The EMT inducing culture conditions described here provide a robust method for the induction of EMT in a variety of cell types. Figures 1 and 2 demonstrate the morphology of and marker expression levels for 4 different human cell lines: T98G glioblastoma cells, HT29 colon adenocarcinoma cells, A549 lung carcinoma cells, and MCF10A mammary epithelial cells. Cells that were treated with the EMT Inducing Media Supplement changed from a classical epithelial morphology (Figures 1A – 1D) to a mesenchymal, spindle-shaped morphology (Figures 1E – 1H). EMT-induced cells appeared less densely packed into tight colonies compared to uninduced cells. Uninduced MCF10A samples contained tightly packed clusters surrounded by more loosely packed cells. These tightly packed clusters were E-cadherin positive (Figure 2D) and disappeared upon treatment with the EMT Inducing Media Supplement (Figure 2H).
EMT was also assessed by the downregulation of epithelial markers and upregulation of mesenchymal markers. The downregulation of E-cadherin expression is typically observed following EMT induction in different cell types3,4. Figure 2 demonstrates surface expression of E-cadherin in the majority of untreated cells (Figures 2A – 2D; red) in comparison to its absence after EMT induction (Figures 2E – 2H; red). One cell line, T98G, was found to have an extremely low basal level of E-cadherin prior to EMT induction, which precluded its analysis. Upregulation of the mesenchymal marker, Fibronectin, was also visible after induction with the EMT Inducing Media Supplement (Figures 2A – 2D versus Figures 2E – 2H; green).
The expression levels of E-cadherin and Fibronectin seen by immunocytochemistry were further confirmed through western blotting of total cell lysates (Figure 3). The western blot confirmed downregulation of E-cadherin and upregulation of Fibronectin expression seen by immunocytochemistry. The bands were quantitated using densitometry and normalized to GAPDH to determine relative fold change following EMT induction. E-cadherin levels were reduced 6.4 and 3.4 fold in A549 and MCF10A cells, respectively. Fibronectin levels were increased 4.6, 4.1, and 2.3 fold in T98G, A549, and MCF10A cells, respectively. Although the western blot does not show significant reduction of E-cadherin protein levels in HT29 cells post-EMT induction, the immunocytochemistry results demonstrate a reduction in surface expression of E-cadherin (Figure 2B versus 2F), which is unable to be distinguished in western blotting of total cell lysates.
To further evaluate EMT status, additional known markers for EMT were analyzed pre-and post-EMT induction. Consistent with previous results, E-cadherin was downregulated in all cell lines examined, indicating that EMT was induced. In addition, the mesenchymal markers, Vimentin and Snail, were upregulated in A549, T98G, and MCF10A cells following treatment with EMT Inducing Media Supplement (Figure 4). The HT29 cell line did not show expression of these mesenchymal markers either with or without EMT induction (data not shown), which may indicate that these cells utilize alternate pathways for EMT induction.
Although TGF-β is sufficient to induce EMT in certain cell contexts7,8,9, other cell types do not respond solely to the addition of TGF-β14. MCF7 human breast cancer cells and PANC-1 human pancreatic carcinoma cells are both reported not to respond to TGF-β signaling alone14. To determine whether both lines could undergo EMT in vitro, cells were cultured in the presence of the EMT Inducing Media Supplement. Cells were stimulated with recombinant TGF-β1 alone, at the concentration within the EMT Inducing Media Supplement. These cells retained their epithelial morphology, consisting of tightly packed colonies of cells (data not shown) and surface E-cadherin levels were similar to those seen in control cells (Figures 5C and 5D versus Figures 5A and 5B). In contrast, cells stimulated with the EMT Inducing Media Supplement showed a drastic decrease in surface E-cadherin levels (Figures 5E and 5F). These cells also obtained a more mesenchymal morphology, as cells became less compact and more spindle-like (data not shown). These results demonstrate that the EMT inducing method described here is able to promote EMT morphology and marker expression changes in cells typically resistant to TGF-β-induced EMT.
In addition to changes in marker expression, another hallmark of mesenchymal cells is their ability to migrate and invade. Migration and invasion of cells cultured with or without the EMT Inducing Media Supplement were analyzed using the 96 well BME Cell Invasion Assay according to the manufacturer’s instructions. Briefly, cells were seeded onto 96-well plates (50,000 cells/well) containing filter inserts with 8.0 μm pores. After 48 hr, migration was assessed by calcein AM staining of cells that had migrated through the filter. Each sample was tested in triplicate twice with similar results. Representative results from one experiment are shown in Figure 6A. Statistically significant increases (P-value<0.003) in cell migration were seen with both A549 and PANC-1 cells following EMT induction. To assess the ability of these cells to invade (i.e. migrate through extracellular matrix), the same assay was performed with a basement membrane extract-coated filter. EMT-induced cells showed a statistically significant (P- value<0.001) increase in invasion capabilities compared to untreated cells as seen in Figure 6B.
The robust induction of EMT is useful for the analysis of gene expression changes and signaling taking place in these cells. Antibody-based array analysis using lysates from both treated and untreated cells is one method to analyze the expression levels of a variety of molecules in a single sample. As an example, we analyzed the levels of phosphorylated MAPK family members in lysates from un-induced and EMT-induced MCF7 and A549 cells using the Proteome Profiler Human Phospho-MAPK Array Kit. The array was run twice from independent EMT-induction treatments with similar results. Figure 7 shows representative data from one array. Both cell types exhibited increased phosphorylation of CREB (S133), ERK1 (T202/Y204), and ERK2 (T185/Y187) in EMT-induced cells compared to controls. Differences in signaling events were observed between the two cell types as well. A549 cells showed an increase in GSK-3 β (S9) phosphorylation, whereas MCF7 cells showed increased p70S6K (T421/S424) phosphorylation.

