

Summary
Vi beskriver indsamlingen af ubefrugtede
Abstract
Mange organismer lokalisere mRNA til specifikke subcellulære omgivelser rumligt og tidsligt kontrol genekspression. Nylige undersøgelser har vist, at størstedelen af transkriptomet er lokaliseret til en tilfældig position i celler og embryoer. En metode til at identificere lokaliserede mRNA er at biokemisk rense en cellestruktur af interesse og for at identificere alle tilhørende udskrifter. Brug af nyligt udviklede high-throughput sekventering teknologier er det nu ligetil at identificere alle RNA'er associeret med en subcellulær struktur. For at lette udskrift identifikation er det nødvendigt at arbejde med en organisme med en fuldt sekventeret genom. Én attraktivt system til biokemisk oprensning af subcellulære strukturer er æg uddrag fremstillet frøen Xenopus laevis. Men X. laevis øjeblikket ikke har et fuldstændigt genom, hvilket hæmmer udskrift identifikation. I denne artikel beskriver vi en metodetil at producere æg uddrag fra en beslægtet frø, X. tropicalis, der har et fuldt sekventeret genom. Vi leverer oplysninger til mikrotubuli polymerisation, rensning og udskrift isolation. Mens denne artikel beskrives en specifik metode til identifikation af mikrotubuli-associerede udskrifter, mener vi, at det vil være let anvendes på andre subcellulære strukturer og vil give en kraftfuld metode til identifikation af lokaliserede RNA'er.
Introduction
Rumlig og tidsmæssig kontrol af genekspression er vigtig for alle celler, og er specielt vigtigt for kontrol af tidlige embryonale pattering 1. Spatial kontrol af genekspression opnås gennem aktiv lokalisering af mRNA til bestemte destinationer inden celler eller embryoner. I mange meget store celletyper mRNA lokalisering (f.eks oocytter, embryoner, og neuroner) anvendes til at begrænse proteinekspression til virkningsstedet af det kodede protein. Da en lokaliseret mRNA kan katalysere mange runder af proteinproduktion det er mere effektivt at lokalisere et mRNA end at lokalisere individuelle proteinmolekyler. Lokaliserede mRNA er typisk translationally undertrykt, indtil de når deres bestemmelsessted, som tjener til yderligere at begrænse lokalisering af kodede protein 2. Ud over de mange veldokumenterede tilfælde af RNA lokalisering at kontrollere embryonale mønster, har flere undersøgelser dokumenterede mRNA'er der er lokaliserettil stedet for virkningen af det kodede protein. Fremtrædende eksempler indbefatter lokalisering af β-actin-3 og Arp2 / 3 4 mRNA'er til forkanten af bevægelige fibroblaster og lokalisering af mRNA'erne for mange mitotiske regulatorer til meiotiske og mitotiske spindler 5-7.
Mange af de klassiske eksempler på lokaliserede mRNAer blev identificeret gennem genetiske skærme til maternelle effekt mutationer og blev senere besluttet at indkode lokaliserede RNA. Imidlertid har de seneste genom-dækkende undersøgelser er begyndt at give bredere indblik i omfanget af lokaliserede RNA'er. En nylig in situ hybridisering skærm i Drosophila embryoer viste, at ~ 70% af alle mRNA'er har en specifik lokalisering, herunder mange nye destinationer 8.. Oprensning af pseudopodia fra musefibroblaster identificeret en forskelligartet gruppe af lokaliserede mRNA 9.. Arbejde fra vores gruppe ved hjælp biokemisk oprensning af mikrotubuli fra meiotic Xenopus æg udvinder identificeret hundredvis af mRNA, der copurify med spindelen 5,7. Vores arbejde viste, at hovedparten af mikrotubulus-lokaliserede mRNA'er koder for proteiner, der fungerer i kontrol i mitosen, støtter tanken om, at mRNA'er er lokaliseret til stedet for virkningen af det kodede protein. Desuden evnen til at detektere mRNA berigelse i en subcellulær fraktion ved biokemisk oprensning fremhæver kraften af denne fremgangsmåde til identifikation af lokaliserede mRNA'er.
De fleste lokaliserede RNA bruger aktiv transport på cytoskelettet enten actin eller mikrotubuli, for at opnå transport til deres endelige bestemmelsessted 10.. For at få en bedre forståelse af omfanget og typer af RNA, der er lokaliseret til bestemte destinationer via en biokemisk tilgang er det nødvendigt at have et in vitro system, der kan rekapitulere cytoskeletale processer. En af de førende systemer til at studere cytoskeletal biologi er æg produceret uddragfra ubefrugtede æg fra frøen Xenopus laevis. X. laevis æg ekstrakter har været brugt i årtier for at studere en bred vifte af cytoskeletale processer og har bidraget meget til vores forståelse af de mekanismer og molekyler, der styrer cytoskeletal montage og dynamik 11.. Desuden X. laevis æg ekstrakter er egnede til store oprensninger af mikrotubuli og tilhørende proteiner 12,13 og der er veldesignede metoder til produktion af forskellige typer af æg ekstrakter 14-16. Men for genomiske undersøgelser er der flere ulemper ved anvendelse af X. laevis som et modelsystem.
