

Summary
Vi beskriver insamling av obefruktade
Abstract
Många organismer lokalisera mRNA till specifika subcellulära destinationer att rumsligt och tidsmässigt kontrollerar genuttryck. Nyligen genomförda studier har visat att majoriteten av transkriptom är lokaliserad till ett icke-slumpvis position i celler och embryon. En metod för att identifiera lokala mRNA är att biokemiskt rena en cellstruktur av intresse och för att identifiera alla tillhörande transkript. Använda nyutvecklade hög genomströmning sekvensering teknik är det nu enkelt att identifiera alla RNA associerade med en subcellulär struktur. För att underlätta utskrift identifiering är det nödvändigt att arbeta med en organism med ett fullt sekvenserat genomet. Ett attraktivt system för biokemisk rening av subcellulära strukturer är ägg extrakt framställda från grodan Xenopus laevis. Men X. laevis närvarande inte har ett fullt sekvenserat genomet, vilket försvårar avskrift identifiering. I den här artikeln beskriver vi en metodatt producera ägg utdrag från en närstående groda, X. tropicalis, som har ett fullt sekvenserade genom. Vi tillhandahåller information för mikrotubuli polymerisering, rening och avskrift isolering. Även här artikeln beskriver en specifik metod för identifiering av mikrotubuli-associerade transkript, tror vi att det lätt kommer att tillämpas på andra subcellulära strukturer och kommer att ge en kraftfull metod för identifiering av lokaliserade RNA.
Introduction
Rumslig och tidsmässig kontroll av genuttryck är viktigt för alla celler, och är särskilt viktigt för kontroll av tidiga embryonala smattrande 1. Rumslig kontroll av genuttryck uppnås genom aktiv lokalisering av mRNA till specifika destinationer inom celler eller embryon. I många mycket stora celltyper, (t.ex. ägg, embryon och neuroner) mRNA lokalisering används för att begränsa protein expression till platsen för verkan av det kodade proteinet. Eftersom en lokaliserad mRNA kan katalysera många rundor av proteinproduktion det är mer effektivt att lokalisera ett mRNA än att lokalisera enskilda proteinmolekyler. Lokaliserade mRNA typiskt translatoriskt undertrycks tills de når sin destination, som tjänar till att ytterligare begränsa lokaliseringen av det kodade proteinet 2. Förutom de många väldokumenterade fall av RNA-lokalisering för att styra embryonala mönstring har flera studier dokumenterade mRNA som är lokaliseradetill platsen för verkan av det kodade proteinet. Framstående exempel lokalisering av β-aktin 3 och Arp2 / 3 4 mRNA till den främre kanten av motila fibroblaster och lokalisering av mRNA för många mitotiska regulatorer till meiotiska och mitotiska spolar 5-7.
Många av de klassiska exemplen på lokala mRNA identifierades genom genetiska skärmar för maternal effekt mutationer och senare bestämdes att koda lokaliserade RNA. Men senare analyser över hela arvsmassan börjat ge bredare inblick i omfattningen av lokaliserade RNA. En nyligen in situ hybridisering skärm i Drosophila embryon visade att ~ 70% av alla mRNA har en specifik lokalisering, inklusive många nya destinationer 8. Rening av pseudopodia från musfibroblaster identifierat en mångskiftande grupp av lokaliserad mRNA 9. Arbete från vår grupp med biokemisk rening av mikrotubuli från meiotisk Xenopus ägg extraherar identifierade hundratals mRNA som samrena med spindeln 5,7. Vårt arbete visar att majoriteten av mikrotubuli-lokaliserade mRNA kodar för proteiner som fungerar i kontrollen av mitos, stöder idén att mRNA lokaliserad till platsen för verkan av det kodade proteinet. Vidare belyser förmågan att detektera mRNA anrikning i en subcellulär fraktion genom biokemisk rening kraften i denna metod för identifiering av lokaliserade mRNA.
De flesta lokaliserade RNA använder aktiv transport på cytoskeletonen, antingen aktin eller mikrotubuli, för att uppnå transport till slutdestinationen 10. För att få en bättre förståelse av omfattningen och typen av RNA som är lokaliserade till specifika platser med en biokemisk metod är det nödvändigt att ha ett in vitro-system som kan rekapitulera cytoskelettala processer. En av de främsta systemen för att studera cytoskeletal biologi är ägg producerade extraktfrån obefruktade ägg från grodan Xenopus laevis. X. laevis ägg extrakt har använts i årtionden för att studera ett brett spektrum av cytoskelett processer och har bidragit mycket till vår förståelse av de mekanismer och molekyler som styr cytoskeletal montering och dynamik 11. Vidare X. laevis ägg extrakt är mottagliga för storskaliga reningar av mikrotubuli och associerade proteiner 12,13 och det finns väl utformade metoder för produktion av olika typer av ägg extrakt 14-16. Men för genomic studier finns det flera nackdelar med användningen av X. laevis som modellsystem.
