
- Fortynne stoffet-behandletpakke med 10 volumer BRB-80 (80 mm rør pH 6,8, 1 mM MgCl2, 1 mM EGTA) + 30% glyserol. Sett sammen 14 ml rund bunn polypropylen rør som inneholder 10 ml av BRB-80 + 60% glyserol pute. Ved hjelp av et bredt hull pipette tips, lag stoffet behandlet ekstrakt reaksjon forsiktig på toppen av BRB-80 + 60% glyserol pute. Sentrifuger i 10 minutter ved 17.000 xg ved 20 ° C i en Sorvall RC-6 sentrifuge utstyrt med en HB-6 rotor (eller tilsvarende) og røradaptere.
- Aspirer supernatanten inneholdende unsedimented ekstrakt materiale, og vaske grensesnittet to ganger med avionisert H2O Sug resterende pute volumet sakte, tar seg ikke å forstyrre gel-lignende pellet inneholder mikrotubuli, microtubule-tilknyttede proteiner og microtubule-tilknyttede RNAene i Taxol-behandlet prøven. Den Nocodazole-behandlet prøve inneholder ikke synlig materiale. Resuspender pelleten i 1 ml TRIzol og fortsett med produsentens instruksjoner for å isolere RNA. Untrflere ganger for ekstrakt (opp til 100 ul) kan direkte resuspendert i 1 ml TRIzol.
- Det er nå kommersielt tilgjengelige kits for å forberede transkriptom biblioteker egnet for RNA-seq. Disse kan kjøpes gjennom http://www.illumina.com/ og http://www.454.com/ .
Representative Results
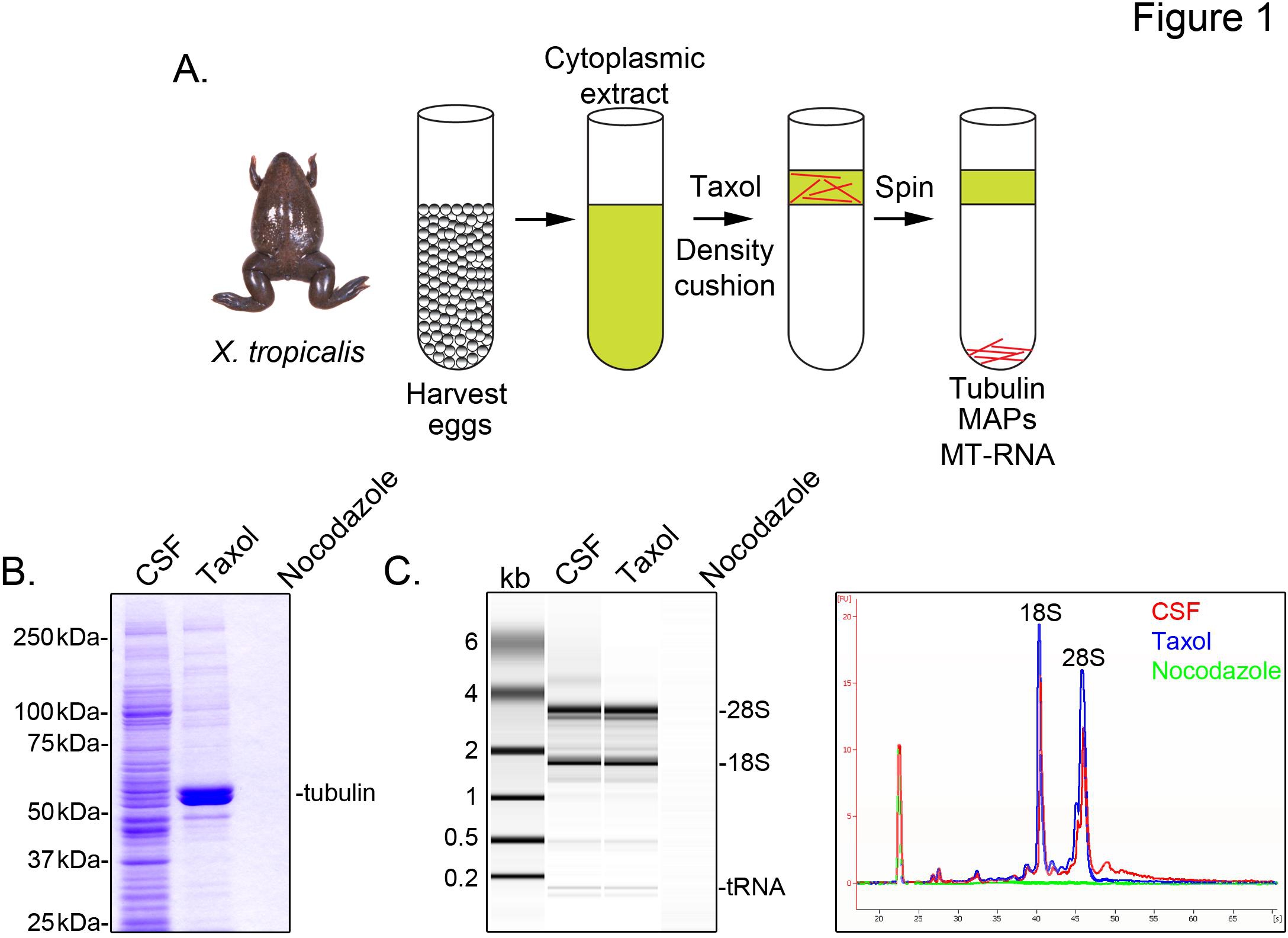
Å identifisere X. tropicalis transkripsjoner forbundet med mikrotubuli, forbereder vi en cytosolic utdrag fra ubefruktede egg arrestert i metafase av meiose II (CSF). Behandling av dette ekstrakt med taxol tillater dannelse av stabile mikrotubuli som kan renses ved sedimentering gjennom en pute glyserol (figur 1A). Coomassie gel Analysen bekrefter at α / β-tubulin sedimenter i en Noel