I årtier Xenopus laevis frøer har været et kraftfuldt system til undersøgelse af udviklingstoksicitet og cellebiologi, på grund af den store oocyte størrelse og robust ydre udvikling 17.. Desuden har opbygningen af æggeekstrakt systemer, der kan rekapitulere mange cellulære processes i et reagensglas har gjort denne frøen en kraftig eksperimentel model. Imidlertid har Xenopus laevis været hæmmet af manglen på en fuldstændig genomsekvens, som er blevet bremset af den allotetraploid karakter genome.In kontrast et nært beslægtet art, Xenopus tropicalis, har en diploid genom, der blev sekventeret i 2010 18. Mens X. tropicalis er ikke så eksperimentelt medgørlig som X. laevis 17 tilgængeligheden af en genom gør det til et attraktivt modelsystem til at udføre genom analyser.
I denne rapport beskriver vi en metode til at gøre meiose II, cytostatika factor-anholdte ekstrakter (CSF) fra X. tropicalis 19.. Vi derefter beskrive en simpel metode til at rense mikrotubuli og tilhørende RNA'er fra dette ekstrakt. RNA'erne kan derefter omdannes til bibliotekerne kan underkastes sekventering hjælp nyligt udviklede high throughput sekventering teknologier. Når bibliotekernesekventeres de kan justeres til genomet af frøen at identificere specifikke mRNA'er, som er beriget i mikrotubulus prøve forhold til den samlede ekstrakt. Dette giver en stærk metode til at opdage mikrotubuli målrettet mRNA lokalisering på et genom-plan. Ud over at være i stand til at detektere lokaliserede mRNA'er, tilbyde brug af high-throughput sekventering og et genom muligheden for at opdage nye transkripter, der ikke i øjeblikket til stede i offentlige database anmærkninger.
Protocol
1.. Generering af X. tropicalis Æg
Alle Xenopus tropicalis frøer bestilles fra NASCO. Vore frøer er opstaldet i et akvatisk habitat recirkulerende vandsystem holdes på 27 ° C. Der er mange muligheder for vand-systemer til pleje af X. tropicalis. Nogle gode generelle oplysninger om denne frø arter kan findes på hjemmesider for Harland og Grainger labs ( http://tropicalis.berkeley.edu/home/ , http://www.faculty.virginia.edu/xtropicalis / ). Vore frøer holdes i tankwater bestående af (0,4 g Ciclid Lake Salts 0,6 g havsalt, 0,625 g NaHCO3 per liter vand, pH 7,0) 20. Denne opskrift resulterer i en ledningsevne på ~ 1800 uS, hvilket er en høj saltholdighed for X. tropicalis. Vi har imidlertid fundet, at vores frøer trives i This miljø og oocyt kvalitet er forbedret. Alternative tankwater opskrifter kan findes over på de ressourcer, der er anført til generel X. tropicalis pleje.
- Frøer injiceres med humant choriongonadotropin (hCG) på tre på hinanden følgende dage for at stimulere æglægning: Først forberede to koncentrationer af hCG løsning. Resuspender 10.000 E lyofiliseret hCG pulver i 10 ml sterilt, deioniseret H2O til en endelig koncentration på 1,000 U / ml. Derefter fortyndes 1 ml af 1.000 U / ml hCG opløsning i 9 ml H2O til en endelig koncentration på 100 U / ml. Opbevar begge løsninger ved 4 ° C.
- På dag 1, forberede 4-6 frøer til æglægning ved injektion med hCG mellem 2:00 til 03:00. Indsprøjtes hver frø i den dorsale lymfe vej tæt kloakken med 0,2 ml 100 U / ml hCG opløsning. Under frøerne hurtigt i de efterfølgende to injektioner vil minimere mængden af frøen affald til stede under æglægning, men er valgfrit.
- På dag 2 injicere de samme frøermed 0,2 ml 100 U / ml hCG opløsning mellem 2:00 til 3:00.
- På dag 3, injiceres de samme frøer med 0,2 ml 1,000 U / ml hCG opløsning, mellem 7:00 til 10:00. Opsæt frøer at lægge æg: fylde en 6-Quart plastikspand med frisk tankwater, tilsæt frøer og sted i mørke ved 25 ° C. Efter denne injektion, vil æglægning begynder efter 4 timer og vil være komplet med 7 timer. Frøer bør lægge æg i et miljø, der holdes på en minimum 25 ° C.
- Gør ekstrakt løsninger og har udstyr klar umiddelbart før indsamling af æg.