För årtionden Xenopus laevis grodor har varit ett kraftfullt system för studier av utvecklings-och cellbiologi, på grund av den stora äggcellen storlek och robust extern utveckling 17. Vidare utveckling av äggextrakt system som kan rekapitulera många cellulära processeer i ett provrör har gjort denna groda en kraftfull experimentell modell. Dock har Xenopus laevis hämmats av bristen på en fullständig arvsmassa, som har saktat av allotetraploid natur genome.In Däremot har en närbesläktad art, Xenopus tropicalis, en diploid arvsmassa som sekvenserades under 2010 18. Medan X. tropicalis är inte så experimentellt lätthanterligt som X. laevis 17 tillgängligheten av ett sekvenserat genomet gör det till en attraktiv modell för att utföra hela genom analyser.
I denna rapport beskriver vi en metod för att göra meios II-, cytostatika faktor-gripna extrakt (CSF) från X. tropicalis 19. Vi beskriver sedan en enkel metod för att rena mikrotubuli och tillhörande RNA från detta extrakt. De RNA kan sedan omvandlas till biblioteken lämpar sig för sekvensering med Nyutvecklad teknik genomströmning sekvensering. När bibliotekensekvenseras de kan anpassas till genomet av grodan att identifiera specifika mRNA som anrikas i mikrotubuli provet jämfört med total extrakt. Detta ger en kraftfull metod för att upptäcka mikrotubuli-riktad mRNA lokalisering på en genomet hela skalan. Förutom att kunna upptäcka lokaliserade mRNA, användning av hög genomströmning sekvensering och ett sekvenserat genomet erbjuder möjligheten att upptäcka nya transkript som inte är närvarande i offentlig databas anteckningar.
Protocol
Ett. Generation X. tropicalis Ägg
Alla Xenopus tropicalis grodor beställs från NASCO. Våra grodor är inrymt i ett akvatiska livsmiljöer recirkulerande vattensystem hålls vid 27 ° C. Det finns många alternativ för vattenanläggningar för vård av X. tropicalis. Några bra allmän information om denna grodarter kan hittas på webbplatserna för Harland och Grainger Labs ( http://tropicalis.berkeley.edu/home/ , http://www.faculty.virginia.edu/xtropicalis / ). Våra grodor bibehålls i tankwater bestående av (0,4 g Ciclid Lake Salter, 0,6 g marin salt, 0,625 g NaHCO 3 per liter vatten, pH 7,0) 20. Detta recept resulterar i en konduktivitet på ~ 1800 | iS, vilket är en hög salthalt för X. tropicalis. Däremot har vi funnit att våra grodor trivs i this miljö-och äggcell kvalitet förbättras. Alternativa tankwater recept kan hittas ovan på de resurser som anges för allmän X. tropicalis vård.
- Grodor injiceras med humant koriongonadotropin (hCG) på tre på varandra följande dagar för att stimulera äggläggningen: Först förbereder två koncentrationer av hCG lösning. Återsuspendera 10000 U av lyofiliserat hCG pulver i 10 ml sterilt, avjoniserat H2O för en slutlig koncentration av 1000 E / ml. Sedan, späd 1 ml 1000 U / ml hCG lösning i 9 ml H2O för en slutkoncentration av 100 E / ml. Förvara båda lösningarna vid 4 ° C.
- På dag 1, förbereda 4-6 grodor för äggläggning genom att injicera med hCG mellan 2:00 till 15:00. Injicera varje groda i den dorsala lymfan sac nära cloacaen med 0,2 ml 100 E / ml hCG lösning. Med grodorna snabbt under de följande två injektioner kommer att minimera mängden grodan avfall närvarande under äggläggningen, men är valfritt.
- På dag 2, injicera samma grodormed 0,2 ml 100 U / ml hCG lösning mellan 2:00 till 15:00.
- På dag 3, injicera samma grodorna med 0,2 ml 1,000 U / ml hCG lösning, mellan 7:00 till 10:00. Ställ upp grodor för att lägga ägg: fylla en 6-liter plasthink med färska tankwater, lägger grodor och placera i mörker vid 25 ° C. Efter denna injektion, kommer äggläggningen påbörjas efter 4 timmar och kommer att vara klar med 7 tim. Grodor ska lägga ägg i en miljö som hålls på ett minimum av 25 ° C.
- Gör extrahera lösningar och har utrustning redo omedelbart före samla ägg.
20X MMR: 100 mM HEPES, pH 7,8, 2 mM EDTA pH 7,8, 2 M NaCl, 40 mM KCl, 20 mM MgCl2, 40 mM CaCl2. Autoklav och förvara i rumstemperatur. Bered en L 1X MMR just före extrahera beredning.