Figure 1. Mesenchymal morphology induction after stimulation with the EMT Inducing Media Supplement. (A – D) Epithelial morphology in the four cell types indicated following culture for 5 days in standard culture media without the EMT Inducing Media Supplement. (E – H) Mesenchymal morphology in the four cell types indicated cultured with the EMT Inducing Media Supplement for 5 days.

Figure 2. Downregulation of epithelial marker expression and upregulation of mesenchymal marker expression in EMT-induced cells. Cells were stained by immunofluorescence to detect the expression of the epithelial marker, E-cadherin (red), and the mesenchymal marker, Fibronectin (green). (A – D) Control cells cultured without the EMT Inducing Media Supplement for 5 days. (E – H) Cells cultured with the EMT Inducing Media Supplement for 5 days. All cells are counterstained with DAPI (blue).
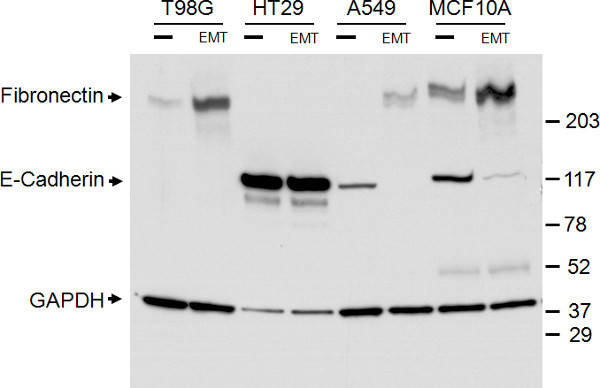
Figure 3. Confirmation of epithelial and mesenchymal marker expression with western blot analysis. Western blot of total cell lysates of the four cell types indicated, treated with (EMT) or without (-) the EMT Inducing Media Supplement for 5 days. Bands for E-cadherin, Fibronectin, and the GAPDH loading control are as indicated on the left. The size marker is as indicated on the right.

Figure 4. Upregulation of the mesenchymal markers, Vimentin and Snail in EMT-induced cells. Cells were stained by multicolor immunofluorescence to detect expression of the epithelial marker, E-cadherin (gray), and the mesenchymal markers, Vimentin (green) and Snail (red). (A – C) Control cells cultured without the EMT Inducing Media supplement for 5 days. (D – F) Cells cultured with the EMT Inducing Media Supplement for 5 days.

Figure 5. Induction of EMT in TGF-β1 non-responsive cells. (A and B) Cells cultured with media alone for 5 days. (C and D) Cells cultured in media containing 10 ng/ml of TGF-β1 for 5 days. (E and F) Cells cultured in EMT Inducing Media Supplement for 5 days. All cells are stained for E-cadherin expression (red) and counterstained with DAPI (blue).

Figure 6. EMT-induced cells have increased migration and invasion capacities. A. Average number of cells that migrated through an 8 μm pore filter after incubation for 48 hr. B. Average number of cells that were able to invade through an 8 μm pore filter coated with basement membrane extract after incubation for 48 hr. Error bars indicate the standard deviation over 3 wells.

Figure 7. Antibody-based array expression analysis of EMT induction. Lysates from A549 (A and C) and MCF7 (B and D) cells cultured with or without EMT Inducing Media Supplement were used for array analysis using the Proteome Profiler Human Phospho-MAPK Array Kit. Spots highlighted in A and B were then quantitated using densitometry. Comparisons of un-induced to EMT-induced lysates are shown in the corresponding bar graphs below (C and D). Click here to view larger figure.