20X MMR: 100 mM HEPES, pH 7,8, 2 mM EDTA pH 7,8, 2 M NaCl, 40 mM KCI, 20 mM MgCl2, 40 mM CaCl2. Autoklave og opbevares ved stuetemperatur. Klargør 1 L 1X MMR lige før udtrække forberedelse.
10X XB: 100 mM HEPES, pH7.7, 10 mM MgCl2, 1 mM CaCl2, 1 M KCl, 500 mM sucrose. Autoklave og opbevares ved 4 ° C. Klargør 1 L 1X XB just før udtrække forberedelse. Dejelly løsning: Forbered 250 ml 3% cystein opløsning i demineraliseret H 2 O og pH til 7,8-8,0 med 10 N NaOH. Forbered lige før udtrække forberedelse.
CSF-XB: Tag 200 ml 1X XB og tilsæt 2 ml 0,5 M EGTA pH 7,7 og 200 ul 1 M MgCl2. Forbered lige før udtrække forberedelse.
CSF-XB +: Tag 50 ml CSF-XB og der tilsættes 50 ul LPC (10 mg / ml hver bestand af Leupeptin, Pepstatin og Chymostatin i DMSO). Tilsæt 50 gl Cytochalasin D (10 mg / ml i DMSO). Forbered lige før udtrække forberedelse.
Forbered en 0,2% gelatine-opløsning i demineraliseret H 2 O, mikrobølgeovn til at opløse og filtrere sterilisere. Opbevares ved stuetemperatur.
Reserve 2 Beckman 2 x ½ tomme ultracentrifugerør.
Forbered to 15 ml glas rundbundede centrifugeglas med 0,5 ml H 2 O i hver for at afbødeultracentrifugerør.
Gør flammepoleret glas Pasteur-pipetter. Snap slutningen off af 5 ¾ tommer glaspipetter at afsløre en bred åbning, og udsættes for ild at udjævne nye udsatte pipette spids.
- Forbered en 500 ml bægerglas til opbevaring af æg ved hvirvlende en 0,2% gelatineopløsning rundt at overtrække væggene i bægerglasset. Kassér gelatineopløsning fra bægerglas efter brug.
- Indsamle æg fra plasticspand bruges til udlægning 6-7 timer efter den tredje injektion på dag 3. Hvis det ønskes, forsigtigt klemme hver frøen gang for at få de resterende æg. Vask æggene gang med frisk tankwater og overføres til en 500 ml bægerglas belagt med 0,2% gelatine løsning.
2.. Fremstilling af ekstrakt fra X. tropicalis Æg
Alle trin af ekstrakt præparat kan udføres ved stuetemperatur, omkring 25 ° C. Gennem vask, er det vigtigt at holde ægs neddykket under flydende, så de forbliver våd. Udsættelse for luft kan medføre æggene at undslippe cellecyklusstop eller lysere.
- Dekanteres så meget tankwater som muligt, mens forbeholde væske nok til at holde æggene våde. Vip bægerglas, der indeholder æg til siden og tilføje ~ 300 ml 1X MFR langsomt til væggen på bægeret er så fysisk agitation af æggene minimeres. Lad æg afregne, så afhældes supernatant indeholdende snavs. X. tropicalis æg er trævlet på dette trin, så fjernelse af aktiverede æg sker efter dejellying. Gentag for i alt tre 1X MFR vaske.
- Dejelly æggene. Afhældes så meget MFR som muligt og tilføje halvdelen af dejelly opløsningen. Swirl kontinuerligt i ca 5 min. Opløsning gelé frakker vil være synlige i supernatanten efter et par minutter. Afhældes og tilsæt den resterende dejelly løsningen. Fortsæt med at hvirvle uafbrudt, indtil æggene pack meget stramt, og alle orientere med deres vegetal pol (polmed hvidt pigment) mod bunden af skålen. Hurtigt dekanteres off så meget dejelly løsning som muligt. Når æggene er dejellied de er meget følsomme over for mekaniske manipulationer.
- Derefter tilsættes forsigtigt XB til æggene. I den første XB vask fjerne æg, der er flygtet CSF anholdelse ved at fjerne lyserede, puffy, hvid og pseudocleavage æg. Aktiveret X. tropicalis æg tendens til at bosætte sig i den øverste centrum, så brug en plastik overførselspipetten at trække disse ud. Også fjerne stykker af hud og frøen affald. Vask æg i alt tre gange med ~ 300 ml 1X XB-løsning, blidt hvirvlende æg mellem vaske og tillader dem at slå sig ned på bunden af bægerglasset. Som før, dekanteres så meget af hver vandopløsning som muligt og samtidig holde æg våd.
- Vask æggene to gange med CSF-XB og dekanteres.