10X XB: 100 mM HEPES, pH7.7, 10 mM MgCl2, 1 mM CaCl2, 1 M KCl, 500 mM sackaros. Autoklav och förvara vid 4 ° C. Bered en L 1X XB just innan extrahera beredning. Dejelly lösning: Bered 250 ml 3% cystein lösning i avjoniserat H2O och pH till 7,8 till 8,0 med 10 N NaOH. Bered omedelbart före extrahera beredning.
CSF-XB: ta 200 ml 1X XB och tillsätt 2 ml 0,5 M EGTA pH 7,7 och 200 ^ 1 M MgCl2. Bered omedelbart före extrahera beredning.
CSF-XB +: ta 50 ml av CSF-XB och tillsätt 50 pl av LPC (10 mg / ml vardera lager av Leupeptin, Pepstatin, och kymostatin i DMSO). Tillsätt 50 l Cytochalasin D (10 mg / ml i DMSO). Bered omedelbart före extrahera beredning.
Bered en 0,2% gelatinlösning i avjoniserat H2O, mikrovågsugn för att lösa upp och filtersterilisera. Förvara vid rumstemperatur.
Reserv 2 Beckman 2 x ½ tum ultracentrifugrör.
Bered två 15 ml glas rundbottnade centrifugrör med 0,5 ml H2O i varje för att dämpaultracentrifug röret.
Gör brand-polerat glas Pasteur pipetter. Snap slutet av av 5 ¾ tums glaspipetter att exponera en bred öppning, och utsättas för lågor för att jämna ut nya exponerade pipettspetsen.
- Förbered en 500 ml glasbägare för lagring av ägg genom att snurra en 0,2% gelatin lösning kring att belägga väggarna i bägaren. Kasta gelatin lösning från bägaren efter användning.
- Samla ägg från plasthink som används för att lägga 6-7 timmar efter den tredje injektionen på dag 3. Om så önskas, tryck försiktigt varje grodan en gång för att få eventuella kvarvarande ägg. Tvätta ägg en gång med färsk tankwater och överför det till 500 ml glasbägare belagd med 0,2% gelatin lösning.
2. Beredning av extrakt från X. tropicalis Ägg
Alla moment i extrakt preparatet kan utföras vid rumstemperatur, ca 25 ° C. Alltigenom tvättar, är det viktigt att hålla äggets nedsänkt under flytande så att de förblir våta. Exponering för luft kan orsaka äggen att undkomma arrestering cellcykeln eller lysera.
- Dekantera så mycket tankwater som möjligt samtidigt reservera tillräckligt med vätska för att hålla äggen våt. Luta bägare innehållande ägg åt sidan och tillsätt ~ 300 ml 1X MMR långsamt till väggen i bägaren, så att fysisk agitation av äggen minimeras. Låt äggen lösa, sedan dekantera av supernatanten innehållande skräp. X. tropicalis ägg är trådiga i detta steg, så avlägsnande av aktiverade ägg sker efter dejellying. Upprepa för totalt tre 1X MMR tvättar.
- Dejelly äggen. Dekantera av så mycket MMR som möjligt och tillsätt hälften av dejelly lösningen. Snurra kontinuerligt under cirka 5 minuter. Dissolving gelé rockar kommer att synas i supernatanten efter ett par minuter. Dekantera av och tillsätt resterande dejelly lösningen. Fortsätt att snurra kontinuerligt tills äggen packa mycket tätt och alla Orienten med sin vegetativa polen (stångenmed vitt pigment) mot botten av skålen. Snabbt dekantera av så mycket dejelly lösning som möjligt. När äggen är dejellied de är mycket känsliga för mekaniska manipulationer.
- Tillsätt försiktigt XB till äggen. I den första XB tvätten, ta bort ägg som har undgått CSF gripandet genom att ta bort lyserade, pösigt, vit, och pseudocleavage ägg. Aktiverade X. tropicalis ägg tenderar att bosätta sig i övre mitten, så använd en plast överföring pipett för att dra ut dessa. Också ta bort bitar av hud och grodan avfall. Tvätta ägg totalt tre gånger med ~ 300 ml 1X XB lösning, försiktigt virvlande ägg mellan tvättar och låta dem bosätta sig på botten av bägaren. Liksom tidigare dekantera så mycket av varje tvättlösning som möjligt och samtidigt hålla ägg våt.
- Tvätta ägg två gånger med CSF-XB och dekantera.
- Lägg CSF-XB + ägg. Använda en gelatin-behandlad avbränd Pasteur pipett ägg till Ultra-centrifugrör med CSF-XB +, noga med att inte utsätta eGGS till luft. Rum inne i 15 ml rör av glas centrifugrör med vattnet dynan.