Coward-avhengig måte, og representerer de store protein artene gjenfunnet i disse forberedelsene (figur 1B). Lavere nivåer av andre proteiner er også tilstede i den taxol pelleten, men ikke i preparatene behandlet med depolymerisering av mikrotubuli medikament nocodazole, noe som indikerer at proteiner i taxol fraksjon spesifikt forbinder med mikrotubuli (kart).
Et Agilent Bioanalyzer blir brukt til å undersøke generell RNA sammensetning i alt X. tropicalis ekstrakt fraksjoner (figur 1C X. laevis egg trekke 5,21. En linje spor av gelen projeksjon avslører mRNA signalet er merkbart lavere mikrotubulus-inneholdende taxol pellets, spesielt i regionen trekkende ovenfor 28S rRNA, som indikerer at en delmengde av mRNA cosediment med mikrotubuli i X. tropicalis. RNA isolert på denne måte er egnet for RNA-seq eksperimenter ved hjelp av kommersielt tilgjengelige reagenser.

Figur 1. Rensing av MT-RNA i RNA-ff. (A) Rensing ordningen for å isolere MT-RNA. Egg er høstet fra kvinnelige X. tropicalis frosker. Etter fremstilling av en cytoplasmisk ekstrakt, er taxol tilsatt for å fremkalle mikrotubuli-polymerisering. Mikrotubuli-og MT-RNA blir renset ved sedimentering gjennom en pute glyserol (B). Coomassie gelanalyse av proteiner isolert ved hjelp av skjemaet beskrevet i (A). Totalt ekstrakt CSF sammenlignet med proteiner sedimenteres i nærvær av taxol eller nocodazole. (C) Bioanalyzer gelanalyse av RNA isolert ved hjelp av skjemaet beskrevet i (A). RNA isolert fra CSF ekstrakt i forhold til RNA sedimenteres i nærvær av taxol eller nocodazole. Både gel projeksjon og linjen spor er vist. Gjengitt med tillatelse fra Sharp, et al., (2011). Klikk her for å se større bilde .