- Tilføj CSF-XB + til æg. Ved hjælp af en gelatine-behandlet flammepoleret Pasteur pipette æg til Ultra-centrifugeglas med CSF-XB +, pas på ikke at udsætte eGGS til luft. Sted inde i 15 ml glascentrifugerør med vand pude.
- Spin æg i en klinisk centrifuge ved 200 xg i 1 min, øge hastigheden til 800 xg og spin for 30 sek.
- Brug et sug til at fjerne så meget buffer som muligt fra æg. De bør være næsten tørt på toppen. Hurtigt flytte æg til en Sorvall RC-6 centrifuge udstyret med en HB-6 rotor (eller tilsvarende) og spin 17.000 xg i 15 minutter ved 20 ° C.
- Fjern den gule cytoplasmatiske lag mellem pigment og lipidlag med en 18 gauge nål fastgjort til en 1 ml sprøjte. Punktere side af røret og træk sprøjtecylinderen langsomt at opnå den cytoplasmiske ekstrakt lag. Undgå pigmentgranula så meget som muligt.
- Overfør cytoplasma til ny ultracentrifugering rør. Det er normalt, at ekstrakt til se en anelse uklar på dette trin. Sted inden for 15 ml glas centrifugerør med vand pude. Spinde 17.000 xg i 10 minutter ved 20 ° C. Gentag ext-fraktion med 18 gauge nål.
- Overførsel cytoplasma til et 1,5 ml mikrofugerør. Skøn ekstrakt volumen og fortynd Cytochalasin D og LPC 1:1000 ind ekstraktet. Bland godt med en 1 ml pipettespids, pipettering op og ned mange gange uden indføring af luftbobler. En typisk udbytte fra et sundt frog koloni er ca 300-500 ul ekstrakt / frog. At bevare maksimal aktivitet, er det nødvendigt at opbevare ekstraktet og udføre eksperimentelle manipulationer ved stuetemperatur (20-25 ° C).
3.. Rensning Taxol-stabiliserede mikrotubuli fra X. tropicalis Extract
- Tilføj Taxol til en 100-200 ml alikvot af ekstraktet på en endelig koncentration på 10 uM og inkuber ved stuetemperatur i 30 min. For kontrolreaktioner, behandle en tilsvarende mængde ekstrakt med mikrotubuli destablilzing drug Nocodazole (10 uM). Reserve 100 ul ubehandlet ekstrakt til analyse.
- Fortynd den lægemiddel-behandledeekstraheres med 10 volumener BRB-80 (80 mM PIPES pH 6,8, 1 mM MgCl2, 1 mM EGTA) + 30% glycerol. Saml 14 ml rundbundede polypropylen rør indeholdende 10 ml BRB-80 + 60% glycerol pude. Ved hjælp af en bred boring pipettespids lag lægemiddel-behandlede ekstrakt reaktion forsigtigt på toppen af BRB-80 + 60% glycerolpude. Centrifuger i 10 minutter ved 17.000 xg ved 20 ° C i en Sorvall RC-6 centrifuge udstyret med en HB-6 rotor (eller tilsvarende) og røradaptorer.
- Aspirer supernatanten indeholdende unsedimented ekstrakt materiale, og vask grænsefladen to gange med deioniseret H2O Aspirer den resterende pude volumen langsomt, pas på ikke at forstyrre gel-lignende pellet indeholdende mikrotubuli, mikrotubuli-associerede proteiner og mikrotubuli-associerede RNA'er i taxolbehandlede prøve. Den Nocodazole-behandlede prøve ikke indeholder synlige materiale. Pellet resuspenderes i 1 ml TRIzol og fortsætte med producentens anvisninger for at isolere RNA. Untrprettet ekstrakt (op til 100 ul) kan resuspenderes direkte i 1 ml TRlzol.
- Der er nu kommercielt tilgængelige kits til fremstilling Transkriptomet biblioteker er egnede til RNA-seq. Disse kan købes gennem http://www.illumina.com/ og http://www.454.com/ .