- Snurra ägg i en klinisk centrifug vid 200 xg under 1 min, öka hastigheten till 800 xg och centrifugera i 30 sek.
- Använd en aspirator för att avlägsna så mycket buffert som möjligt från ägg. De bör vara nästan torr på toppen. Snabbt flytta ägg till en Sorvall RC-6-centrifug utrustad med en HB-6 rötor (eller motsvarande) och snurra 17.000 xg under 15 minuter vid 20 ° C.
- Ta bort den gula cytoplasmatiska skikt mellan pigment och skikten lipid med användning av en 18 gauge nål fäst till en 1 ml spruta. Punktering den sida av röret och dra sprutcylindern långsamt för att erhålla cytoplasmatiskt extrakt skiktet. Undvik pigmentkorn så mycket som möjligt.
- Överför cytoplasman till nya ultracentrifugering röret. Det är normalt att extraktet att synas en del moln vid detta steg. Placera inuti 15 ml glasflaska centrifugrör med vatten dyna. Snurra igen 17.000 xg under 10 minuter vid 20 ° C. Upprepa extdiffraktion med 18 gauge nål.
- Överför cytoplasman till ett 1,5 ml mikrofugrör. Uppskatta extrakt volym och späd Cytochalasin D och LPC 1:1000 i extraktet. Blanda väl med en 1 ml pipettspets, pipettera upp och ned flera gånger utan införande av luftbubblor. Ett typiskt utbyte från en frisk groda koloni är cirka 300-500 il extrakt / frog. För att bevara maximal aktivitet, är det nödvändigt att lagra i och utför experimentella manipulationer vid rumstemperatur (20-25 ° C).
Tre. Reningsanläggningar Taxol-stabiliserade mikrotubuli från X. tropicalis Extract
- Lägg Taxol till en 100 till 200 | il alikvot av extraktet vid en slutlig koncentration av 10 | iM och inkubera vid rumstemperatur i 30 min. För kontroll reaktioner, behandla en motsvarande volym av extraktet med mikrotubuli-destablilzing drogen Nocodazole (10 M). Reserv 100 | il obehandlat extrakt för analys.
- Späd läkemedelsbehandladeextrahera med 10 volymer BRB-80 (80 mM PIPES pH 6,8, 1 mM MgCl2, 1 mM EGTA) + 30% glycerol. Montera 14 ml rundbottnad polypropenrör innehållande 10 ml av BRB-80 + 60% glycerol kudde. Med hjälp av en bred borrning pipettspets, skikt den läkemedelsbehandlade extrakt reaktionen försiktigt på toppen av BRB-80 + 60% glycerol kudde. Centrifugera i 10 minuter vid 17.000 xg vid 20 ° C i en Sorvall RC-6-centrifug utrustad med en HB-6 rötor (eller motsvarande) och adaptrar tube.
- Aspirera supernatanten innehållande osedimenterade extraktmaterial och tvätta gränssnittet två gånger med avjoniserat H2O Aspirera återstående kudden volymen långsamt, noga med att inte störa gel-liknande pellets innehållande mikrotubuli, mikrotubuli-associerade proteiner och mikrotubuli-associerade RNA i taxolbehandlade prov. Den Nocodazole-behandlade provet inte innehåller synliga materialet. Suspendera pelleten i 1 ml TRIzol och fortsätt med tillverkarens anvisningar för att isolera RNA. Untreated extrakt (upp till 100 | il) kan återsuspenderas direkt i 1 ml TRIzol.
- Det finns nu kommersiellt tillgängliga kit för framställning av transkriptom bibliotek som är lämpliga för RNA-seq. Dessa kan köpas genom http://www.illumina.com/ och http://www.454.com/ .