Discussion
I denne rapporten har vi beskrevet en enkel metode for å produsere CSF-arrestert egg ekstrakter fra X. tropicalis 19 og bruke dette ekstraktet for å studere microtubule-tilknyttede RNA 7. Den grunnleggende prosedyren for å produsere CSF-arrestert egg ekstrakter fra X. tropicalis er det samme som anvendt for X. laevis med noen viktige forskjeller. En av de mest utfordrende sidene til å jobbe med X. tropicalis frosker er å skaffe nok høy kvalitet egg for å lage et utdrag med microtubule nucleation eller spindelenheten aktivitet sammenlignes med X. laevis egg ekstrakter. For å oppnå optimale egglegging forhold mens hindre glidning fra meiose II cellesyklus arrest, intervallet mellom hormon injeksjoner for X. tropicalis er kortere enn det som brukes for X. laevis, og tidspunktet fra den tredje hCG-injeksjonen til begynnelsen av egglegging er også mye kortere. Med X. laevis timingen fra hCG-injeksjonen til than begynnelsen av det å legge egg er slik at det er praktisk og effektiv for egg for å legges over natten i buffer. Imidlertid, på grunn av den kortere tid mellom hCG-injeksjonen og egglegging med X. tropicalis er det ofte nødvendig å manuelt uttrykke egg fra frosker. En annen vesentlig forskjell mellom å gjøre egg ekstrakt fra de to forskjellige frosker er dejellying trinn. Med X. laevis eggene er så stor at det er lett å avgjøre når gelé pelsen er oppløst ved å observere hvor nært eggene er plassert i begeret. Som dejellying reaksjon starter, eggene begynner å pakke tettere. Men X. tropicalis egg er mye mindre og det kan være ganske vanskelig å avgjøre når gelé coat har oppløst av egg pakking tetthet alene. Vi har funnet ut at den mest pålitelige metoden for å bestemme når gelé strøk er oppløst, er å overvåke retningen på dyret (svart) og vegetal (hvit) polene. Når alle vegetal stolper orient mot bunnen av begerglasset gelé belegge er fjernet nok til å fortsette med ekstraktet. Til slutt, mens X. laevis egg ekstrakt kan lagres på kjølige temperaturer (4-12 ° C) har vi observert at det er avgjørende å opprettholde X. tropicalis egg ekstrakt ved romtemperatur (20-25 ° C) i løpet av fremstilling og eksperimentelle manipulasjoner for å bevare biokjemisk aktivitet. På grunn av forskjellene i brukervennlighet foretrekker vi å bruke X. laevis frosker for produksjon av egg ekstrakt. Men for eksperimenter som krever eller er tilrettelagt av en organisme med en koplende genom, X. tropicalis er et utmerket alternativ system.
Metoden som vi har beskrevet i denne rapporten bruker Noel Coward som en microtubule-stabiliserende middel for å indusere microtubule polymerisasjon. Vi valgte denne metoden fordi taxol er en robust mikrotubulus-stabiliserende middel som muliggjør storskala isolering av renset mikrotubuli. Metoden thpå vi beskrev kunne trolig bli forbedret ved å sammenligne proteiner og RNAene forbundet med mikrotubuli ved hjelp av alternative microtubule polymerisasjonsmetoder. Alternativer kan omfatte polymerisering hjelp GTP-indusert polymerisasjon (en klassisk teknikk), 22 eller ved hjelp av Ran-GTP som microtubule polymerizer å etterligne mikrotubuli indusert av kromatin-drevet spindelenheten 23. Til slutt, bruk av renset sperm kjerner å indusere microtubule polymerisasjon vil være det nærmeste etterligne til hvilke typer mikrotubuli som nukleerte under mitose (sentrosomen, kromatin, og kinetochore mediert). Ulemper med disse alternative kilder av mikrotubulus kjernedannelse er at de kjernedannende midler er ikke så lett tilgjengelig som Taxol og de ikke kjernekoking eller stabiliserer mikrotubuli så effektivt som taxol. Derfor ville hver av disse metodene være vanskeligere å bruke for store purifications. Fordelen ved å sammenligne flere forskjellige typer mikrotubulidynamikk nukleatorerer at det kan være mulig å identifisere proteiner og / eller RNA'er som er spesifikke for hver vei av mikrotubuli kjernedannelse.
Metoden som vi har beskrevet her tar nytte av cytoplasma ekstrakter av amfibier. Imidlertid kan denne metoden bli utvidet til bruken av ekstrakt system fra andre organismer. Mitotisk ekstrakter har blitt beskrevet fra synkroniserte menneskelig vev kultur celler 24 som trofast rekapitulere mange aspekter av microtubule montering. Vi har med hell brukt disse ekstrakter å identifisere microtubule-tilknyttede RNA fra Jakten celler fem. Lignende microtubule rensing ordninger har blitt beskrevet i mange forskjellige organismer 25,26, selv om microtubule forbundet RNA har ikke blitt undersøkt. Fremgangsmåten beskrevet her kan bli brukt med hvilken som helst organisme som kan produsere et konsentrert cytoplasmisk ekstrakt stand kjernedannende mikrotubuli.