Representative Results
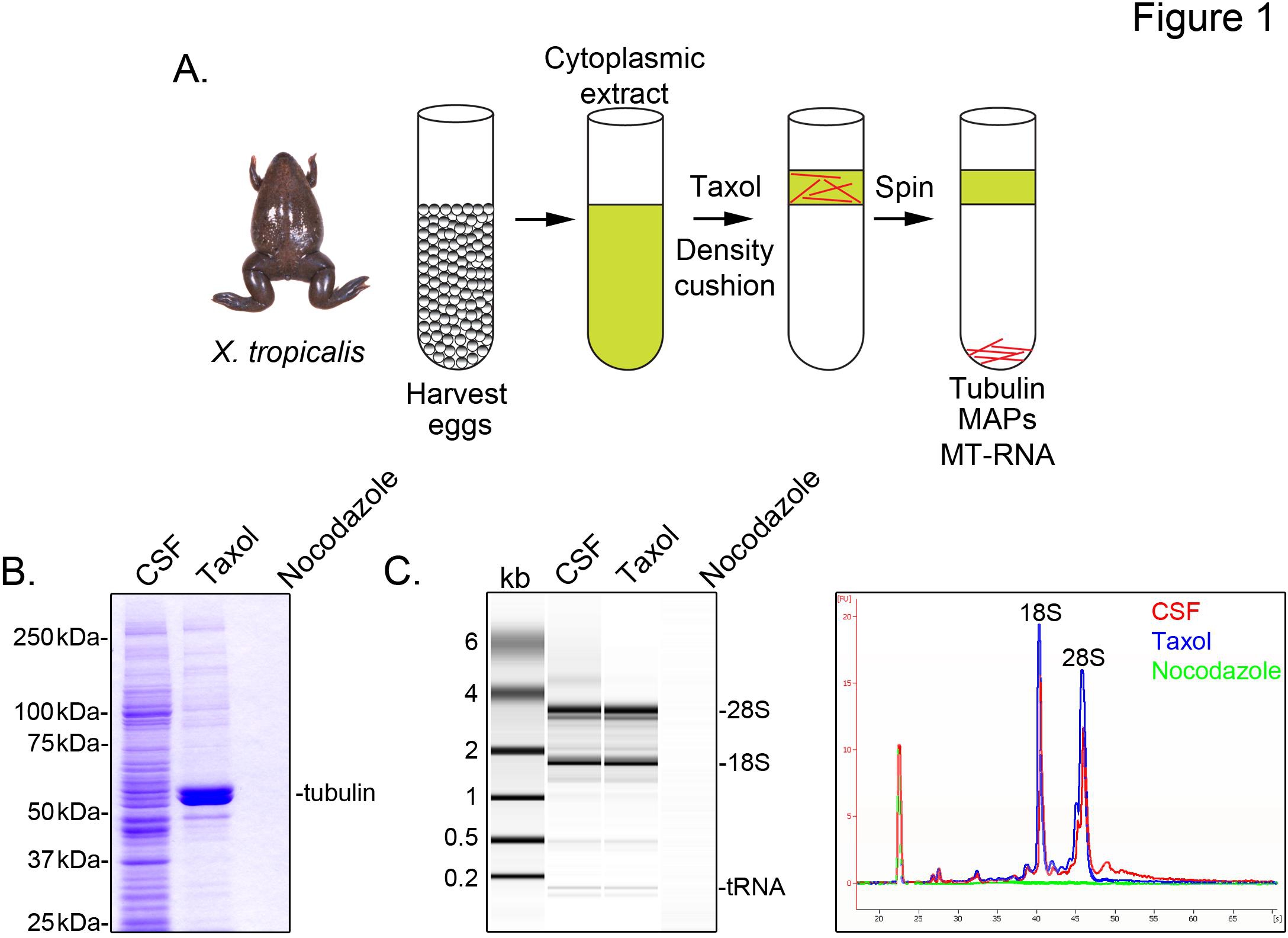
At identificere X. tropicalis udskrifter forbundet med mikrotubuli, vi forbereder et cytosol uddrag ubefrugtede æg, arresteret i metafase af meiosen II (CSF). Behandling af denne ekstrakt med taxol tillader dannelsen af stabile mikrotubuli som kan renses ved sedimentering gennem en glycerolpude (figur 1A). Coomassie gel analyse bekræfter, at α / β-tubulin sedimenter i en Taxol-afhængig måde, og repræsenterer de store proteinspecies inddrevet i disse præparater (figur 1B). Lavere niveauer af andre proteiner er også til stede i taxol pellet, men ikke i præparater behandlet med mikrotubuli depolymerisering lægemiddel nocodazol, hvilket indikerer, at proteiner i Taxol fraktion specifikt forbinder med mikrotubuli (kort).
En Agilent Bioanalyzer bruges til at undersøge generelle RNA komposition i alle X. tropicalis ekstrakt fraktioner (figur 1C X. laevis æg ekstrakt 5,21. En linje spor af gelen projektion afslører mRNA-signalet er markant lavere i mikrotubulus-indeholdende taxol pellet, især i regionen migrerer over 28S rRNA, hvilket indikerer, at en delmængde af mRNA'er cosediment med mikrotubuli i X. tropicalis. RNA isoleret på denne måde er egnet til RNA-seq eksperimenter under anvendelse af kommercielt tilgængelige reagenser.