Representative Results
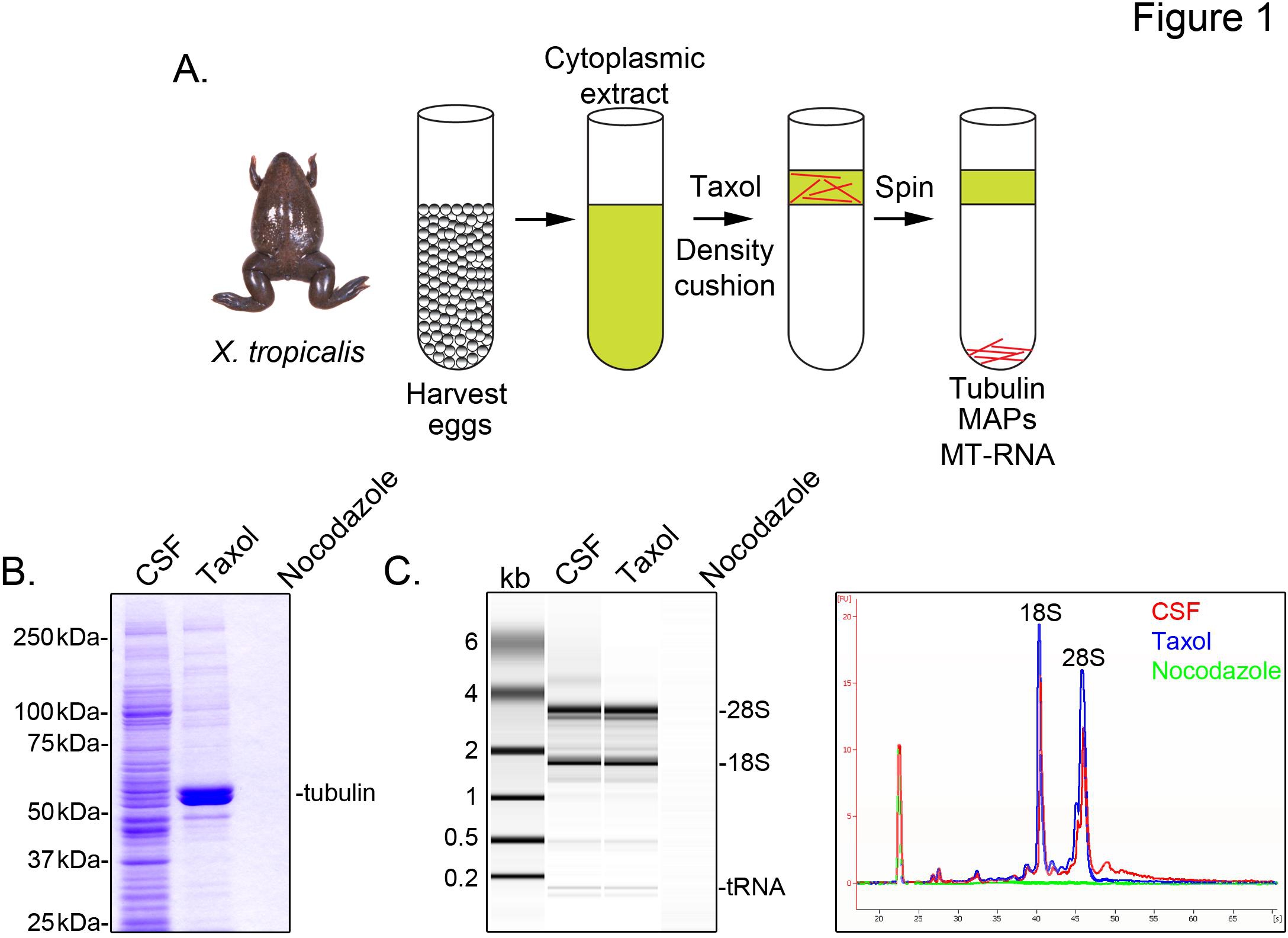
Att identifiera X. tropicalis transkript samband med mikrotubuli, förbereder vi en cytosoliskt extrakt från obefruktade ägg greps i metafas av meios II (CSF). Behandling av detta extrakt med taxol medger bildning av stabila mikrotubuli som kan renas genom sedimentering genom en glycerol kudde (Figur 1A). Coomassie gel analys bekräftar att α / β-tubulin sediment i en taxol-beroende sätt, och utgör den största proteinspecies återvinns i dessa preparat (Figur 1B). Lägre nivåer av andra proteiner är också närvarande i den taxol pelleten, men inte i beredningar behandlade med mikrotubuli depolymeriserande läkemedlet nokodazol, vilket indikerar att proteiner i taxol fraktionen specifikt associerar med mikrotubuli (MAP).
En Agilent Bioanalyzer används för att undersöka allmänna RNA sammansättning i alla X. tropicalis extrakt fraktioner (Figur 1C X. laevis äggextrakt 5,21. En rad spår av gelén projektionen avslöjar mRNA-signalen är betydligt lägre i mikrotubuli-innehållande Taxol pellets, framför allt i området migrerar ovanför 28S rRNA, vilket indikerar att en delmängd av mRNA cosediment med mikrotubuli i X. tropicalis. RNA isolerat på detta sätt är lämplig för RNA-Seq experiment med användning av kommersiellt tillgängliga reagens.