Til slutt, selv om den fremgangsmåte som vi deskriftlærd her diskuterer rensing av mikrotubuli og tilhørende proteiner og RNA, kan denne tilnærmingen være generaliseres til andre subcellular strukturer. Mens de fleste lokaliserte mRNA ikke har blitt identifisert ved hjelp av biokjemiske metoder de siste fremskritt innen DNA og RNA sekvensering teknologier gjør denne tilnærmingen en attraktiv metode for å identifisere lokaliserte RNA. I denne framgangsmåten være subcellulære eller sub-embryo strukturen av interesse kan bli isolert eller renset. Da de tilknyttede proteiner og RNA'er kan identifiseres på et genom bred skala. RNA kan deretter sammenlignes med den RNA-innholdet av det totale celle eller embryo for å identifisere anrikede lokaliserte RNAer. Denne tilnærmingen kan brukes med hele egg (dyr og vegetal separasjon, lik den tilnærmingen som identifiserte de første lokaliserte RNA i Xenopus 27), aktin tilknyttede RNAene, ER-tilknyttede RNAene, mitokondrier-tilknyttede RNAene, eller noen subcellular struktur som kan bli renset med tilhørende RNA intakt. Basert påvårt arbeid på microtubule-assosiert RNA vi spår at dette ville være en utmerket metode for å oppdage nye proteiner som fungerer på et gitt sted. Videre vil identifikasjon av lokalisering og omfang av alle lokaliserte RNA gi innsikt i hvordan celler og embryoer bruke mRNA lokalisering for å kontrollere genuttrykk.
Disclosures
Ingen interessekonflikter erklært.
Materials
| Name | Company | Catalog Number | Comments |
| REAGENTS | |||
| Xenopus tropicalis | NASCO | LM00823MX | |
| human Chorionic Gonadotropin | Sigma-Aldrich | CG10 | |
| HEPES | Sigma-Aldrich | H4034 | |
| EDTA | Sigma-Aldrich | E5134 | |
| NaCl | Sigma-Aldrich | S3014 | |
| KCl | Sigma-Aldrich | P9541 | |
| MgCl2 | Sigma-Aldrich | M8266 | |
| CaCl2 | Sigma-Aldrich | C8106 | |
| sucrose | Sigma-Aldrich | S0389 | |
| NaOH | Sigma-Aldrich | S5881 | |
| EGTA | Sigma-Aldrich | E3889 | |
| Leupeptin | Sigma-Aldrich | L9783 | |
| Pepstatin | Sigma-Aldrich | P5318 | |
| Chymostatin | Sigma-Aldrich | C7268 | |
| Cytochalasin D | Sigma-Aldrich | C8273 | |
| Gelatin, porcine skin | Sigma-Aldrich | G1890 | |
| PIPES | Sigma-Aldrich | P6757 | |
| Taxol | Sigma-Aldrich | T7191 | |
| Nocodazole | Sigma-Aldrich | M1404 | |
| Trizol | Invitrogen | 15596-026 | |
| L-Cysteine, free base | USB Corporation | 14030 | |
| Cichlid Lake Salt | Seachem | 47894 | |
| Marine salt | Seachem | SC7111 | |
| NaHCO3 | Sigma-Aldrich | S6014 | |
| EQUIPMENT | |||
| 1 ml syringes | BD Biosciences | 309659 | |
| 18 gauge needles | BD Biosciences | 305195 | |
| 30 gauge needles | BD Biosciences | 305106 | |
| Rubbermaid Plastic bucket | Amazon | 6306 | |
| Beckman Polyallomer 2 x ½ inch Ultracentrifuge tubes | Beckman | 326819 | |
| 15 ml round-bottomed glass centrifuge tubes | Fisher Scientific | 45500-15 | |
| Rubber adapter sleeves for 15 ml tubes | Kimble-Chase | 45550-15 | |
| 5 ¾ inch glass Pasteur pipettes | Fisher Scientific | 13-678-20A | |
| 14 ml polypropylene round-bottom tube | BD Biosciences | 352059 | |
| Sorvall HB-6 rotor | Thermo Scientific | 11860 | |
| Sorvall RC-6 centrifuge | Thermo Scientific | 46910 | |
References
- Martin, K. C., Ephrussi, A. mRNA localization: gene expression in the spatial dimension. Cell. 136, 719-730 (2009).