Figur 1. Oprensning af MT-RNA for RNA-seq. (A) Oprensning ordning at isolere MT-RNA. Æg høstes fra kvindelige X. Tropicalis frøer. Efter fremstilling af en cytoplasmisk ekstrakt, taxol tilføjes for at inducere mikrotubuli polymerisation. Mikrotubuli og MT-RNA oprenset ved sedimentering gennem en glycerolpude. (B) Coomassie gel analyse af isolerede proteiner ved anvendelse af ordningen beskrevet i (A). Samlet CSF ekstrakt i forhold til proteiner sedimenterede i nærvær af taxol eller nocodazol. (C) Bioanalyzer gel analyse af RNA isoleret bruger ordningen beskrevet i (A). RNA isoleret fra CSF ekstrakt i forhold til RNA aflejret i nærvær af taxol eller nocodazol. Både gel projektion og den linje spor vises. Genoptrykt med tilladelse fra Sharp, et al., (2011). Klik her for at se større billede .
Discussion
I denne rapport har vi beskrevet en enkel metode til at producere CSF-anholdte æg uddrag X. tropicalis 19 og bruge dette ekstrakt til at studere mikrotubuli-associerede RNA 7.. Den grundlæggende procedure for at producere CSF-anholdt æg uddrag fra X. tropicalis er den samme som anvendt for X. laevis med et par vigtige forskelle. Et af de mest udfordrende aspekter at arbejde med X. tropicalis frøer er at opnå tilstrækkelig høj kvalitet æg til at gøre en ekstrakt med mikrotubuli nukleering eller spindel samling aktivitet sammenlignelig med X. laevis æg ekstrakter. For at opnå optimale æglæggende betingelser samtidig forhindre afvigelser fra meiose II cellecyklusstop, bliver intervallet mellem hormonindsprøjtninger for X. tropicalis er kortere end den, der anvendes for X. laevis, og timingen af den tredje hCG injektion til begyndelsen af æglægningen er også meget kortere. Med X. laevis timingen fra hCG injektionen til than begyndelsen af æglægningen er sådan, at det er praktisk og effektivt for æg, der skal fastsættes natten ind i buffer. Men på grund af den kortere tid mellem hCG injektion og æglægning med X. Tropicalis det er ofte nødvendigt manuelt udtrykke æg fra frøer. En anden væsentlig forskel mellem at gøre æg uddrag af to forskellige frøer er dejellying skridt. Med X. laevis æggene er så store, at det er let at bestemme, hvornår jelly lag er opløst ved at observere, hvor tæt æggene er fordelt i bægerglasset. Som dejellying reaktion påbegyndes, æggene begynder at pakke mere tæt. Men X. tropicalis æg er meget mindre, og det kan være ganske svært at afgøre, hvornår jelly lag er opløst ved ægpakkeri tæthed alene. Vi har fundet, at den mest pålidelige metode til at bestemme, hvornår gelé lag er opløst, er at overvåge orienteringen af dyr (sort) og vegetabilsk (hvid) poler. Når alle vegetative poler orient mod bunden af bægerglasset jelly lag er blevet fjernet nok til at fortsætte med ekstraktet. Endelig mens X. laevis æggeekstrakt kan opbevares ved kølige temperaturer (4-12 ° C) har vi observeret, at det er afgørende at fastholde X. tropicalis æggeekstrakt ved stuetemperatur (20-25 ° C) under forberedelse og eksperimentelle manipulationer for at bevare biokemisk aktivitet. På grund af forskellene i brugervenlighed, vi foretrækker at bruge X. laevis frøer til produktion af æg ekstrakt. Men for eksperimenter, der kræver eller lettes af en organisme med en genom, X. tropicalis er et glimrende alternativ system.
Den metode, som vi har beskrevet i denne rapport anvendt taxol som en mikrotubulus-stabiliserende middel til at inducere mikrotubuli polymerisation. Vi valgte denne metode, fordi taxol er en robust mikrotubulus-stabiliserende middel, der letter storstilet isolering af oprensede mikrotubuli. Fremgangsmåden thved vi beskrev kunne sandsynligvis forbedres ved at sammenligne proteiner og RNA'er associeret med mikrotubuli anvender alternative mikrotubuli polymerisationsmetoder. Alternativer kunne omfatte polymerisation under anvendelse GTP-induceret polymerisation (en klassisk teknik), 22 eller ved hjælp af Ran-GTP som en mikrotubulus polymerizer at efterligne mikrotubuli induceret af kromatin-drevet spindel samling 23. Endelig anvendelse af oprenset sæd kerner at inducere mikrotubuli polymerisation ville være det tætteste efterligner på de typer af mikrotubuli, der er kerneholdige under mitosen (centrosome, chromatin og kinetochore medieret). Ulemper til disse alternative kilder til mikrotubuli nukleering er, at de nukleeringsmidler ikke er så let tilgængelig som taxol og de ikke nucleate eller stabilisere mikrotubuli så effektivt som taxol. Derfor ville hver af disse metoder være sværere at bruge til oprensninger i stor skala. Fordelen ved at sammenligne flere forskellige typer af mikrotubuli kimdannelsesmidlerer, at det kunne være muligt at identificere proteiner og / eller RNA'er, der er specifikke for hver vej af mikrotubuli nukleering.