Figur 1. Rening av MT-RNA för RNA-punkter. (A) reningsschema för att isolera MT-RNA. Ägg skördas från kvinnliga X. Tropicalis grodor. Efter beredning av ett cytoplasmatiskt extrakt, är Taxol för att inducera mikrotubuli polymerisering. Microtubules och MT-RNA renas genom sedimentation genom en glycerol kudde. (B) Coomassie-gelanalys av isolerade proteiner som använder systemet som beskrivs i (A). Totalt CSF-extrakt jämfört med proteiner sedimenterade i närvaro av taxol eller nokodazol. (C) Bioanalyzer gelanalys av RNA isolerades med användning av det system som beskrivits i (A). RNA isolerat från CSF extrakt jämfört med RNA sedimenterades i närvaro av taxol eller nokodazol. Både gelen projektion och spåren linje visas. Omtryckt med tillåtelse från Sharp, et al., (2011). Klicka här för att se större bild .
Discussion
I denna rapport har vi beskrivit en enkel metod för att producera CSF-greps ägg extrakt från X. tropicalis 19 och använder detta extrakt för att studera mikrotubuli-associerade RNA 7. Den grundläggande proceduren för att producera CSF-greps ägg extrakt från X. tropicalis är samma som användes för X. laevis med några viktiga skillnader. En av de mest utmanande aspekterna att arbeta med X. tropicalis grodor är att få tillräckligt hög kvalitet ägg för att göra ett utdrag med mikrotubuli nucleation eller spindelenhet aktivitet jämförbar med X. laevis ägg extrakt. För att uppnå optimala äggläggning förhållanden samtidigt förhindra glidning från meios II cellcykelstopp, intervallet mellan hormoneinjections för X. tropicalis är kortare än den som används för X. laevis, och tidpunkten från tredje hCG injektionen till början av äggläggningen är också mycket kortare. Med X. laevis timingen från hCG injektionen till than i början av äggläggningen är sådan att det är bekvämt och effektivt för ägg som skall fastställas natten i buffert. Men på grund av den kortare tid mellan hCG-injektionen och äggläggning med X. tropicalis är det ofta nödvändigt att manuellt uttrycker ägg från grodor. En annan viktig skillnad mellan att göra ägg utdrag ur två olika grodorna är dejellying steget. Med X. laevis äggen är så stora att det är lätt att avgöra när gelé pälsen har löst genom att observera hur nära äggen är åtskilda i bägaren. Som dejellying reaktionen påbörjas, äggen börjar att packa tätare. Emellertid X. tropicalis ägg är mycket mindre och det kan vara ganska svårt att avgöra när gelé pälsen har löst genom ägg packningsdensiteten ensam. Vi har funnit att den mest tillförlitliga metoden för att bestämma när gelé pälsen har lösts upp är att övervaka orienteringen av djuret (svart) och vegetabiliskt (vita) poler. När all den vegetativa poler orient mot botten av bägaren gelé pälsen har avlägsnats tillräckligt för att gå vidare med extraktet. Slutligen, medan X. laevis äggextrakt kan lagras vid svala temperaturer (4-12 ° C) har vi sett att det är viktigt att bibehålla X. tropicalis äggextrakt vid rumstemperatur (20-25 ° C) under beredning och experimentella manipulationer för att bevara biokemisk aktivitet. På grund av skillnaderna i användarvänlighet föredrar vi att använda X. laevis grodor för produktion av ägg extrakt. Men för experiment som kräver eller underlättas av en organism med ett sekvenserat genomet, X. tropicalis är ett utmärkt alternativt system.
Den metod som vi har beskrivit i denna rapport använder taxol som ett mikrotubuli-stabiliseringsmedel för att inducera mikrotubuli polymerisering. Vi valde denna metod eftersom taxol är en robust microtubule-stabiliseringsmedel som underlättar storskalig isolering av renade mikrotubuli. Metoden: eat vi beskrivit skulle sannolikt kunna förbättras genom att jämföra proteiner och RNA förknippade med mikrotubuli använder alternativa metoder mikrotubuli polymerisering. Alternativ kan innefatta polymerisation med GTP-inducerad polymerisation (en klassisk teknik), 22 eller använda Ran-GTP som mikrotubuli polymerizer att efterlikna mikrotubuli inducerade av kromatin-driven spolenheten 23. Slutligen använder renat spermier kärnor att inducera mikrotubuli polymerisation skulle vara närmast härma på de typer av mikrotubuli som kärnbildas under mitos (centrosomen, kromatin, och kinetochore medierad). Nackdelar med dessa alternativa källor av mikrotubuli kämbildning är att de kärnbildningsmedel är inte så lätt tillgänglig som taxol och de gör inte KÄRNA eller stabilisera mikrotubuli så effektivt som taxol. Därför skulle var och en av dessa metoder vara svårare att använda för storskaliga reningar. Fördelen med att jämföra flera olika typer av mikrotubuli nukleatorerär att det skulle vara möjligt att identifiera proteiner och / eller RNA som är specifika för varje bana av mikrotubuli nucleation.