- Besse, F., Ephrussi, A. Translational control of localized mRNAs: restricting protein synthesis in space and time. Nat. Rev. Mol. Cell Biol. 9, 971-980 (2008).
- Lawrence, J. B., Singer, R. H. Intracellular localization of messenger RNAs for cytoskeletal proteins. Cell. 45, 407-415 (1986).
- Mingle, L. A., et al. Localization of all seven messenger RNAs for the actin-polymerization nucleator Arp2/3 complex in the protrusions of fibroblasts. J. Cell Sci. 118, 2425-2433 (2005).
- Blower, M. D., Feric, E., Weis, K., Heald, R. Genome-wide analysis demonstrates conserved localization of messenger RNAs to mitotic microtubules. J. Cell Biol. 179, 1365-1373 (2007).
- Eliscovich, C., Peset, I., Vernos, I., Mendez, R. Spindle-localized CPE-mediated translation controls meiotic chromosome segregation. Nat. Cell Biol. 10, 858-865 (2008).
- Sharp, J. A., Plant, J. J., Ohsumi, T. K., Borowsky, M., Blower, M. D.
- Lecuyer, E., et al. Global analysis of mRNA localization reveals a prominent role in organizing cellular architecture and function. Cell. 131, 174-187 (2007).
- Mili, S., Moissoglu, K., Macara, I. G. Genome-wide screen reveals APC-associated RNAs enriched in cell protrusions. Nature. 453, 115-119 (2008).
- Holt, C. E., Bullock, S. L. Subcellular mRNA localization in animal cells and why it matters. Science. 326, 1212-1216 (2009).
- Desai, A., Murray, A., Mitchison, T. J., Walczak, C. E. The use of Xenopus egg extracts to study mitotic spindle assembly and function in vitro. Methods Cell Biol. 61, 385-412 (1999).
- Gache, V., Waridel, P., Luche, S., Shevchenko, A., Popov, A. V. Purification and mass spectrometry identification of microtubule-binding proteins from Xenopus egg extracts. Methods Mol. Med. 137, 29-43 (2007).
- Gache, V., et al.
- Cross, M. K., Powers, M. Obtaining eggs from Xenopus laevis females. J. Vis. Exp. (18), e890 (2008).
- Cross, M. K., Powers, M. Preparation and fractionation of Xenopus laevis egg extracts. J. Vis. Exp. (18), e891 (2008).
- Cross, M., Powers, M. In vitro nuclear assembly using fractionated Xenopus egg extracts. J. Vis. Exp. (19), e908 (2008).
- Harland, R. M., Grainger, R. M. Xenopus research: metamorphosed by genetics and genomics. Trends Genet. 27, 507-515 (2011).
- Hellsten, U. The genome of the Western clawed frog Xenopus tropicalis. Science. 328, 633-636 (2010).
- Brown, K. S. Xenopus tropicalis egg extracts provide insight into scaling of the mitotic spindle. J. Cell Biol. 176, 765-770 (2007).
- Godfrey, E. W., Sanders, G. E. Effect of water hardness on oocyte quality and embryo development in the African clawed frog (Xenopus laevis). Comp. Med. 54, 170-175 (2004).
- Groisman, I., et al. CPEB, maskin, and cyclin B1 mRNA at the mitotic apparatus: implications for local translational control of cell division. Cell. 103, 435-447 (2000).
- Budde, P. P., Desai, A., Heald, R. Analysis of microtubule polymerization in vitro and during the cell cycle in Xenopus egg extracts. Methods. 38, 29-34 (2006).
- Kalab, P., Pu, R. T., Dasso, M. The ran GTPase regulates mitotic spindle assembly. Curr. Biol. 9, 481-484 (1999).
- Gaglio, T., Saredi, A., Compton, D. A. NuMA is required for the organization of microtubules into aster-like mitotic arrays. J. Cell Biol. 131, 693-708 (1995).
- Hughes, J. R., et al. A microtubule interactome: complexes with roles in cell cycle and mitosis. PLoS Biol. 6, e98 (2008).
- Suprenant, K. A., Tempero, L. B., Hammer, L. E. Association of ribosomes with in vitro assembled microtubules. Cell Motil. Cytoskeleton. 14, 401-415 (1002).
- Rebagliati, M. R., Weeks, D. L., Harvey, R. P., Melton, D. A. Identification and cloning of localized maternal RNAs from Xenopus eggs. Cell. 42, 769-777 (1985).

{kind=link}