Den metode, som vi har beskrevet her drager fordel af cytoplasmatiske ekstrakter af padder. Imidlertid kan denne fremgangsmåde udvides til brugen af ekstraktet system fra andre organismer. Mitotiske uddrag er beskrevet fra synkroniserede humane vævskulturceller 24 at trofast rekapitulere mange aspekter af mikrotubuli. Vi har med succes brugt disse ekstrakter til at identificere mikrotubuli-associerede RNA fra HeLa-celler 5. Lignende mikrotubuli oprensningsskemaer er blevet beskrevet til mange forskellige organismer 25,26, selvom mikrotubuli forbundet RNA'er ikke er blevet undersøgt. Den fremgangsmåde der er beskrevet her, kan anvendes sammen med enhver organisme, der kan producere en koncentreret cytoplasmatisk ekstrakt kan kimdannende mikrotubuli.
Endelig, selv om den tilgang, at vi deskriftklog her diskuterer rensning af mikrotubuli og tilhørende proteiner og RNA'er, kan denne fremgangsmåde være generaliseres til andre subcellulære strukturer. Mens de fleste lokaliserede mRNA ikke er blevet identificeret ved hjælp af biokemiske metoder de seneste fremskridt i DNA-og RNA-sekventering teknologier gør denne tilgang til en attraktiv metode til at identificere lokaliserede RNA. I denne tilgang enhver subcellulært eller sub-embryo struktur af renter kan isoleres eller oprenses. Så de tilknyttede proteiner og RNA'er kan identificeres på et genom bred skala. RNA'er kan derefter sammenlignes til RNA indholdet af den samlede celle eller embryo at identificere beriget lokaliserede RNA'er. Denne fremgangsmåde kan bruges med hele æg (animalsk og vegetabilsk separation, svarende til den tilgang, identificerede de første lokaliserede RNA i Xenopus 27), actin tilknyttede RNA'er, ER-associerede RNA'er, mitokondrier-associerede RNA'er, eller nogen subcellulære struktur, der kan oprenses med tilhørende RNA'er intakte. Baseret påvores arbejde med mikrotubuli-associeret RNA forudser vi, at dette ville være en fremragende metode til at opdage nye proteiner, der fungerer på en given lokalitet. Desuden vil identifikation af placeringen og omfanget af alle lokaliserede RNA give indsigt i, hvordan celler og embryoer bruge mRNA lokalisering til at styre genekspression.
Disclosures
Ingen interessekonflikter erklæret.
Materials
| Name | Company | Catalog Number | Comments |
| REAGENTS | |||
| Xenopus tropicalis | NASCO | LM00823MX | |
| human Chorionic Gonadotropin | Sigma-Aldrich | CG10 | |
| HEPES | Sigma-Aldrich | H4034 | |
| EDTA | Sigma-Aldrich | E5134 | |
| NaCl | Sigma-Aldrich | S3014 | |
| KCl | Sigma-Aldrich | P9541 | |
| MgCl2 | Sigma-Aldrich | M8266 | |
| CaCl2 | Sigma-Aldrich | C8106 | |
| sucrose | Sigma-Aldrich | S0389 | |
| NaOH | Sigma-Aldrich | S5881 | |
| EGTA | Sigma-Aldrich | E3889 | |
| Leupeptin | Sigma-Aldrich | L9783 | |
| Pepstatin | Sigma-Aldrich | P5318 | |
| Chymostatin | Sigma-Aldrich | C7268 | |
| Cytochalasin D | Sigma-Aldrich | C8273 | |
| Gelatin, porcine skin | Sigma-Aldrich | G1890 | |
| PIPES | Sigma-Aldrich | P6757 | |
| Taxol | Sigma-Aldrich | T7191 | |
| Nocodazole | Sigma-Aldrich | M1404 | |
| Trizol | Invitrogen | 15596-026 | |
| L-Cysteine, free base | USB Corporation | 14030 | |
| Cichlid Lake Salt | Seachem | 47894 | |
| Marine salt | Seachem | SC7111 | |
| NaHCO3 | Sigma-Aldrich | S6014 | |
| EQUIPMENT | |||
| 1 ml syringes | BD Biosciences | 309659 | |
| 18 gauge needles | BD Biosciences | 305195 | |
| 30 gauge needles | BD Biosciences | 305106 | |
| Rubbermaid Plastic bucket | Amazon | 6306 | |
| Beckman Polyallomer 2 x ½ inch Ultracentrifuge tubes | Beckman | 326819 | |
| 15 ml round-bottomed glass centrifuge tubes | Fisher Scientific | 45500-15 | |
| Rubber adapter sleeves for 15 ml tubes | Kimble-Chase | 45550-15 | |
| 5 ¾ inch glass Pasteur pipettes | Fisher Scientific | 13-678-20A | |
| 14 ml polypropylene round-bottom tube | BD Biosciences | 352059 | |
| Sorvall HB-6 rotor | Thermo Scientific | 11860 | |
| Sorvall RC-6 centrifuge | Thermo Scientific | 46910 | |
References
- Martin, K. C., Ephrussi, A. mRNA localization: gene expression in the spatial dimension. Cell. 136, 719-730 (2009).