Den metod som vi har beskrivit här utnyttjar cytoplasmaextrakt av amfibier. Emellertid kan denna metod utvidgas till användningen av extrakt system från andra organismer. Mitotiska extrakt har beskrivits från synkroniserade humana vävnadskulturceller 24 som troget rekapitulera många aspekter av mikrotubuli montering. Vi har med framgång använt dessa extrakt för att identifiera mikrotubuli-associerade RNA från HeLa-celler 5. Liknande mikrotubuli reningsscheman har beskrivits för många olika organismer 25,26, även om mikrotubuli associerade RNA inte har undersökts. Den metod som beskrivs här kan användas med vilken organism som helst som kan producera ett koncentrerat cytoplasmatiskt extrakt med förmåga att kärnbildande mikrotubuli.
Slutligen, även om det tillvägagångssätt som vi deskriftlärd här diskuterar rening av mikrotubuli och associerade proteiner och RNA, kan detta tillvägagångssätt vara generaliseras till andra subcellulära strukturer. Medan de flesta lokaliserade mRNA inte har identifierats med hjälp av biokemiska metoder de senaste framstegen inom DNA-och RNA-sekvensering teknik gör detta tillvägagångssätt en attraktiv metod för att identifiera lokala RNA. I denna strategi varje subcellulär eller sub-embryo strukturen av intresse kan isoleras eller renas. Då de associerade proteiner och RNA kan identifieras på ett genom bred skala. RNA kan sedan jämföras med RNA-halten av den totala cell eller embryo att identifiera anrikade lokaliserade RNA. Denna metod kan användas med hela ägg (animaliskt och vegetabiliskt separation, liknande den metod som identifierade den första lokaliserade RNA i Xenopus 27), aktin tillhörande RNA, ER-associerade RNA, mitokondrier-associerade RNA, eller till någon subcellulärt struktur som kan renas med tillhörande RNA intakta. Baserat påvårt arbete på mikrotubuli-associerade RNA vi förutspår att detta skulle vara en utmärkt metod för att upptäcka nya proteiner som fungerar på en viss plats. Dessutom kommer identifiering av platsen och omfattningen av alla lokaliserade RNA ge insikt i hur celler och embryon använda mRNA lokalisering att kontrollera genuttryck.
Disclosures
Inga intressekonflikter deklareras.
Materials
| Name | Company | Catalog Number | Comments |
| REAGENTS | |||
| Xenopus tropicalis | NASCO | LM00823MX | |
| human Chorionic Gonadotropin | Sigma-Aldrich | CG10 | |
| HEPES | Sigma-Aldrich | H4034 | |
| EDTA | Sigma-Aldrich | E5134 | |
| NaCl | Sigma-Aldrich | S3014 | |
| KCl | Sigma-Aldrich | P9541 | |
| MgCl2 | Sigma-Aldrich | M8266 | |
| CaCl2 | Sigma-Aldrich | C8106 | |
| sucrose | Sigma-Aldrich | S0389 | |
| NaOH | Sigma-Aldrich | S5881 | |
| EGTA | Sigma-Aldrich | E3889 | |
| Leupeptin | Sigma-Aldrich | L9783 | |
| Pepstatin | Sigma-Aldrich | P5318 | |
| Chymostatin | Sigma-Aldrich | C7268 | |
| Cytochalasin D | Sigma-Aldrich | C8273 | |
| Gelatin, porcine skin | Sigma-Aldrich | G1890 | |
| PIPES | Sigma-Aldrich | P6757 | |
| Taxol | Sigma-Aldrich | T7191 | |
| Nocodazole | Sigma-Aldrich | M1404 | |
| Trizol | Invitrogen | 15596-026 | |
| L-Cysteine, free base | USB Corporation | 14030 | |
| Cichlid Lake Salt | Seachem | 47894 | |
| Marine salt | Seachem | SC7111 | |
| NaHCO3 | Sigma-Aldrich | S6014 | |
| EQUIPMENT | |||
| 1 ml syringes | BD Biosciences | 309659 | |
| 18 gauge needles | BD Biosciences | 305195 | |
| 30 gauge needles | BD Biosciences | 305106 | |
| Rubbermaid Plastic bucket | Amazon | 6306 | |
| Beckman Polyallomer 2 x ½ inch Ultracentrifuge tubes | Beckman | 326819 | |
| 15 ml round-bottomed glass centrifuge tubes | Fisher Scientific | 45500-15 | |
| Rubber adapter sleeves for 15 ml tubes | Kimble-Chase | 45550-15 | |
| 5 ¾ inch glass Pasteur pipettes | Fisher Scientific | 13-678-20A | |
| 14 ml polypropylene round-bottom tube | BD Biosciences | 352059 | |
| Sorvall HB-6 rotor | Thermo Scientific | 11860 | |
| Sorvall RC-6 centrifuge | Thermo Scientific | 46910 | |
References
- Martin, K. C., Ephrussi, A. mRNA localization: gene expression in the spatial dimension. Cell. 136, 719-730 (2009).