- Besse, F., Ephrussi, A. Translational control of localized mRNAs: restricting protein synthesis in space and time. Nat. Rev. Mol. Cell Biol. 9, 971-980 (2008).
- Lawrence, J. B., Singer, R. H. Intracellular localization of messenger RNAs for cytoskeletal proteins. Cell. 45, 407-415 (1986).
- Mingle, L. A., et al. Localization of all seven messenger RNAs for the actin-polymerization nucleator Arp2/3 complex in the protrusions of fibroblasts. J. Cell Sci. 118, 2425-2433 (2005).
- Blower, M. D., Feric, E., Weis, K., Heald, R. Genome-wide analysis demonstrates conserved localization of messenger RNAs to mitotic microtubules. J. Cell Biol. 179, 1365-1373 (2007).
- Eliscovich, C., Peset, I., Vernos, I., Mendez, R. Spindle-localized CPE-mediated translation controls meiotic chromosome segregation. Nat. Cell Biol. 10, 858-865 (2008).
- Sharp, J. A., Plant, J. J., Ohsumi, T. K., Borowsky, M., Blower, M. D.
- Lecuyer, E., et al. Global analysis of mRNA localization reveals a prominent role in organizing cellular architecture and function. Cell. 131, 174-187 (2007).
- Mili, S., Moissoglu, K., Macara, I. G. Genome-wide screen reveals APC-associated RNAs enriched in cell protrusions. Nature. 453, 115-119 (2008).
- Holt, C. E., Bullock, S. L. Subcellular mRNA localization in animal cells and why it matters. Science. 326, 1212-1216 (2009).
- Desai, A., Murray, A., Mitchison, T. J., Walczak, C. E. The use of Xenopus egg extracts to study mitotic spindle assembly and function in vitro. Methods Cell Biol. 61, 385-412 (1999).
- Gache, V., Waridel, P., Luche, S., Shevchenko, A., Popov, A. V. Purification and mass spectrometry identification of microtubule-binding proteins from Xenopus egg extracts. Methods Mol. Med. 137, 29-43 (2007).
- Gache, V., et al.
- Cross, M. K., Powers, M. Obtaining eggs from Xenopus laevis females. J. Vis. Exp. (18), e890 (2008).
- Cross, M. K., Powers, M. Preparation and fractionation of Xenopus laevis egg extracts. J. Vis. Exp. (18), e891 (2008).
- Cross, M., Powers, M. In vitro nuclear assembly using fractionated Xenopus egg extracts. J. Vis. Exp. (19), e908 (2008).
- Harland, R. M., Grainger, R. M. Xenopus research: metamorphosed by genetics and genomics. Trends Genet. 27, 507-515 (2011).
- Hellsten, U. The genome of the Western clawed frog Xenopus tropicalis. Science. 328, 633-636 (2010).
- Brown, K. S. Xenopus tropicalis egg extracts provide insight into scaling of the mitotic spindle. J. Cell Biol. 176, 765-770 (2007).
- Godfrey, E. W., Sanders, G. E. Effect of water hardness on oocyte quality and embryo development in the African clawed frog (Xenopus laevis). Comp. Med. 54, 170-175 (2004).
- Groisman, I., et al. CPEB, maskin, and cyclin B1 mRNA at the mitotic apparatus: implications for local translational control of cell division. Cell. 103, 435-447 (2000).
- Budde, P. P., Desai, A., Heald, R. Analysis of microtubule polymerization in vitro and during the cell cycle in Xenopus egg extracts. Methods. 38, 29-34 (2006).
- Kalab, P., Pu, R. T., Dasso, M. The ran GTPase regulates mitotic spindle assembly. Curr. Biol. 9, 481-484 (1999).
- Gaglio, T., Saredi, A., Compton, D. A. NuMA is required for the organization of microtubules into aster-like mitotic arrays. J. Cell Biol. 131, 693-708 (1995).
- Hughes, J. R., et al. A microtubule interactome: complexes with roles in cell cycle and mitosis. PLoS Biol. 6, e98 (2008).
- Suprenant, K. A., Tempero, L. B., Hammer, L. E. Association of ribosomes with in vitro assembled microtubules. Cell Motil. Cytoskeleton. 14, 401-415 (1002).
- Rebagliati, M. R., Weeks, D. L., Harvey, R. P., Melton, D. A. Identification and cloning of localized maternal RNAs from Xenopus eggs. Cell. 42, 769-777 (1985).

{kind=link}