- Besse, F., Ephrussi, A. Translational control of localized mRNAs: restricting protein synthesis in space and time. Nat. Rev. Mol. Cell Biol. 9, 971-980 (2008).
- Lawrence, J. B., Singer, R. H. Intracellular localization of messenger RNAs for cytoskeletal proteins. Cell. 45, 407-415 (1986).
- Mingle, L. A., et al. Localization of all seven messenger RNAs for the actin-polymerization nucleator Arp2/3 complex in the protrusions of fibroblasts. J. Cell Sci. 118, 2425-2433 (2005).
- Blower, M. D., Feric, E., Weis, K., Heald, R. Genome-wide analysis demonstrates conserved localization of messenger RNAs to mitotic microtubules. J. Cell Biol. 179, 1365-1373 (2007).
- Eliscovich, C., Peset, I., Vernos, I., Mendez, R. Spindle-localized CPE-mediated translation controls meiotic chromosome segregation. Nat. Cell Biol. 10, 858-865 (2008).
- Sharp, J. A., Plant, J. J., Ohsumi, T. K., Borowsky, M., Blower, M. D. Functional analysis of the microtubule-interacting transcriptome. Mol. Biol Cell. 22, 4312-4323 (2011).
- Lecuyer, E., et al. Global analysis of mRNA localization reveals a prominent role in organizing cellular architecture and function. Cell. 131, 174-187 (2007).
- Mili, S., Moissoglu, K., Macara, I. G. Genome-wide screen reveals APC-associated RNAs enriched in cell protrusions. Nature. 453, 115-119 (2008).
- Holt, C. E., Bullock, S. L. Subcellular mRNA localization in animal cells and why it matters. Science. 326, 1212-1216 (2009).
- Desai, A., Murray, A., Mitchison, T. J., Walczak, C. E. The use of Xenopus egg extracts to study mitotic spindle assembly and function in vitro. Methods Cell Biol. 61, 385-412 (1999).
- Gache, V., Waridel, P., Luche, S., Shevchenko, A., Popov, A. V. Purification and mass spectrometry identification of microtubule-binding proteins from Xenopus egg extracts. Methods Mol. Med. 137, 29-43 (2007).
- Gache, V., et al. Xenopus meiotic microtubule-associated interactome. PLoS One. 5, e9248 (2010).
- Cross, M. K., Powers, M. Obtaining eggs from Xenopus laevis females. J. Vis. Exp. (18), e890 (2008).
- Cross, M. K., Powers, M. Preparation and fractionation of Xenopus laevis egg extracts. J. Vis. Exp. (18), e891 (2008).
- Cross, M., Powers, M. In vitro nuclear assembly using fractionated Xenopus egg extracts. J. Vis. Exp. (19), e908 (2008).
- Harland, R. M., Grainger, R. M. Xenopus research: metamorphosed by genetics and genomics. Trends Genet. 27, 507-515 (2011).
- Hellsten, U. The genome of the Western clawed frog Xenopus tropicalis. Science. 328, 633-636 (2010).
- Brown, K. S. Xenopus tropicalis egg extracts provide insight into scaling of the mitotic spindle. J. Cell Biol. 176, 765-770 (2007).
- Godfrey, E. W., Sanders, G. E. Effect of water hardness on oocyte quality and embryo development in the African clawed frog (Xenopus laevis). Comp. Med. 54, 170-175 (2004).
- Groisman, I., et al. CPEB, maskin, and cyclin B1 mRNA at the mitotic apparatus: implications for local translational control of cell division. Cell. 103, 435-447 (2000).
- Budde, P. P., Desai, A., Heald, R. Analysis of microtubule polymerization in vitro and during the cell cycle in Xenopus egg extracts. Methods. 38, 29-34 (2006).
- Kalab, P., Pu, R. T., Dasso, M. The ran GTPase regulates mitotic spindle assembly. Curr. Biol. 9, 481-484 (1999).
- Gaglio, T., Saredi, A., Compton, D. A. NuMA is required for the organization of microtubules into aster-like mitotic arrays. J. Cell Biol. 131, 693-708 (1995).
- Hughes, J. R., et al. A microtubule interactome: complexes with roles in cell cycle and mitosis. PLoS Biol. 6, e98 (2008).
- Suprenant, K. A., Tempero, L. B., Hammer, L. E. Association of ribosomes with in vitro assembled microtubules. Cell Motil. Cytoskeleton. 14, 401-415 (1002).
- Rebagliati, M. R., Weeks, D. L., Harvey, R. P., Melton, D. A. Identification and cloning of localized maternal RNAs from Xenopus eggs. Cell. 42, 769-777 (1985).

{kind=link}